HAL Id: hal-02353351
https://hal.umontpellier.fr/hal-02353351
Submitted on 7 Nov 2019
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of
sci-entific research documents, whether they are
pub-lished or not. The documents may come from
teaching and research institutions in France or
abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est
destinée au dépôt et à la diffusion de documents
scientifiques de niveau recherche, publiés ou non,
émanant des établissements d’enseignement et de
recherche français ou étrangers, des laboratoires
publics ou privés.
Adaptation of Paramecium caudatum to variable
conditions of temperature stress
Alison Duncan, Simon Fellous, Elsa Quillery, Oliver Kaltz
To cite this version:
Alison Duncan, Simon Fellous, Elsa Quillery, Oliver Kaltz. Adaptation of Paramecium caudatum to
variable conditions of temperature stress. Research in Microbiology, Elsevier, 2011, 162 (9),
pp.939-944. �10.1016/j.resmic.2011.04.012�. �hal-02353351�
Adaptation of Paramecium caudatum to variable conditions of temperature
stress
Alison B. Duncan
*
, Simon Fellous, Elsa Quillery, Oliver Kaltz
Institut des Sciences de l’Evolution (ISEM), UMR 5554, Universite´ Montpellier 2, Place Eugene Bataillon, 34095 Montpellier cedex 05, France Received 19 October 2010; accepted 4 April 2011
Abstract
The environment is rarely constant and organisms are exposed to spatial and temporal variation that will impact life-histories. It is important to understand how such variation affects the adaptation of organisms to their local environment. We compare the adaptation of populations of the ciliate Paramecium caudatum exposed to constant (23!C or 35!C) and temporally variable temperature environments (random daily
fluctu-ations between 23!C or 35!C). Consistent with theory, our experiment shows the evolution of specialists when evolution proceeds in constant
environments and generalists when the environment is temporally variable. In addition, we demonstrate costs for specialists of being locally adapted through reduced fitness in novel environments. Conversely, we do not find any costs for generalists, as all populations from variable environments had equal or superior performance to specialists in their own environment. The lack of a cost for generalists is emphasised by the presence of a super generalist that has the highest performance at both assay temperatures.
Keywords: Local adaptation; Specialists; Generalists; Temperature variation; Thermal stress
1. Introduction
Environmental heterogeneity is considered a universal driver of evolutionary change and adaptation (Levins, 1968; Bell, 1997, 2010). Indeed, in nature, organisms are expected to be adapted to the environment, or environments, they experience most commonly. This manifests itself in the evolution of specialists when organisms are exposed to a mostly constant environment, and generalists when changes in environmental state are frequent (Levins, 1968; Kassen, 2002). Accordingly, evolution in spatially structured, constant environments should generate specialists that maxi-mise fitness in their own environment. However, adaptation in one environment is predicted to arise at a cost expressed as low performance in other environments. This could happen due to antagonistic pleiotropy where genes beneficial in one
environment are costly in another. Consequently, evolution of a specialist should result in patterns of local adaptation whereby locally selected residents have a higher fitness than foreign, unselected non-residents (Kawecki and Ebert, 2004). In contrast, temporally variable environments should select for generalist strategies that enable persistence in all envi-ronments encountered. Thus, rather than maximising fitness within environments, selection for generalists should maxi-mise average fitness over all environments. As a result, the fitness of generalists in each single environment may be less than that of respective specialists. This is comparable to a low-risk “bet-hedging” strategy, minimising the variance in performance between environments and allowing at least some average performance in all environments encountered (Stearns, 2000; Kassen, 2002).
The evolution of generalists depends on various genetic and ecological factors, such as the amount of standing genetic variation in populations and environmental structure (Kassen, 2002). An important aspect of the temporal environment is the frequency of exposure to different environments (Levins, * Corresponding author.
E-mail address:ali.b.duncan@gmail.com(A.B. Duncan).
Please cite this article in press as: Duncan, A.B., et al., Adaptation of Paramecium caudatum to variable conditions of temperature stress, Research in doi:10.1016/j.resmic.2011.04.012
1968; Bell, 2010). If frequency of exposure to one environ-ment is common and the other rare, local adaptation to the rare environment may not occur. However, if the rare environment is particularly harsh, adaptation to it may be required to guarantee survival following its occurrence.
