Reaching with the sixth sense: Vestibular contributions to voluntary motor control in the human right parietal cortex
7
0
0
Texte intégral
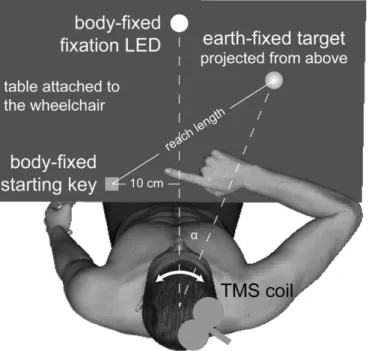
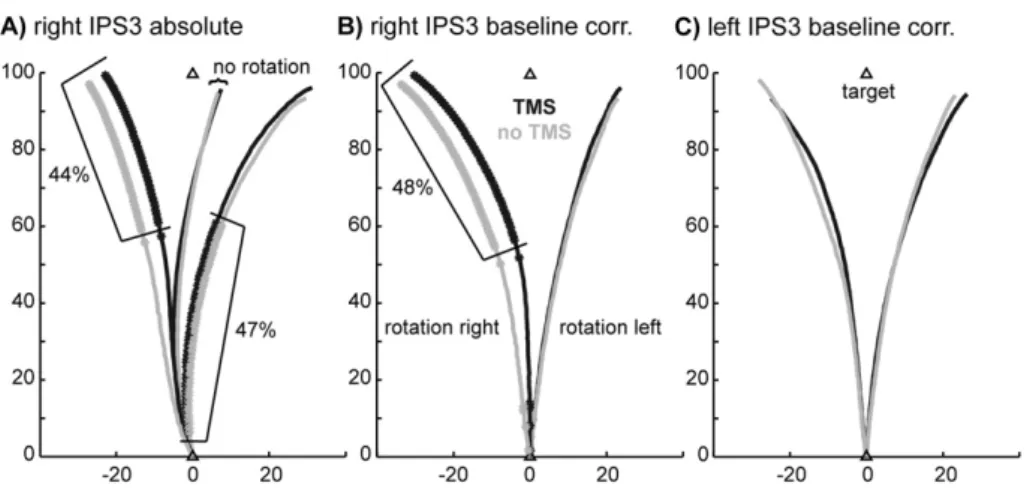
Figure
Documents relatifs