Secreted enzymatic activities of ectomycorrhizal fungi as a case study of functional diversity and functional redundancy
12
0
0
Texte intégral
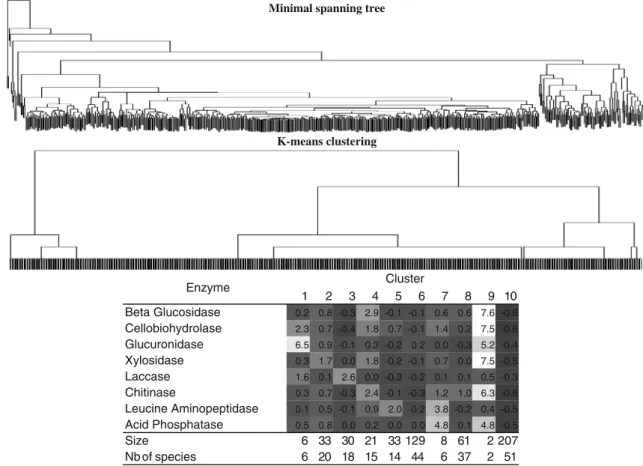
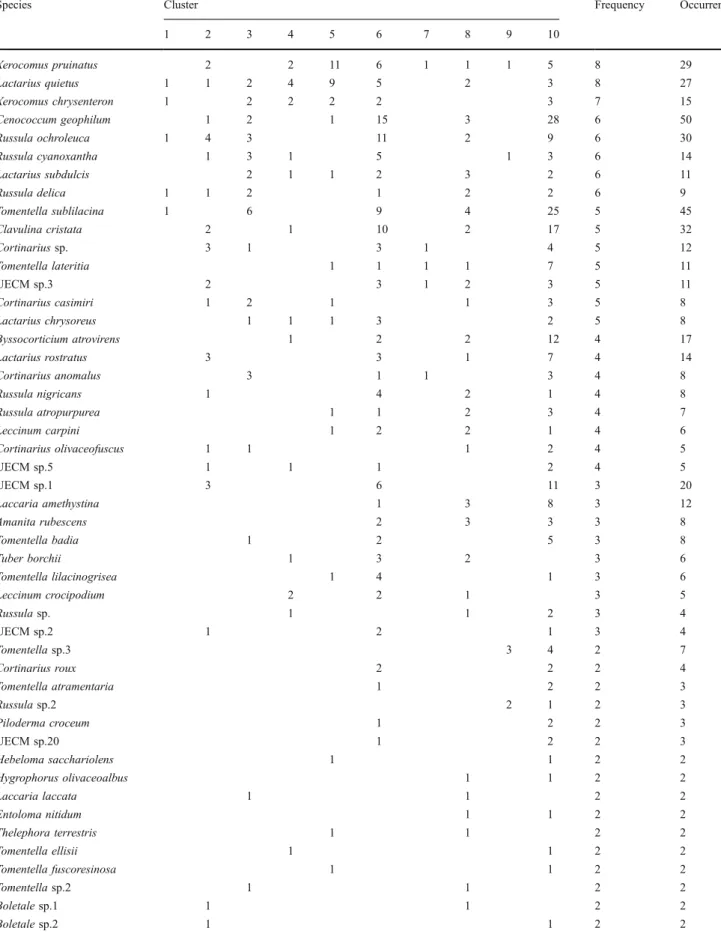
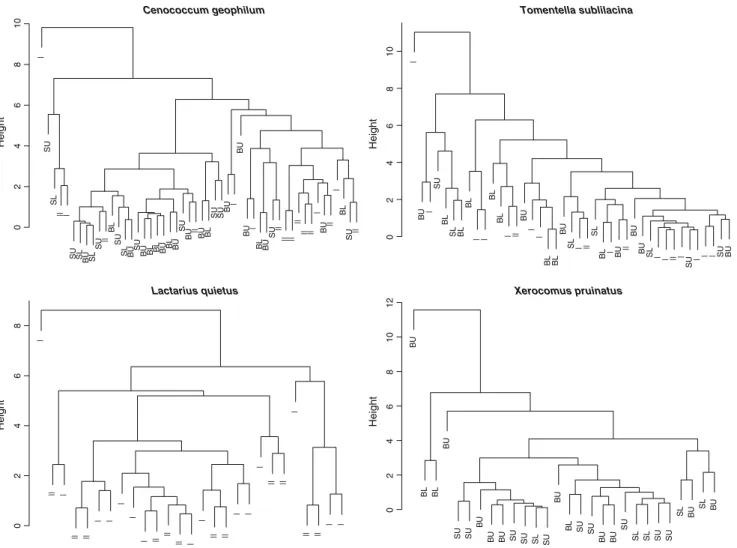
Figure
Documents relatifs
