Targeted disruption of the MHC class II Aa gene in C57BL/6 mice
8
0
0
Texte intégral
Figure
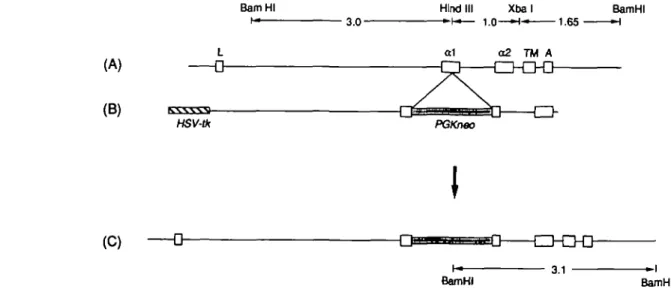

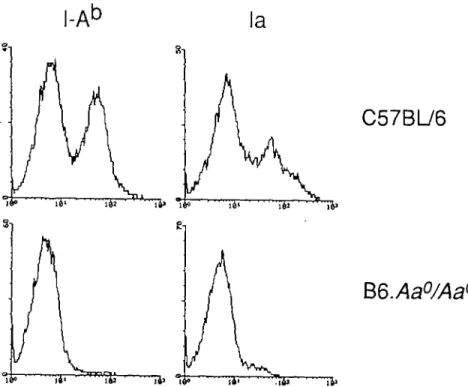
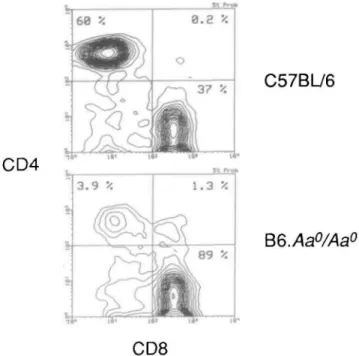
+2
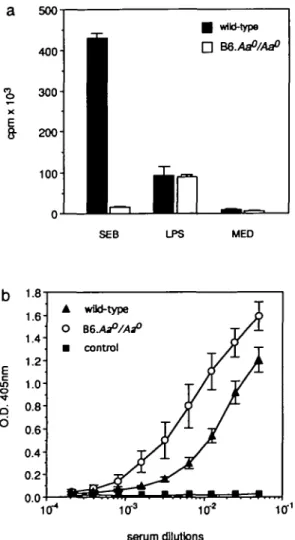
Documents relatifs