Asymmetries of the visual system and their influence on visual performance and oculomotor dynamics
45
0
0
Texte intégral
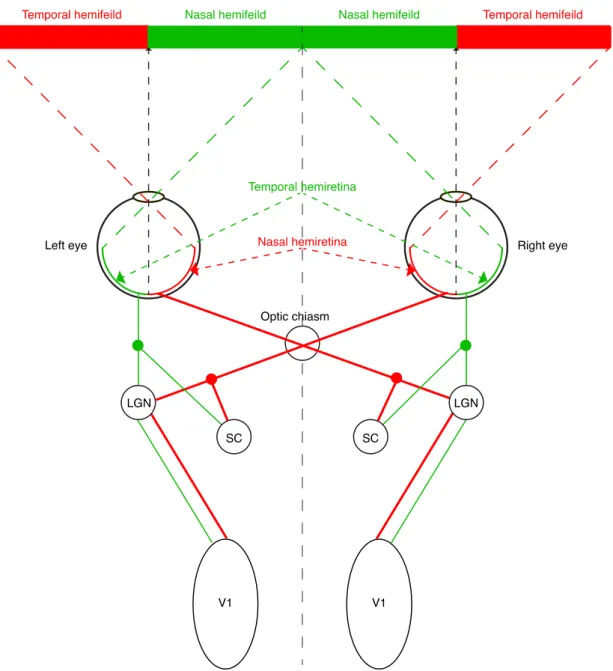
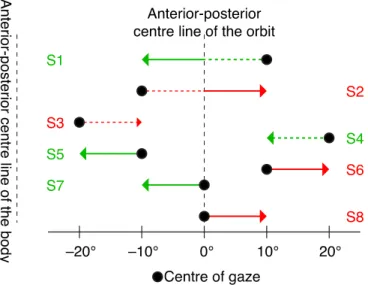
Figure
Documents relatifs