Microbial organisms are ideal to study adaptation to a variable environment due to their short generation time, large population sizes and the ability to manipulate their environ-ment in a controlled fashion (Jessup et al., 2004). In this study, we use experimental microcosms to investigate the evolution of specialists and generalists in spatially and temporally variable environments. This experiment is the first to investi-gate how different frequencies of exposure to different envi-ronments influences the evolution of generalists. We cultured experimental populations of the ciliate Paramecium caudatum in two constant temperature environments (23!C and 35!C)
and in 4 temporally variable temperature environments, with random daily fluctuations between 23 !C and 35 !C. These
variable environments differed in the frequency of occurrence of temperature stress (35!C), ranging from rare (w25% of the
time) to frequent (w75%). After 4 months (30e40 genera-tions), survival and replication at 23 and 35!C were compared
among populations from the different treatments. The constant environments should select for specialists. Therefore, we predicted performance of populations tested in their own environment to be superior to performance in foreign constant temperature environments (i.e., local adaptation). The variable environments should select for generalists. Thus, we expected the performance of Paramecium from variable treatments to have intermediate fitness to those evolved in constant envi-ronments. Further, we expected performance of Paramecium evolved in the different variable environments to differ in each of the constant assay environments according to the amount of time previously spent at each temperature.
2. Materials and methods 2.1. Study organisms
P. caudatum is a freshwater ciliate found in still water bodies in the northern hemisphere that feeds on bacteria and detritus within the water column (Wichterman, 1986). Reproduction is predominantly asexual through mitotic divi-sion. Under exponential growth conditions, P. caudatum divides 1e3 times every 24 h and optimal growth temperatures generally range between 24 and 28 !C (Wichterman, 1986).
There is also evidence of within-species genetic variation in growth and survival at different temperatures (Fels and Kaltz, 2006). In our laboratory, Paramecium are maintained at 23!C,
in a culture medium of dried organic lettuce supplemented with the bacterium Serratia marcescens as food (Nidelet,
2007). Two Paramecium clones were used in this experi-ment, clone K8 and clone VEN (see (Duncan et al., 2010) for details). Each population in the selection experiment comprised 20 ml from mass cultures of each clone in a 50 ml Falcon tube.
2.2. Selection experiment
In a 120-day selection experiment, populations of P. caudatum were randomly assigned to one of 6 selection treatments, two constant temperature treatments (23 !C, and
35!C " 0.5!C) and four variable temperature treatments (with
mean temperatures of 26.2!C, 27.8!C, 30.2!C or 32.8!C).
These variable temperature environments corresponded to 27%, 40%, 60% and 73% of time during the experiment spent at 35!C,
respectively. Variable mean temperatures were achieved by randomly changing tubes daily between 23!C and 35!C. Each
tube was randomly allocated an individual sequence corre-sponding to its mean temperature. At weekly intervals, 3 ml of each population was removed and replaced with fresh culture medium containing 50 P. caudatum from source populations of the same clone kept at 23!C.Table 1shows mean population
sizes prior to the onset of the experiment. All populations were larger than the 50 individuals added each week, indicating self-sustaining populations even at higher temperatures. The experiment contained a total of 48 replicate populations, with 4 replicates per genotype and treatment (2 genotypes # 6 selection treatments # 4 replicate populations). On day 120 of the selection experiment, two 1-ml samples were removed from each population and each transferred to a 1.5 ml Eppendorf tube with 500 ml of medium. One sample was placed at 23!C and the
other at 35!C for a 48 h acclimation period.
2.3. Adaptation assay
We phenotyped the division of individual Paramecium, from each replicated population at 23 !C and 35 !C. Four
Paramecium cells were individually isolated from each 1 ml sample that had experienced a 48 h acclimation period prior to the onset of the experiment. Each individual cell was placed in a 60 ml drop of culture medium arranged inside the lids of 24-microwell plates (Nunc", Fisher Scientific, France). The Paramecium were checked for survival and division 24, 30, 36 and 48 h after start of the experiment.
