Shear stress affects the architecture and cohesion of Chlorella vulgaris biofilms
12
0
0
Texte intégral
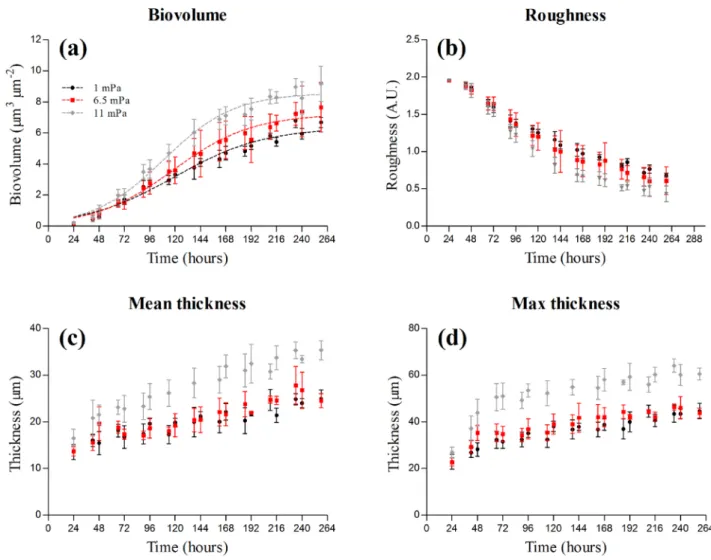
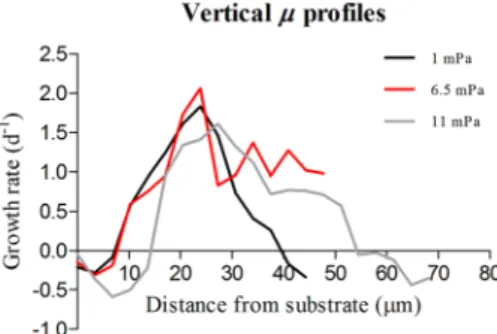
Figure

Documents relatifs