Characterization of N-glycans from mouse brain neural cell adhesion molecule
12
0
0
Texte intégral
Figure
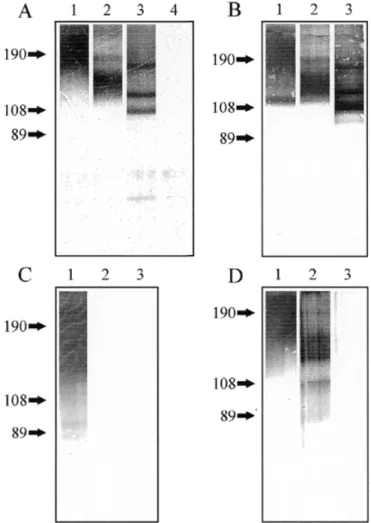
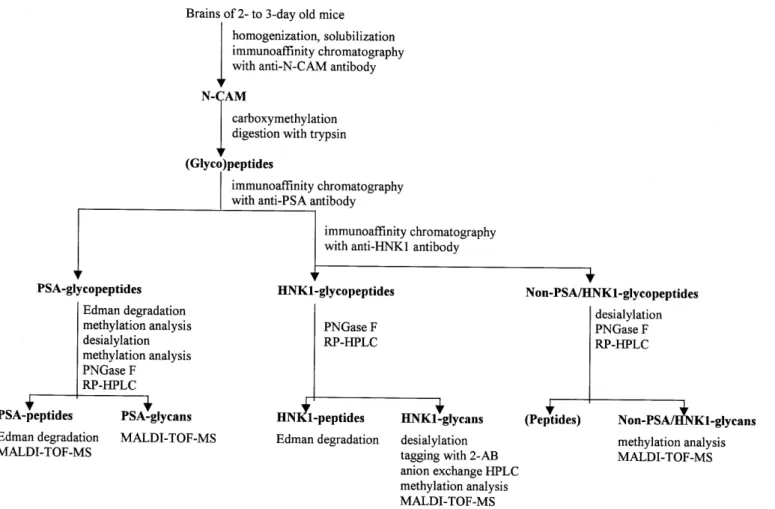
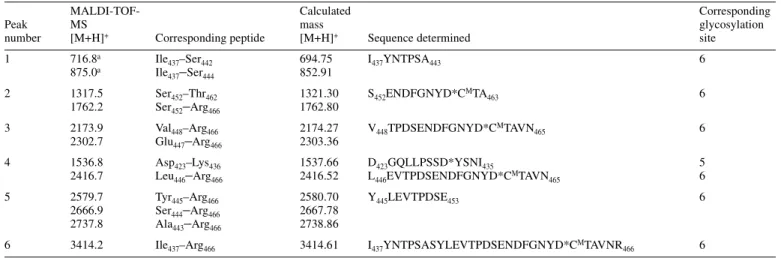
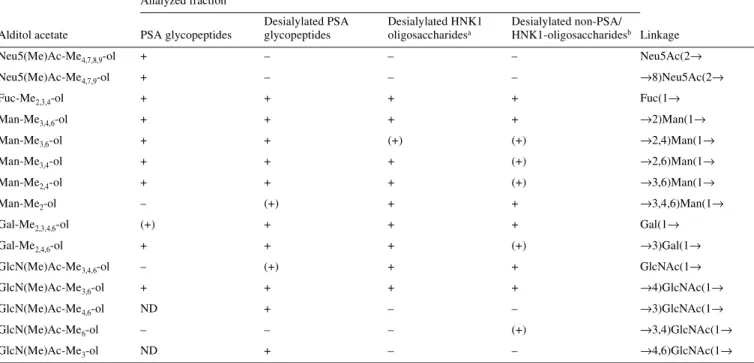
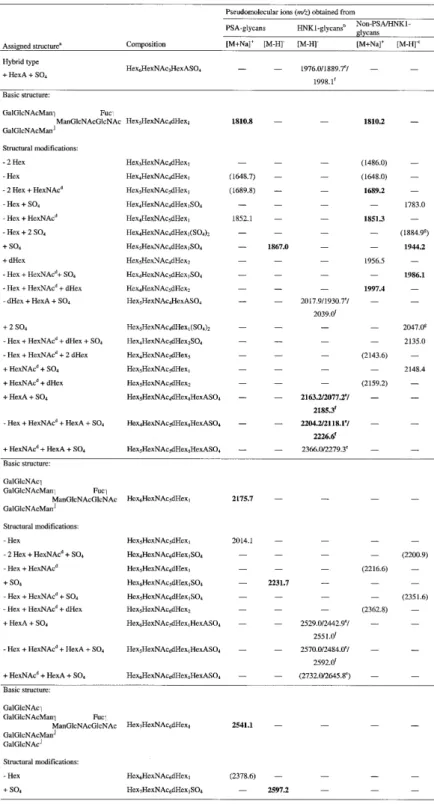
![Fig. 5. MALDI-TOF mass spectra of native desialylated PSA glycans. Glycans were analysed in the positive-ion (A) or negative-ion (B) reflectron mode and were recorded as [M+Na] + or [M-H] – pseudomolecular ions, respectively.](https://thumb-eu.123doks.com/thumbv2/123doknet/14920992.662932/5.892.67.424.143.582/desialylated-glycans-analysed-positive-negative-reflectron-pseudomolecular-respectively.webp)
+4
Documents relatifs