2.4. Statistical analysis
We used repeated-measures ANOVA to analyse the performance of individual Paramecium at 23!C and 35!C for
48 h after onset of the experiment. Performance was measured as the log2(number of cells þ 1) in each 60 ml drop through Table 1
Mean population sizes for Paramecium populations in each of the selection environments prior to the onset of the experiment ("standard error).
23!C Constant 26!C Variable 28!C Variable 30!C Variable 32!C Variable 35!C Constant
812 ("118) 1188 ("412) 867 ("458) 565 ("221) 281 ("148) 372 ("284)
2 A.B. Duncan et al. / Research in Microbiology xx (2011) 1e6
time. Selection treatment, assay temperature and time were included in models as fixed factors and host clone, replicate population and assay microwell plate as random factors. Non-significant terms were removed from models in a stepwise fashion ( p > 0.10).
Unlike specialists, generalists are predicted to minimise differences in fitness between environments. To test this hypothesis, we averaged log2(number of cells þ 1) after 48 h
for each replicate population at 23 (x23!C) and 35!C (x35!C).
FollowingBell (2010), we calculated the environmental vari-ance as Var(E) ¼ 1/2 (x23!C & x35!C)2 for each replicate
population. In an ANOVA, we tested for difference in Var(E) among selection treatments. All analysis was done using JMP 8 (SAS, 2008).
3. Results
There was a steady increase in density during the experi-ment for all populations at 23 !C. In contrast, populations at
35!C increased only marginally during the first 24 h of the
experiment before stopping division or death. Time series analysis revealed a significant 3-way interaction between time, assay temperature and selection temperature (F5, 1190¼ 2.73,
p ¼ 0.0183). This means that differences in relative perfor-mance of populations from the different constant and variable selection treatments, at the two assay temperatures, became apparent over the course of the assay. Paramecium from the 23!C constant treatment tended to grow better at 23!C than
those from the 35!C constant treatment. The reverse was true
for Paramecium from the 35 !C treatment, that had greater
survival at 35 !C than Paramecium from the 23 !C constant
treatment. Paramecium from the variable treatments varied in their responses, with their relative ranks depending on the
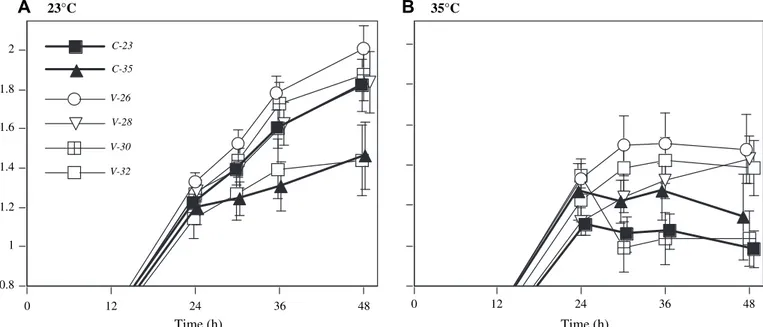
assay temperature and identity of the variable treatment (Fig. 1). The host clone and its interactions were not signifi-cant and never explained more than 3.5% of the variance in the model.
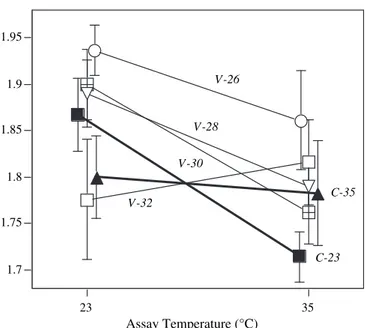
Fig. 2illustrates general patterns observed during the 48 h period for Paramecium from the different selection treatments. Crossing of reaction norms for growth of Paramecium from the two constant environments at the two assay temperatures is clearly shown. Performances of Paramecium from populations with variable selection treatments were generally equal to or greater than resident lines from the respective constant treat-ment. One variable treatment (populations with a mean of 26 !C) had the highest performance at both assay
tempera-tures, indicating the presence of a super generalist.
The environmental variance in performance Var(E) did not significantly differ among selection treatments (F5, 28¼ 0.85,
p ¼ 0.5264). Thus, there was no evidence that populations from variable (generalist) treatments had reduced variation in performance at 23 and 35!C compared to those from constant
(specialist) treatments. Fig. 2 illustrates this comparable variation in performance between populations from constant and variable treatments at 23 !C and 35!C.
