HAL Id: hal-02044588
https://hal.archives-ouvertes.fr/hal-02044588
Submitted on 2 Apr 2019
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Localisation of two candidate genes for mental
retardation using a YAC physical map of the
Xq21.1-21.2 subbands
Laurence Colleaux, Melanie May, Jérôme Belougne, Denis Lepaslier, Charles
Schwartz, Michel Fontes
To cite this version:
Laurence Colleaux, Melanie May, Jérôme Belougne, Denis Lepaslier, Charles Schwartz, et al.. Local-isation of two candidate genes for mental retardation using a YAC physical map of the Xq21.1-21.2 subbands. Journal of Medical Genetics, BMJ Publishing Group, 1996, 33, pp.353 - 357. �hal-02044588�
IJMedGenet1996;33:353-357
Original articles
Localisation of
two
candidate
genes
for
mental
retardation
using
a
YAC
physical
map
of
the
Xq21.1-21.2
subbands
Laurence Colleaux, Melanie May, Jerome
Belougne,
Denis Lepaslier,Charles Schwartz, Michel Fontes
Abstract
Genetic studies in families with X linked mental retardation havesuggestedthe loc-ation ofseveral MR genes in the humanq21 region. Since the establishment of cloned resourcesis an essential step towards the cloning of genes involved in inherited dis-eases, we built a yeast artificial chro-mosome (YAC)contigandan STS map of this part ofthe Xchromosome. Thecontig,
which extends from PGK1 in Xql3.3 to
DXS1002 in Xq21.2, consists of 30 YACs
mappedwith21 markers and spans about
6Mb. TheYACcontigwasusedas a frame-workto localise severalpreviously known genes and CEPH/Genethon polymorphic markers, aswellas toconstruct aphysical
mapoftheregionsurroundingoneofthese genes. We recently localised a presumed
MRlocus to theregion flankedbyDXS233
(proximal) andCHM (distal).Inthe
pres-entwork, the zinc fingergene, ZNF6,has been shown to lie within this region and
tobehighly expressedinbrain, makingit
a good candidate MR gene. Similarly the VDAC1 gene has been mapped between DXS986andDXS72anditscandidategene
statusfor theAllan-Herndon-Dudley
syn-dromeis discussed.
(J7MedGenet 1996;33:353-357)
Key words: Xq21; mentalretardation; YAC contig.
The dark staining bandXq21 is assumed to
be
aregionin which there arefewgenes.However,
there is strong evidence that this region may
contain genes involved in brain function. For
example, chromosomalrearrangementssuch as
deletions, microduplications, or inversions of part of this chromosomal region have been reported as being associated with mental
re-tardation.1 Invariably,inmales with interstitial deletions of the Xq21 band, the mental re-tardation is associated with some other entity,
usually choroideremia (CHM), but sometimes both CHM and deafness
(DFN3).125-7
Other strong evidence for the presence of a gene or genes in Xq21 involved in mental retardation conditions comes from the fact that some Xlinkedmentalretardation (XLMR) syndromes, suchastheMiles-Carpenter,8 Goldblatt,9
Cow-chock-Fishbeck,9 or Allan-Herndon-Dudley
syndromes,'0 1'aswellasseveralnon-syndromic
MR entities such as MRX7,'2 MRX8,'3 or
MRX31,1'
exhibit maximal lod scores with markers in this region. Characterisation of the deletion of Xq21 patients with overlapping phenotypeshas recently allowed us tolocalisea gene involved in MR within the DXS233/ CHM interval.'5 Using newly isolated cDNA clonestoscreenformutations in patients with
some forms of XLMR linked to the 2-2 Mb
Xql3.3region between DXS441 and
PGKI,'6
wehave identified the XNP (Xlinked nuclear protein) geneasbeing responsible for the ATR-X syndrome.'7 Based on the success of these combined analyses, we decided to conduct
physical mapping studies on theXq21 region.
Previous analyses have focused mainly on the ordering of probes using a panel of deletion
ortranslocation patients.'8 A report of PFGE experiments, conducted withsomeprobes from theregion, has been published.'9 However, no
cloned resources,encompassing at least part of the interval between PGK1 and DXS1002, have been describedso far, neither hasa com-plete map including physical distances. We therefore decided to cover this region with YACs and to linkthis newcontig to our already existing YAC contig around PGK1. This was
accomplished using an STS basedapproachto
isolateMegaYACs from the CEPHlibrary and resulted in the generation of a 6 Mb YAC contig.
