Grass Physiognomic Trait Variation in African Herbaceous Biomes
Texte intégral
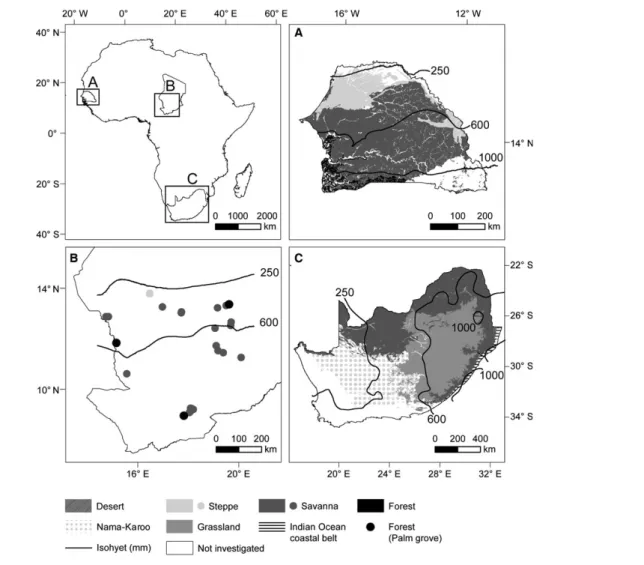
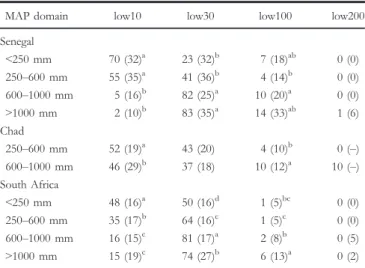
Figure

Documents relatifs
Concernant l’impact sur le système racinaire, l’indice de stress hydrique est estimé pour chaque pointe racinaire en fonction de la FTSW locale (la FTSW dans le volume de sol où
In partnership with Patuchev experimental set-up (UE FERlus - INRA Lusignan – France), biochemical (overall composition, fatty acids) and microbiological characteristics (lactic
Can't say that I understand what they did, but I can understand that there were lots of reasons why people may have been inclined or tempted to abandon their
Tous les plus grands classiques de la culture littéraire européenne sont là : Rabelais et Cervantès, ou encore Laurence Sterne, dont les imaginaires grotesques peuplent à
The group SO(n), as well as other groups of linear transformations, is usually not defined in abstract by characterizing its elements g, but specifying the properties of one
The cirques and ramparts on Réunion Island have been listed as part of the world cultural and natural heritage since 2010 and the invasive alien species (IAS) are
Also known as: Andean pampas grass, purple pampas grass, Andes grass, Cortaderia, jubata grass, pampas grass, pink pampas.. WHY IS IT
Sown grass-margin strips, which have typically been established to limit pesticide drift and soil erosion, are now also promoted for their positive, ecological impact on
