Retention and Leaching of Elevated N Deposition in a Forest Ecosystem with Gleysols
24
0
0
Texte intégral
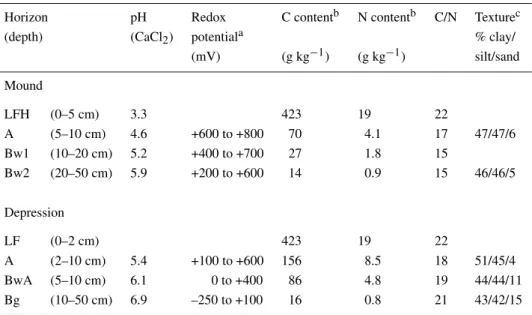
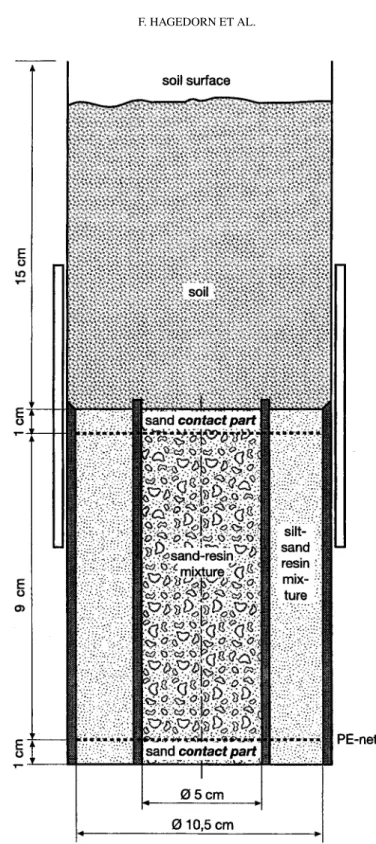
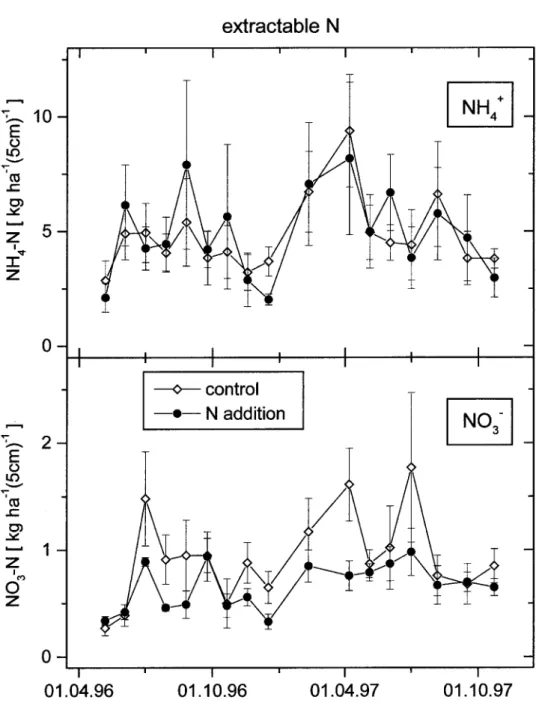
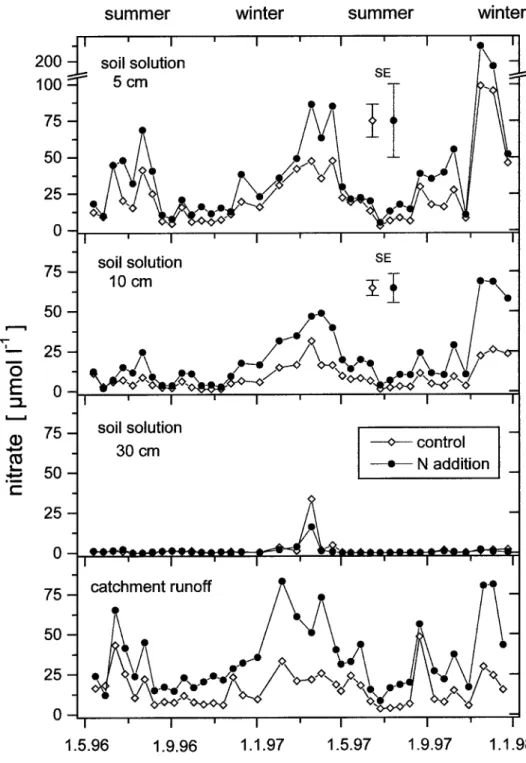
