Conserved domains control heterochromatin localization and silencing properties of SU(VAR)3-7
12
0
0
Texte intégral
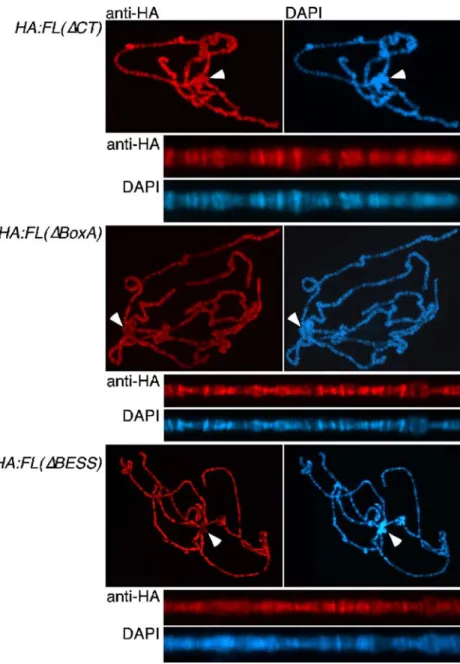
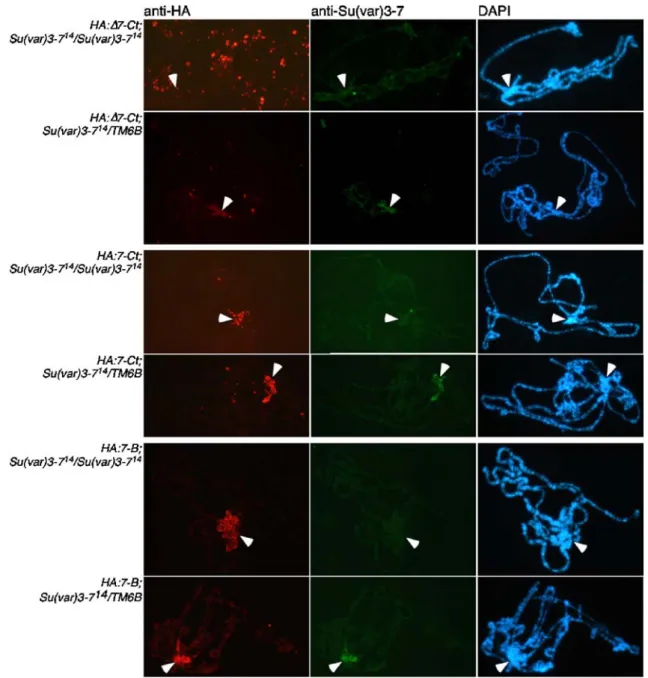
Figure


Documents relatifs