We projected the fitness of Paramecium at variable mean temperatures by combining fitness measured at 23 !C and at
35 !C. We calculated mean (geometric) fitness of variable
generalists and constant specialists at different variable treat-ment mean temperatures. The geometric mean is a more appropriate measure of fitness across temporally variable environments (Bell, 2010). Fig. 3 shows better fitness of generalists in all variable environments, in particular for increasing mean temperatures, i.e., in environments with more frequent exposure to 35 !C. Some generalists even appear
capable of invading constant specialist populations, in
0 12 24 36 48 Time (h) 35°C 0.8 1 1.2 1.4 1.6 1.8 2 0 12 24 36 48 Time (h) 23°C V-32 V-30 V-28 V-26 C-35 C-23
A
B
Fig. 1. Changes in mean number of paramecia through time (24, 30, 36, 48 h), measured at 23!C (A) and 35!C (B), starting from a single individual. Means shown
for 2 constant temperature treatment origins (23!C ¼ C-23; 35!C ¼ C-35) and for 4 variable treatment origins (fluctuating between 23 and 35!C, with means
between 26 and 32!C: V-26, V-28, V-30, V-32). Numbers are log
2-transformed; thus values >1 indicate an increase in number. Error bars denote standard errors. Please cite this article in press as: Duncan, A.B., et al., Adaptation of Paramecium caudatum to variable conditions of temperature stress, Research in
particular, the stressful constant 35 !C environment. One
generalist (V26, from a variable treatment with a mean of 26!C)
may even be universally best fit across all environments.
4. Discussion
We observed a crossing of reaction norms for Paramecium from the different constant selection treatments when tested at 23!C and 35!C. We will first discuss the implications of these
results in terms of evolution of specialists in constant
environments and then the evolution of generalists in variable environments.
4.1. Specialisation in constant environments
Populations from the two constant temperature environ-ments (23!C and 35!C) were observed to have higher levels
of division when assayed at their own selection temperature. This finding is consistent with the idea that spatially hetero-geneous environments favour the evolution of specialists, each adapted to their own environment (Kawecki and Ebert, 2004). Experimental evolution of specialisation to different (constant) temperature regimes has also been shown, e.g., for the bacterium Escherichia coli and the green alga Chlamydomo-nas rheinhardtii reviewed in (Kassen, 2002). Although our results are coherent with reciprocal selection in each constant environment, this may not actually be the case. All populations had been cultured at 23!C for a number of years prior to the
onset of the experiment, such that 4 months of additional selection may not improve fitness at this temperature (see also (Bennett et al., 1992)). Accordingly, the observed cross-reaction norms may simply reflect a change in Paramecium cultured under temperature stress (35!C). Thus, adaptation to
35!C apparently comes at a cost whereby populations have
reduced performance at 23!C.
The mechanisms responsible for the observed patterns are not clear. The observed results are coherent with genetic change and evolution occurring in the different populations. Indeed, previous work has indicated a genetic basis of temperature-dependent variation in growth and survival in P. caudatum (Fels and Kaltz, 2006; Duncan et al., 2010). Thus, given the duration of our long-term experiment (4 months), and the population sizes at 35!C (500e1000 individuals), this
is a plausible explanation. However, it is also possible that these results are attributable to acclimation of the Paramecium to the different temperature environments. It is clear, however, from our results, that the 48 h conditioning period prior to the onset of the experiment did not efface local adaptation in our experiment. Previous work has shown that patterns of accli-mation for Paramecium tetraurelia to stressful temperatures (35!C as in our experiment) are lost within 24 h (Hennessey and Nelson, 1979). This result suggests that 48 h is sufficient to eliminate acclimation and that our results are consistent with evolution. However, another study found that patterns of acclimation for P. caudatum between more permissive envi-ronments (10 !C and 25 !C) remained for up to a week
(Tsukuda and Takeuchi, 1984). Molecular analysis affirming genetic change in the experiment is required to distinguish between these hypotheses. In addition, true understanding of the mechanisms behind these patterns will require more detailed experiments regarding the time required for accli-mation and its subsequent duration.