This YAC contig was then used to localise precisely several previously known genes and to build a long range restriction map of the DXS995-DXS1002 region. Our results show that the MR critical region between DXS233 andpJ7.6A'5 isnolarger than 1-2 Mb and that the zinc finger gene ZNF6 lies within this
region.We alsoidentified,within the same MR critical region, a new potential CpG island whichmight be indicative of at least a second
transcription unit within this MR critical
re-gion. Moreover, the resources we have pro-duced allowedus tolocalise another previously known gene, the VDAC 1 gene,20 between DXS986 and DXS72. This result, in com-bination with the potential function of the VDAC1 protein, makes the VDAC1 gene a
good candidateforbeing involvedinthe
Allan-Herndon-Dudley
syndrome. INSERMU406, Faculte deMedecine de LaTimone, 27BlvdJean Moulin, 13385Marseille Cedex 5,France LColleaux JBelougne MFontes Greenwood Genetic Center,Greenwood,SouthCarolina,USA
M May CSchwartz Centred'Etudedu Polymorphisme Humain, Paris, France DLepaslier Correspondenceto: DrColleaux. Received14November1995
Revisedversionacceptedfor publication9January1996
Colleaux, May, Belougne,Lepaslier, Schwartz, Fontes
TableI Previouslyunpublished STSs used to construct the contig
Name Probe Primersequence Product Temp size (bp) DXS571 pHX79 AATATTGGTGCAGGCACTGT 129 55°C
AATCAGATGCAGTGATGGGT
DXS447 TCCACACCGACTTGTATTGC 226 57°C T-l'ACCAGCTTGGATGAGGC
VDAC1 cDNA TGAACAAGAAGTTGGAGACCG 208 60°C
TTCTTGCCATCCAGAAGAGC DXS72 pX65H7 CAATTTTlCTGCCAATTAGGATC 221 55°C GTAGTTGACTTCGAGCTGGC ZNF6 5' cDNA ACTCTTGATCATGGCCTTGC 228 56°C CAGTAACAAGGTCAGCCACG 3'cDNA AGCATTTGGTGATGAGAGGG 244 56°C CCTGCACGTATGAATCTTCCTAC CHM cDNA CACCAAACTGGATACCCA 237 65°C TGAGCAAGTCAATGTGCT
The primer sequences are given in a 5'to 3' orientation. When the primers derive from a previously published probe, the name of the corresponding probe is indicated. cDNA: primer design according to the cDNA sequence corresponding to the gene.
Methods
YAC LIBRARY SCREENING
Pools of the CEPH MegaYAC library were
screenedby PCR for thepresenceof DXS571,
DXS447, and DXS72 using the primers and
the annealing conditions shown in table 1.
Screening for locus DXS326 was carried out
using the primers described in Barker and
Fain.2"
The PCR conditionswere 96°C for 30seconds, one minute at the annealing tem-perature,and720C for40secondsfor40cycles.
LONG RANGE RESTRICTION MAPPING
YAC DNAembeddedin agarose wasdigested
to completion with either
SfiI,
NotI, MluI, orEagI using the buffer supplied by the
manu-Table2 STSs andprobecontentofYACs betweenPGKI andDXS1002
Six(kb) S I DXS1225 DXS571 DXS447 DXS11971DXS986 |VDACI IDXS72 IDXS169 DXS, 326b 0 330 b _ 95fl 1640kb __ _= 792a5 1520 k __ = = = 910hl 170kI_ - - = = 9411a2 I79 kb _-= = = =_ 657e12 480kb_ = 921 10 1230kb__= 829bl2 ND = = = 813bl0 ND_
11540
7 1250 kb== __=-946>^8 1360 kb 927b5 0kb 967el2 1140 kb _ _ 90402 770kb_ _ _ = _ = _ X21alO ISXOkh 955a12 1300 kb I <I = <1 i 321f8 813a2 E1023 1025kh 5OOkb 632b10 710kb -659h3 490kb -666a6 710 kb -I-=-=--= = = 668e6 830 kb I 7253 1100kb -9160 1400kkb -979aO10 1600kb -I - -.. -I909 4-25-k --i --i-- i-li-i- - i i 922b5 1350kb _
S26 DXS995 Pu3f4
facturer
(New England Biolabs).
