AGE DETERMINATION OF ARIANTA ARBUSTORUM (L.) (PULMONATA) BASED ON GROWTH BREAKS AND INNER LAYERS
5
0
0
Texte intégral
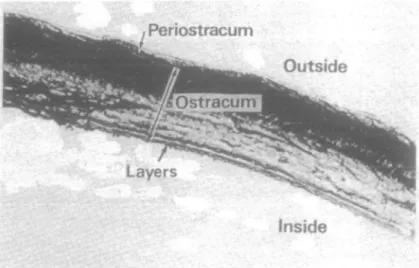
Figure


Documents relatifs