The responses of Paramecium from the different pop-ulations may hint at the underlying physiological mechanisms. At 35 !C, the resident population selected at 35 !C has
a division advantage only during the first 24 h before division declines at the same rate as Paramecium from 23!C. This is 1.7 1.75 1.8 1.85 1.9 1.95 23 35 Assay Temperature (°C) C-23 C-35 V-26 V-28 V-30 V-32
Fig. 2. Mean cumulative paramecium growth over 48 h at two assay temperatures (23 and 35!C), as measured by the area under the curve AUC,
based on log2-transformed (number of paramecia þ1). Means shown for 2 constant temperature treatment origins (C-23, C-35) and for 4 variable treat-ment origins (V-26, V-28, V-30, V-32). Error bars denote standard errors.
Fig. 3. Mean fitness of constant (C-23, C-35) and variable (V-26, V-28, V-30, V-32) treatment origins estimated for each of the 4 variable treatment envi-ronment temperature means. Fitness was estimated from the mean log2 -transformed number of individuals at 23 or 35!C after 48 h. For constant
environments, fitness is the arithmetic mean at 23 or 35!C, respectively. For
variable environments, geometric means for performance at 23 and 35!C were
calculated and weighted for the relative frequency of time spent at the two temperatures.
4 A.B. Duncan et al. / Research in Microbiology xx (2011) 1e6
consistent with a constitutive heat stress protection mecha-nism, such as altered membrane structure (Sasaki et al., 2006). In contrast, at 23 !C, the resident advantage is cumulative,
increasing with time, whereas growth from populations orig-inating from 35!C increases at first before levelling off. An
absence of sustained growth for Paramecium from 35!C may
be attributable to the cost of the heat-stress protection mech-anism when resources (food, oxygen) become limiting in the 60 ml drop. Although not expected to serve a constitutive function for heat tolerance, fitness costs of HSP over-expression have been demonstrated in various organisms (Ketola et al., 2004; Sorensen et al., 2003).
4.2. Evolution of generalists in variable environments Our results clearly showed that Paramecium from variable selection environments grow reasonably well at both 23 !C
and 35 !C, consistent with the evolution of a generalist
strategy. However, patterns of growth for populations from the different variable treatments were not the same across pop-ulations, nor was the performance of these populations inter-mediate between those of the constant-temperature specialists, as would be expected for typical generalists e.g., (Legros and Koella, 2010). Indeed, environmental variance in growth between the two assay temperatures for populations from variable environments was comparable to that observed for populations from constant environments. This result is inconsistent with the evolution of a bet-hedging strategy pre-dicted for generalists, but consistent with other empirical observations (Leroi et al., 1994; Reboud and Bell, 1997; Kassen and Bell, 1998; Barrett et al., 2005). It seems that generalist evolution here does not select for intermediate fitness across environments encountered, but instead permits optimal adaptation to different components of the environment presented (here: 23!C and 35!C).
Further, one novel aspect of our experiment was the use of different types of variable environments, where we manipu-lated the time that populations spent at 23 !C and 35 !C.
We did not observe that evolution in the performance of Paramecium at the different variable treatments varied in the constant assay environments according to the amount of time they had spent in each. The different relative frequencies of exposure to 35!C or 23 !C had no obvious influence on the
order of performance at either assay temperature, nor on the order of the average performance over both temperatures. Combined, these results emphasise an absence of trade-off ensuing different amounts of time spent at each temperature, and no cost for being a generalist in this experiment.
Instead, our results indicate higher fitness of (some) gener-alists in all variable environments, in particular at increasing mean temperatures, with more frequent exposure to 35!C. Thus,
most variable treatments appear to be better adapted to variable environments than constant treatments (Fig. 3). Moreover, some of the variable treatments produced populations that may even be capable of invading the specialist populations at constant temperatures, especially at 35 !C. In fact, one variable
envi-ronment (V-26) seems to have generated supergeneralists with
over all superior growth at both assay temperatures, and highest projected fitness across all variable mean temperatures (Fig. 3). The absence of a cost for our generalist populations may be attributable to the evolutionary consequences of adapting in a variable environment (Lenormand et al., 2009). Fluctuations in selective pressure can induce “Evolutionary revolutions” sensu
Lenormand et al. (2009). That is, when ancestral populations are fixed to a local fitness peak in an adaptive landscape (in our case populations initially adapted to 23C), stochasticity in selection can allow populations to reach a peak higher in the fitness landscape. Here, transient exposure to 35 !C could help
Paramecium find a strategy that was initially worse at 23!C but
eventually leads to higher fitness at both temperatures. This phenomenon can be compared to "roundabout selection" as exemplified byMacLean et al. (2002).