Thefrag-ments were then electrophoresed on a 1%
agarose
(Gibco BRL)
gel in a CHEF DRIIPFGE apparatus (Biorad) using switch times from 40 seconds to 120 seconds at 200V for
30 to 38 hours in 025xTBE at 12°C. The
DNA was then transferred to nylon Biodyne
membranes (Pall Industries).
PROBE PREPARATION AND HYBRIDISATION
CONDITIONS
Probeswerelabelledusing the randompriming
procedure as previously described.22 Southern
blothybridisationswerecarriedoutina
stand-ard SSC buffer (5x
SSC,
5xDenhardt's, and01% SDS) for 16 to 18 hours at 65°C in a
rotary hybridisation oven. Washes were
per-formedasfollows: 20 minutes in 2xSSC/0 1%
SDS, 20minutes in 0 5xSSC/0-1%SDS,and
five to 20 minutes in 0 1 xSSC/0-1% SDS
according to the stringency of the washes. A
Northernblotfilterwaspurchasedfrom
Clon-tech and hybridised according to the
manu-facturer's recommendation. All membranes
wereexposedtoX-OMAT AR film(KODAK)
from one hourtofive
days.
Results
ISOLATION OF YAC CLONES AND CONTIG
CONSTRUCTION
Asapreliminarysteptowards theconstruction ofa YAC contig encompassing the Xq21
re-IID 2 IX IN D*S100tXI
-i - i - - - T T
-I I I i I tI
I
Theupperline indicates theSTS/probemarkers used to build the YACcontigand their order as itappearson thecontig map (fig 1).The size of the YACshas
been determinedaccordingtothe CEPH-GENETHONdatabase or in thisstudy.ND: notdetermined. .L'±I13 kt - -I I - - - I - --- 1-, I
i
i I 354 ND ..I.n IsA so16 ....,.T
Localisationoftwocandidate genesformental retardationusinga YACphysicalmapoftheXq21.1-21.2 subbands Xcen Xtel DXS232 DXS233 DXS995 DXS 121 DXS355 N m m m
=1
m=
ES MS MS E S M E ESM ,, ZNF6 DXS326 +___ -- ME3'S
E5'
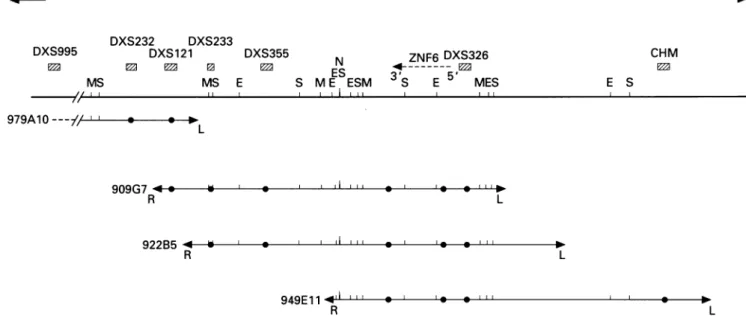
MES L 909G7 -R 922B5 L III 949E11 -" *I I 0 0 I NO. R LFigure1 Longrangerestrictionmapofthe DXS232-CHMregion. LocationoftheXq21 referenceloci is indicatedbyhatchedboxes. Restrictionsites
areE=EagI, M=MluI, N=NotI, and S=SfiI. Filled circlesrepresentthe YAC restrictionfragmenthybridisingwith theprobeindicated above. The
probe DXS326and theprobecorrespondingtothe 5' endofthe ZNF6genehave beenarbitrarilyorderedas ourresults donotallowordering ofonewith
respect tothe other.
gion, the CEPH MegaYACs library was
screened for the loci DXS571, DXS447, DXS72,andDXS326. Three DXS571 positive YACs (813blO, 829b12, 921glO), two DXS447positiveYACs (854e7,967e12),three DXS72positive YACs(767e4,821alO,321f8),
and three DXS326 positive YACs (909g7,
922b5, 949el1) were characterised at this
stage. The CEPH-Genethon database23 was
thenusedtoidentifyYACs that either mapped totheregiononthe basisoftheirSTScontents (six YACs positive for DXS995 (632b10,
659h3, 666a6, 668e6, 725b3, and894c7),one
additional YACpositive forDXS571 (657e 12),
oneYAC positive for DXS1225 (910hl), and
two YACs positive for DXS986 (946e8, 927b5)) orappearedtocontig withYACs that hadalready been placed in the contig (95 1fl 1, 792a5, 948a2 that werein contig with 910hl
and 749h4, 955al2, 904f2, 838c5 that were
in contig with 821alO). In total, 28 CEPH MegaYACs were further studied with 21
markers includingfivegenes.
