Can soil testate amoebae be used for estimating the time since death? : A field experiment in a deciduous forest
Texte intégral
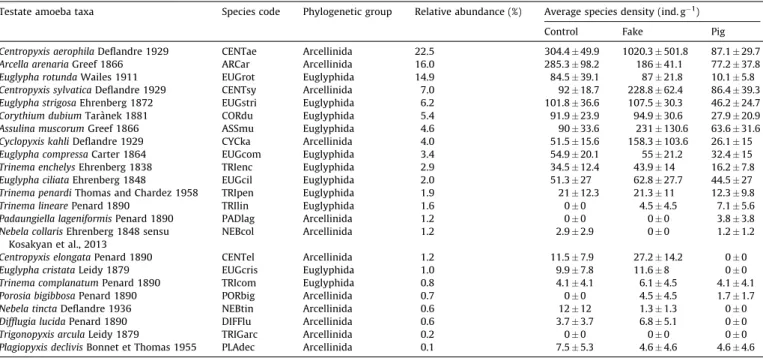
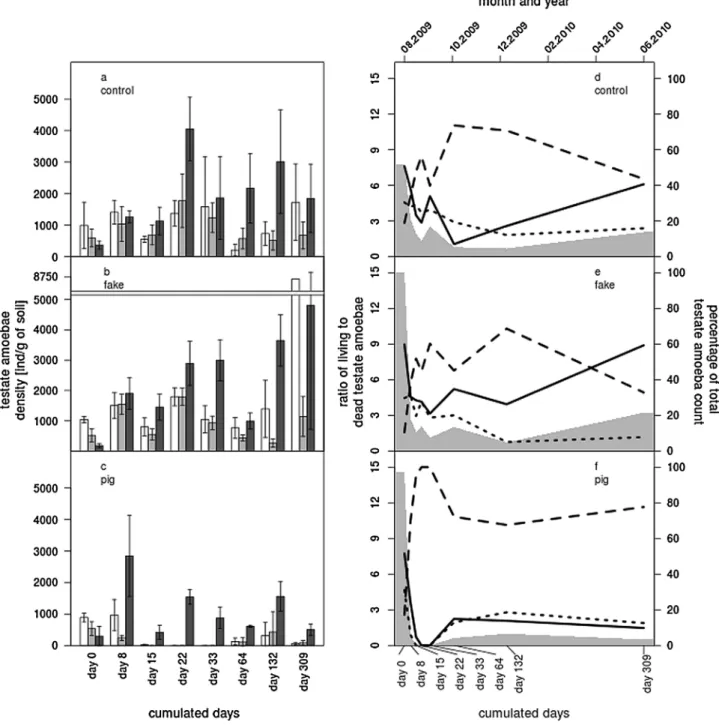
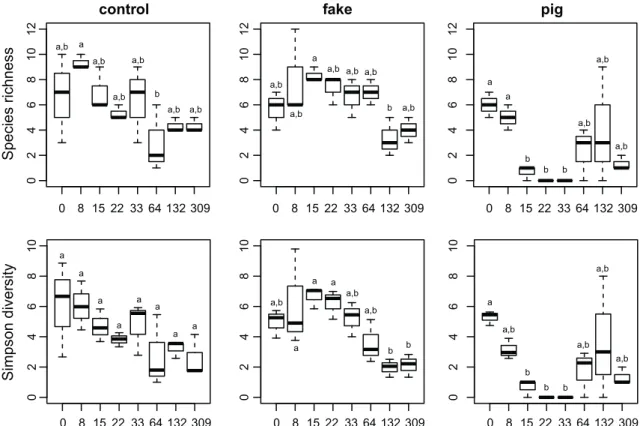
Figure
Documents relatifs
Using this partition of the subsystems and particularly exploiting the time margins corresponding to idle times of machines, the approach allows an optimization of the non-critical
“the distance between the robot and the human operator is always greater than or equal
This mountainous region is drained by several coastal rivers to the Pacific Ocean, and original sediment fingerprinting methods had to be developed to track the dispersion
Whereas all these measures were, in the US view, subsidies pursuant to Articles 1 and 2 of the WTO Agreement on Subsidies and Countervailing Measures ( ‘SCM Agreement ’ or
See Matthias Lehmann, "Volcker Rule, Ring-Fencing or Separation of Bank Activities – Comparison of Structural Reform Acts around the World," 10-12.;
A third model would involve consulting the relatives of the deceased (unless the deceased had made specific provision for this situation whilst still alive)
The Eportfolio: How can it be used in French as a second language teaching and learning.. Practical paper presenting
Furthermore, as shown by Exam- ple 1, in order to determine this differential Hilbert function it may be necessary to compute derivatives of the original equations up to order ν,