The physiological mechanism for heat tolerance may not be the same for constant and variable temperature treatments. For example, heat shock proteins are associated with a response that is induced relatively quickly following expo-sure to heat stress (Feder and Hofmann, 1999) and may not last. Therefore, in temporally variable environments, increased heat stress tolerance may be mediated by selection on the HSP machinery (Ketola et al., 2004). In contrast, constitutive HSP overexpression may be less rewarding under sustained exposure to heat stress, because of the costs asso-ciated with it (Sorensen et al., 2003; Ketola et al., 2004). Thus long-term constant exposure to high temperature may even lead to lower HSP expression levels (Cavicchi et al., 1995; Bettencourt et al., 1999). Heat tolerance in our constant 35 !C treatment may therefore have another mechanistic
basis, such as the altered membrane structures mentioned above (Sasaki et al., 2006; Toyoda et al., 2009). The costs associated with each mechanism may be very different, which may explain the different reaction norms for populations from constant and variable environments observed in this experi-ment. Alternatively, the costs we observe for specialists in foreign environments may arise following mutation accumu-lation of genes/alleles that are only deleterious in foreign environments (Kassen, 2002). Accumulation of such genes will not occur in variable environments because they will be consistently selected against.
4.3. Conclusions
This experiment demonstrates that specialists appear when selection occurs in constant environments, and costs associ-ated with specialisation are consistent with theory. In contrast, evolution under variable conditions produces generalists with often superior fitness at both assay temperatures. Although we cannot be sure, we postulate that there may be different mechanisms mediating heat tolerance for generalists and specialists in our experiment. Constitutive and induced quan-tification of heat shock protein expression in Paramecium would confirm this hypothesis. Aside from the mechanism, we cannot distinguish whether the phenotypic differences between populations are genetic, or due to trans-generational acclimation. Confirmation of this hypothesis would require
molecular analysis demonstrating genetic change in pop-ulations. Regardless of the mechanism, we demonstrate that patterns of local adaptation unravel differently in constant and variable environments.
Acknowledgements
We would like to thank two anonymous reviewers for helpful comments on the manuscript. This work was supported by grants from the Universite´ Montpellier 2 (A. B. D.), the Agence National de la Recherche (“ANR-09-BLAN-0099”, A. B. D., S. F. and O. K.) and the Centre National de Recherche Scientifique (programme “INSU-EC2CO-Microbien” N!125
to O.K.). This is ISEM contribution 2011e029.
References
Barrett, R.D., MacLean, R.C., Bell, G., 2005. Experimental evolution of pseudomonas fluorescens in simple and complex environments. Am. Nat. 166, 470e480.
Bell, G., 1997. Selection: The Mechanism of Evolution. Chapman & Hall, Florence, KY.
Bell, G., 2010. Fluctuating selection: the perpetual renewal of adaptation in variable environments. Phil. Trans. R. Soc. B-Biol. Sci. 365, 87e97. Bennett, A.F., Lenski, R.E., Mittler, J.E., 1992. Evolutionary adaptation to
temperature. I. Fitness responses of Escherichia coli to changes in its thermal environment. Evolution 46, 16e30.
Bettencourt, B.R., Feder, M.E., Cavicchi, S., 1999. Experimental evolution of hsp70 expression and thermotolerance in drosophila melanogaster. Evolution 53, 484e492.
Cavicchi, S., Guerra, D., Latorre, V., Huey, R.B., 1995. Chromosomal analysis of heat-shock tolerance in drosophila-melanogaster evolving at different temperatures in the laboratory. Evolution 49, 676e684.
Duncan, A.B., Fellous, S., Accot, R., Alart, M., Chantung Sobandi, A., Cosiaux, A., Kaltz, O., 2010. Parasite-mediated osmotic stress protection of paramecium caudatum infected by holospora undulata is host genotype specific. FEMS Microbiol. Ecol. 74, 353e360.