TheindividualSTSorprobecontentofeach YAC was then determined by the combined use of PCR and hybridisation. In addition to the previously published original set ofSTSs from this region, several additional ones were
developed. They are listed in table 1 and the
STS dataare summarised intable 2. This led
to theassembly oftwo contigs spanning from PGK1 to DXS26 and from DXS995 to
DXS 1002. Since an 850kb YAC and cosmid
contig encompassing the DFN3 locus and link-ing the two loci DXS26 and DXS995 had
previously been described,24 two additional
YACs fromtheICRFlibrary, ICRFy9O1E 1023 and ICRFy900B1210 respectively, were also
studied andincluded in the finalunique contig.
RESTRICTION MAPPING OF THE MR CRITICAL REGION AND ESTIMATION OF THE CONTIG SIZE
Asdescribedin theintroduction,we
previously
localisedtheproximalanddistal borders of the candidate
region
for one MR gene in Xq21 between DXS233andCHMrespectively.'5Wetherefore decidedtoconstructarestrictionmap
of this
region
to orderclosely
spacedmarkers and estimate the true distances between loci.Mapping was conducted using the four rare
cutterendonucleases
SfiI, MluI, NotI,
andEagI
and the YACs 979a10, 909g7, 922b5, and 949e1 1. Themap wasassembledusingprobesderived from the left and right ends of the
pYAC4 vector, as well as internal probes
cor-respondingtoSTSsor genes.Eventually,
mem-braneswerehybridised with total humanDNA
toensurethat all restriction fragments had been
detected. The result is shown in fig 1. Using
this map, itwaspossible toorderanumberof
markersfrom the region. From thecentromere to the telomere, the following order was
found: DXS232-DXS121-DXS233-DXS355-(DXS326, DXS1167, ZNF6)-CHM-DXS1002-DXS95. This is in agreement with
data previously obtained by other groups.'825
Analysis of the rare cutter restriction map of
theregion showed atleast one cluster ofsites
containingaNotI site, which couldcorrespond
to a CpG island, approximately 400kb distal
to DXS233, betweenDXS355 and DXS326.
Previous studies26 have shown that the human ZNF6 gene is localised between
DXS233 and CHM. A probe specific for the 5' end of the cDNA, hybridised to a 300kb
SfiI
fragment, a 400kbMluI fragment, and a200 kbEagI fragment, positioningthat end of the gene approximately 800 kb distal to
DXS233.A probecorrespondingtothe3' end
CHM
E S
355
Colleaux, May, Belougne, Lepaslier, Schwartz, Fontes
approximately 3-8 Mb between DXS447 and
n;)
rL)
1~t]
!_
t
DXS1002.
According to our long rangere-striction maps, theminimumdistancesbetween DXS995-DXS355 and between
VDAC1-DXS 169 are about 1 Mb and 400 kb
re-ki) spectively.The size of themost proximal YACs (326b 10,
910h1,
657e12) andtheirchimerismstatus
according
to the CEPH databases sug-7^>\-ii* * i gest that the distance between PGK1 andDXS447 should be about
600
kb.Finally,
aprevious publication
reported
the distancebe-4.4 tp
kinltween
DXS26 andsuit DXS995p t to be 250kb.24Therefore, considering all these data, we es-timate the size of ourcontig to be about 6 Mb
fromPGK1 to DXS1002.
2.4
Ibactin
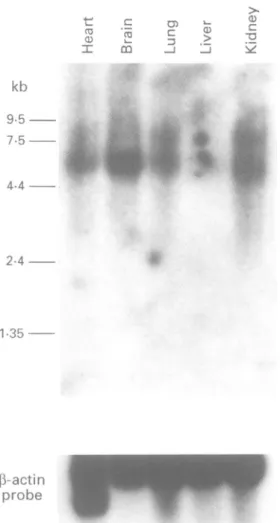
profbeFigure2 Expression analysis of theZNF6gene. A humanmultiplefetaltissuenorthern blot(Clontech) was
hybridisedwith aprobe correspondingtothe ZNF6gene.