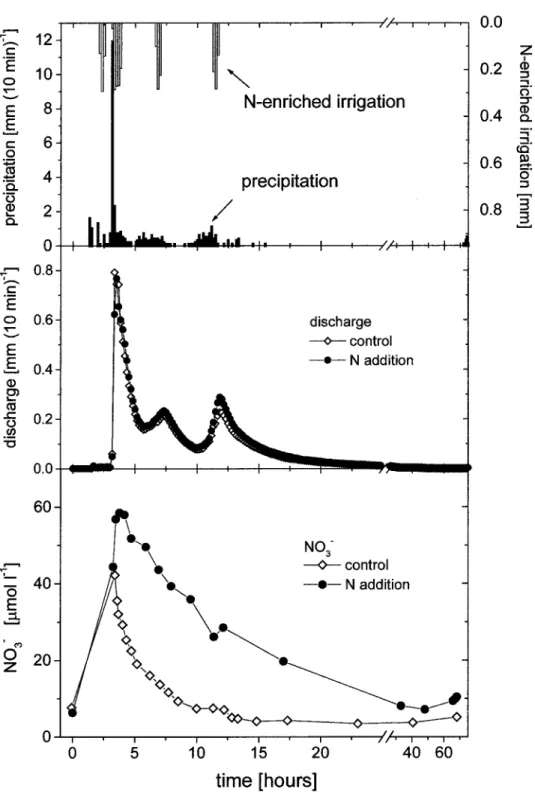
Figure
+7
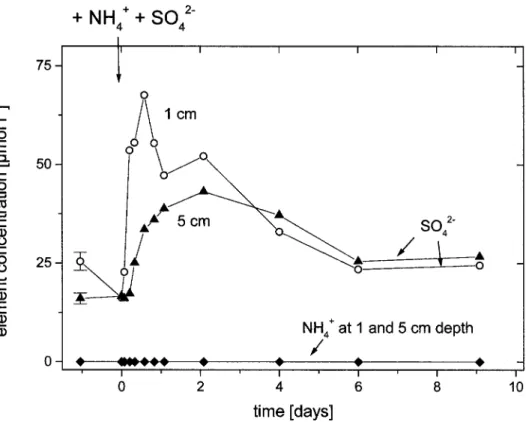
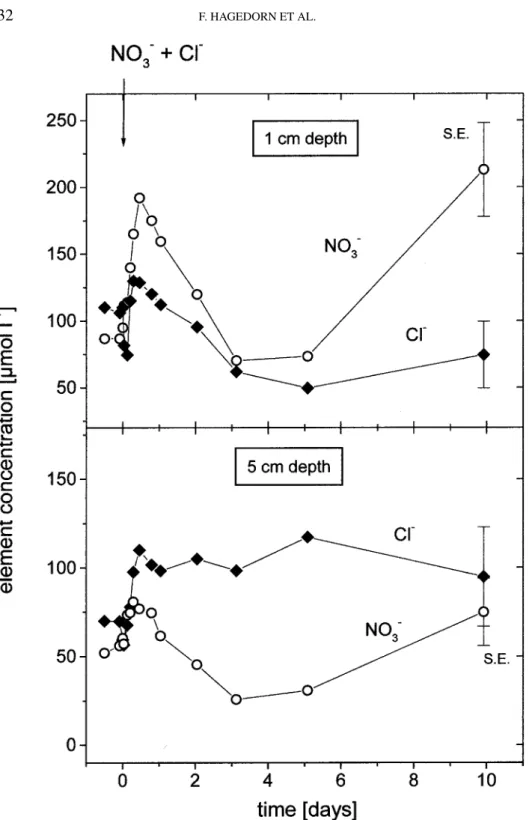
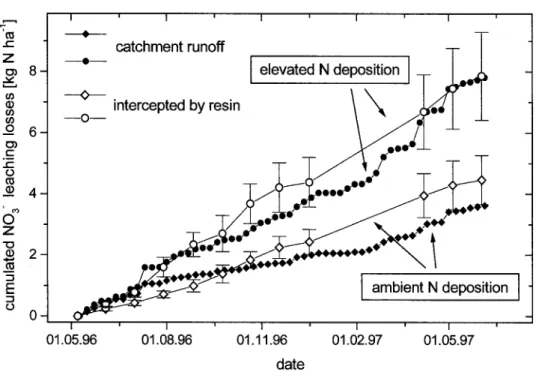

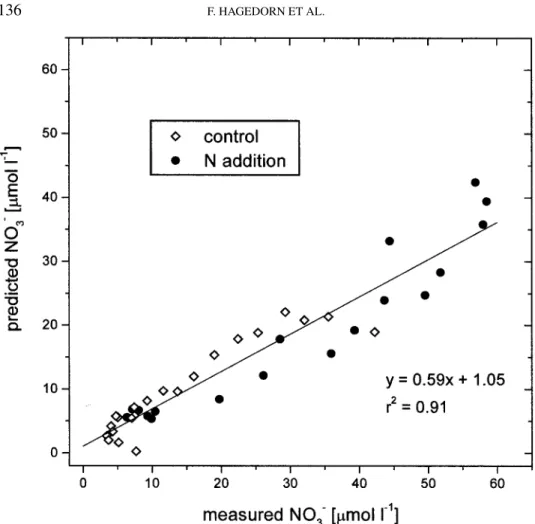
Documents relatifs