Feder, M.E., Hofmann, G.E., 1999. Heat-shock proteins, molecular chaper-ones, and the stress response: evolutionary and ecological physiology. Ann. Rev. Physiol. 61, 243e282.
Fels, D., Kaltz, O., 2006. Temperature-dependent transmission and latency of holospora undulata, a micronucleus-specific parasite of the ciliate para-mecium caudatum. Proc. R. Soc. B-Biol. Sci. 273, 1031e1038.
Hennessey, T., Nelson, D.L., 1979. Thermosensory behavior in paramecium-tetraurelia e quantitative assay and some factors that influence thermal avoidance. J. Gen. Microbiol. 112, 337e347.
Jessup, C.M., Kassen, R., Forde, S.E., Kerr, B., Buckling, A., Rainey., P.B., Bohannan, B.J., 2004. Big questions, small worlds: microbial model systems in ecology. Trends Ecol. Evol. 19, 189e197.
Kassen, R., 2002. The experimental evolution of specialists, generalists, and the maintenance of diversity. J. Evol. Biol. 15, 173e190.
Kassen, R., Bell, G., 1998. Experimental evolution in chlamydomonas. Iv. Selection in environments that vary through time at different scales. Heredity 80, 732e741.
Kawecki, T.J., Ebert, D., 2004. Conceptual issues in local adaptation. Ecol. Lett. 7, 1225e1241.
Ketola, T., Laakso, J., Kaitala, V., Airaksinen, S., 2004. Evolution of hsp90 expression in tetrahymena thermophila (protozoa, ciliata) populations exposed to thermally variable environments. Evolution 58, 741e748. Legros, M., Koella, J.C., 2010. Experimental evolution of specialization by
a microsporidian parasite. BMC Evol. Biol. 10, 159.
Lenormand, T., Roze, D., Rousset, F., 2009. Stochasticity in evolution. Trends Ecol. Evol. 24, 157e165.
Leroi, A.M., Bennett, A.F., Lenski, R.E., 1994. Temperature acclimation and competitive fitness: an experimental test of the beneficial acclimation assumption. Proc. Nat. Acad. Sci. U.S.A 91, 1917e1921.
Levins, R., 1968. Evolution in Changing Environments. Princton University Press, NJ.
MacLean, R.C., Bell, G., 2002. Experimental adaptive radiation in Pseudo-monas. Am. Nat. 160, 569e581.
Nidelet, T., 2007. L’effet de la structuration spatiale et de l’he´te´roge´ne´ite´ environnementale sur les interactions hoˆte-parasite: Une approche d’e´volution expe´rimentale et d’e´pide´miologie. UPMC, Univ Paris. Reboud, X., Bell, G., 1997. Experimental evolution in chlamydomonas. Iii.
Evolution of specialist and generalist types in environments that vary in space and time. Heredity 78, 507e514.
SAS, 2008. Jmp Statistics and Graphics Guide (Version 8.0.1.). SAS Institute, Cary, N.C.
Sasaki, T., Konoha, Y., Toyoda, T., Yasaka, Y., Przybos, E., Nakaoka, Y., 2006. Correlation between thermotolerance and membrane properties in para-mecium aurelia. J. Exp. Biol. 209, 3580e3586.
Sorensen, J.G., Kristensen, T.N., Loeschcke, V., 2003. The evolutionary and ecological role of heat shock proteins. Ecol. Lett. 6, 1025e1037. Stearns, S.C., 2000. Daniel bernoulli (1738): evolution and economics under
risk. J. Biosci. 25, 221e228.
Toyoda, T., Hiramatsu, Y., Sasaki, T., Nakaoka, Y., 2009. Thermo-sensitive response based on the membrane fluidity adaptation in paramecium mul-timicronucleatum. J. Exp. Biol. 212, 2767e2772.
Tsukuda, H., Takeuchi, Y., 1984. Heat-resistance and contractile vacuolar activity of paramecium-caudatum acclimated to different temperatures. Comp. Biochem. Physiol. A-Physiol. 77, 641e645.
Wichterman, R., 1986. The Biology of Paramecium. Plenum Press, New York.
6 A.B. Duncan et al. / Research in Microbiology xx (2011) 1e6