Theprobeidentifies a5 5kbtranscripthighly expressedin
brain,lessabundantlyinheart, lung, andkidney,and almostundetectable in liver. Thesameblotwasalso
hybridisedwith the actinprobetodetermine the relative
quantities ofRNA loadedperlane.
of thegenehybridisedtoa200kb
Sfi1
fragmentand a350 kb EagI fragment. Accordingto the
map (fig 1), this result indicates that the 5'
specific probe hybridises to a fragment distal
to the one hybridised by the 3' end, thereby
establishing thatZNF6 istranscribedtelomere
to centromere and is localised between
DXS355 and DXS326.
Recentstudies havemapped thehumangene
encodingthemitochondrialvoltagedependent anionchannelgene,VDAC1,20intotheXql
3-q21 interval. We therefore decided to localise
thisgenepreciselyonthe YAC contig byPCR
amplification to associated STSs. The results
show thatitliesbetween DXS986 and DXS72.
Similarly to whatwe did for the ZNF6 gene,
the use of specific 5' end or 3' end specific
probes and preliminary data on the physical
map of the region surrounding the VDAC1 gene allowed us to determine that it is
ori-entatedfrom the telomeretothe centromere. Finally,toapproximatethesize of thecontig
we combined the chimerism/size data
de-termined either by ourselves or through the
CEPH-Genethon database. The three
non-overlapping, non-chimeric YACs, 946e8, 813a2, and 949ell, give a minimum size of
EXPRESSION STUDIES OF THE ZNF6 GENE
Since the ZNF6 gene, which maps between DXS233 and CHM, encodes for a putative
zinc finger transcriptional factor, its position and putative function led us to speculate that a deficiency in the expression ofsuch a gene
could be involved in a form of mental
re-tardation.Inordertostudy its candidategene status, we performed a
northern
blotex-periment,using RNAsextractedfrom avariety
of fetal tissues (fig 2). A 5-5 kb transcriptwas
identified and showed that the ZNF6 gene
is highly expressed in fetal brain with lower
expression inheart, kidney, and lung andvery low expressioninliver. In addition to this 5-5 kb
transcript, alarger one, about 7-5 kb, was also
identified in kidney and lung. Whether this
largerfragment correspondsto an
alternatively
spliced transcript remains to be determined; however, our finding that the ZNF6 gene is
highlyexpressedinbrain makesit agood
can-didateforbeing involvedin the MRsyndrome.
Discussion
Thepresentworkdescribesthegeneration ofa 6MbYACcontigspanningthegenomic region
between PGK1 in
Xql3.3
and DXS1002 inXq21.2.
When
we started this work, a small YAC contig linking DXS26 and DXS995 had beendescribed,24
and pulse field electro-phoresisanalysis
ofgenomic
DNAhad shown the closeproximity of loci DXS232, DXS121, andDXS233.'
However,despite
these data,the
region
runningfromDXS447toDXS1002 wassplitinto manycontigs and severalprobesorSTSs, suchasDXS326,
DXS355,
orZNF6,
were notincluded in anycontig.25
Thecontig
we havebuilt contains 30 YACs and21 STSshave beenordered within this contig. Because
the STS order we deduced from our YAC
contigisconsistentwith the
ordering
ofprobes
resultingfrom theuseofa
panel
ofdeletion ortranslocation
patients,'8
we think that these YACs are representative of the corresponding genomic region and shouldprovide
usefulclonedresources forfurther
analysis.
Since our
previous
resultsconcerning
themolecular analysis of males with mental
re-tardation and deletions of Xq21
placed
theputative MR region between DXS233 and
CHM,'5
we builtalong rangerestrictionmap of the DXS232toDXS1002 interval. It shows 356Localisation oftwocandidategenesfor mental retardationusinga YACphysicalmapoftheXq21.1-21.2subbands
that the critical region for the MR gene, be-tween DXS233 and pJ7.6A, is about 1 2 Mb.
Interestingly,ourphysicalmapis ingood agree-mentwiththe data obtainedby Merryetal' on
the longrange restriction map ofthegenomic
regioncontainingDXS232andDXS233,
sug-gestingthat theseYACsarefully representative
of thegenomic region analysed.Twocandidate
MR genes, based upon their chromosomal
localisation, have already been characterised.
Thefirstone,the ZNF6gene,codesforazinc
finger transcriptional factor and is localised
betweenmarkers DXS355 and DXS326. Our
studyonpatientswithcontiguousdeletion
syn-dromesshowthatitis localisedwithinthe MR
critical region,'5 being deleted in all patients
with MR associated with CHM only or with
CHM anddeafness,butpresentintwopatients
with only CHM. Its candidate gene status is
strongly supported by the results ofnorthern
blotexperimentswhichshowedhigh expression
infetalbrain. Wealso characterisedapotential new CpG island within this critical region.
Since the construction of the physical map of
this region has allowed us to orientate the
ZNF6 gene as cen-3'-ZNF6-5'-tel, this new
CpG island could possibly be an island
in-dicative of a new transcription unit. cDNA
selectionexperimentsareatpresent being
per-formedtotry to isolatethis newgene.
Thesecondcandidategene,VDAC1, is
loc-alised close to DXS72. Since recent findings
have shown that abnormalities in genes
en-codingthesechannelscause anumber of
neuro-logical disorders in mice27 and men,28 the
VDAC1 gene may well be involved in neuro-muscular disorders, most notably the Allan-Herndon-Dudley syndrome. Indeed, this
syn-drome,which ischaracterisedbyseveremental
retardation and spastic paraplegia, has been
localised within DXS571 and DXS326 with
a lod score of about 7 (Schwartz et al, in preparation). Furthermore, amongthe numer-ous maleviable deletions spanning almost the
entire Xq21 region studied, most areonly as-sociated with a mild MR condition. Only
patients deleted proximal to DXS72 exhibit
severe mental retardation.6 Thus, one could
speculate that the AHD gene is very likely
localised upstream of this locus, between
DXS571 and DXS72, making the VDAC1
geneaverygoodcandidategeneforthe
Allan-Hermdon-Dudley syndrome. As illustrated by
the recent finding, resulting from the com-bination ofpositional cloning and candidate
gene approaches, that the POU domain gene
BRN4 is implicated in X linked mixed
deaf-ness,29the physical map reported here should
greatly facilitatethe identification ofgenes
in-volved in XLMR as well as othergenetic
dis-eases assignedtothispart ofthe chromosome.
Using the YACclones describedhere,workis
now under way to create an extensive
tran-scriptional map oftheXq2 1region.
LC,JB,and MFweresupportedbyagrantfromtheAssociation
Francaise contre les Myopaties (AFM). MM and CS were supported, in part, by agrant from theSouth Carolina De-partment ofDisabilitiesand SpecialNeeds and agrant from
NICHD.
1 MerryDE, LesloJG, SosnoskiDM,etal. Choroideremia and deafness withstapes fixation:acontigousgenedeletion
syndromeinXq21. Am_7HumnGenet 1989;45:530-40.
2 SchwartzM,Rosenberg T,NiebuhrE,etal.Choroideremia: further evidence forassignment of the locus to Xql
3-Xq21.Hum7n Genet1986;74:449-52.
3 Abeliovitch D, Dagan J, Kimchi-Sarfati C, Zlotogora J.
ParacentricinversionX(q21.2q24)associated with mental retardation in males and normal ovarian function in fe-males.Am]_MedGenet1995;55:359-62.
4 Yokoyama Y, Narahara K,TsujiK,etal. Growthhormone deficiency and empty sella syndrome in a boy with
dup(X)(q13.3-q21.2).Anm Med Genet1992;42:660-4.
5 NussbaumRL, LeskoJG,Lewis RA,LedbetterSA, Led-betterDH.Isolation of anonymoussequencesfromwithin a submicroscopic X-chromosome deletion in a patient
withchoroideremia,deafness and mental retardation. Proc Natl Acad Sci USA 1987;84:6521-5.
6 CremersFPM,vande PolDJR,Diergaarde PJ,etal.Physical
mapping of the choroideremia locususingXq21deletions associated with complex syndromes. Genomics 1989;4:
41-6.
7 Hodgson SV, Robertson ME, Fear CN, et al. Prenatal
diagnosis of X-linked choroideremia with mental
re-tardation, associated withacytologicallydetected
X-chro-mosomedeletion. Hum Genet 1987;75:286-90.
8 CarpenterNJ,WaziriM,ListonJ,Patil SR. Studieson X-linked mental retardation: evidence forageneintheregion
Xql 1-q22.AmIHum Genet 1988;43:A139.
9 NeriG, ChiurazziP,ArenaJF, Lubs HA. XLMRgenes:
update 1994.Ami] Med Genet1994;51:542-9.
10 Schwartz CE, UlmerJ, BrownA, Pancoast I, Goodman HO, Stevenson RE.Allan-Hermdonsyndrome.II.Linkage
to DNA marker inXq21. Anm 7 Hum Genet 1990;47:
454-8.
11 Schwartz CE, Martin J, Ouzts L, Arena JF, Lubs AH,
Stevenson RE.Allan-Hermdon-Dudleysyndrome,linkage
analysis inathirdfamily and refinement of the localization inXq21.Cytogenet Cell Genet 1994;67:351.
12JedeleKB, MichelsVV,Wagner KV.Linkageanalysis dem-onstrating association of DXYS 1 andnonspecificX-linked mental retardation.Ami ]fHumi Genet1990;47:A62.
13 SchwartzCE,May M,Huang T,etal.MRX8:anX-linked mental retardation condition withlinkagetoXq21.Amn_7
Med Genet1992;43:467-74.
14 PartingtonMW, Ryan A, DonnellyAJ, MulleyJC. Two families with X-linked mental retardation(XLMR)from Queensland. An_7Med Genet(inpress).
15 May M, Colleaux L, Murgia A,etal. Molecularanalysis of four males with mental retardation and deletions ofXq21
places theputative MRregion inXq21.1 betweenDXS233 and CHM.Humn Mol Genet1995;4:1465-6.
16 VillardL, GeczJ, ColleauxL,etal. Construction ofaYAC contigspanning theXql3.3subband. Genomics1995;26:
115-22.
17 GibbonsRJ, Picketts DJ, VillardL, Higgs DR. Mutations in a putative global transcriptionalregulator causes X-linked mental retardationwith y-thalassemia(ATR-X syn-drome). Cell 1995;80:837-45.
18 Philippe C, Amould C, SloanF, etal. Ahigh-resolution
interval map of the q21 region of the human X chro-mosome.Genontics1995;27:539-43.
19 CremersFPM, SankilaEM, BrunsmannF,etal.Deletions inpatients with classical choroideremia vary in size from 45 kb to severalmegabases. Am_7Humn Genet 1990;47:
622-8.
20 Blachly-DysonE, Baldini A, LittM, McCabeER, Forte M. Human genes encoding thevoltage-dependent anion channel(VDAC) of the outer mitochondrial membrane: mapping and identification of two new isoforms.Genonmics
1994;20:62-7.
21 BarkerDF, Fain PR. Definition and mapping of STSsat
STR and RFLP loci inXpll-Xq22. Genomics 1993;18:
712-16.
22 FeinbergAP, Vogelstein B. A technique forradiolabelling
DNA restriction endonucleasefragmentstohighspecific
activity. Anal Biochemn1984;137:266-7.
23 CohenD, Chumakov I, Weissenbach J. A firstgeneration physical map of the human genome. Nature 1993,366: 698-701.
24 HuberI,Bitner-GlindziczM, de KokYJM,etal.X-linked mixeddeafness(DFN3): cloning and characterization of the critical region allows identification of novel mi-crodeletions. Hunm Mol Genet1994;3:1151-4.
25 Willard HF, Cremers F, Mandel JL, Monaco AP, Nelson DL, Schlessinger D. Report of the fifth international
workshoponhuman Xchromosome mapping 1994.
Cyto-genet Cell Geniet1994;67:295-358.
26 LloydSL, Sargent CA, Chalmers J, Lim E, HabeebuSSM,
Affara NA. An X-linked zinc finger gene mapping to
Xq21.1-q21.3 closelyrelatedtoZFXandZFY: possible
origins from common ancestralgene. Nucleic Acids Res 1991;19:4835-41.
27 Nakatsu Y, Tindale RF, DeLorey TM, et al. A cluster of three GABAAreceptor subunit genes is deleted ina
neurologicalmutantof themouseplocus. Nature1993;
364:448-50.
28 Shiang R, Ryan SG, Zhu YZ, Hahn AF, O'Connell P,
WasmuthJJ.Mutationsinthe a,subunit of theinhibitory
glycinereceptorcausethedominantneurologicaldisorder hyperekplexia.NatureGenet1993;5:351-8.
29 de KokYJM, vander Maarel S, Bitner-Glindzicz M, et al.Association between X-linked mixed deafness and mutations in the POU domain gene POU3F4. Science 1995;267:685-8.