HAL Id: hal-02786889
https://hal.inrae.fr/hal-02786889
Submitted on 5 Jun 2020
HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Effets de la fauche et du réchauffement climatique sur la
diversité végétale dans les prairies plurispécifiques :
approche par méta-analyse
Justin Badeau
To cite this version:
Justin Badeau. Effets de la fauche et du réchauffement climatique sur la diversité végétale dans les prairies plurispécifiques : approche par méta-analyse. Milieux et Changements globaux. 2018. �hal-02786889�
1
2017 – 2018
MASTER FAGE
Biologie et Ecologie pour la Forêt,
l’Agronomie et l’Environnement
Spécialité
FGE
Effets de la fauche et du réchauffement climatique sur la
diversité végétale dans les prairies plurispécifiques : approche
par méta-analyse
JUSTIN BADEAU
Mémoire de stage, soutenu à Nancy le 04/09/2018
Catherine, PICON – COCHARD, directrice de recherche UREP
Alice, MICHELOT – ANTALIK, maître de conférence LAE
UMR 0874 Ecosystème prairial (UREP) – INRA, Site de Crouël
5 chemin de Beaulieu
2
REMERCIEMENTS
Je tiens à remercie tout particulièrement Catherine Picon-Cochard pour sa sympathie et toute l'aide apportée dans la réflexion sur le sujet et la rédaction du rapport.
Je remercie aussi Gianni Bellocchi pour m'avoir initié à l'utilisation des modèles en écologie ainsi que pour l'aide qu'il m'a apporté durant toute la durée du stage.
3
SOMMAIRE
I. INTRODUCTION 1
II. ETAT DE L'ART 2
A. Définitions 2
B. Réponses de la diversité végétale aux facteurs de gestion 4 C. Réponses de la diversité végétale aux facteurs climatiques 6
D. La méta-analyse 8
E. Objectifs 8
III. MATERIELS et METHODES 9
A. Documentation 9
a. Recherche des sources bibliographiques 9
b. Sélection des articles pertinents 10
c. Création des bases de données 10
B. Analyses statistiques 10
a. Extraction des données 10
b. Traitement des données 11
IV. RESULTATS 13
A. Méta-analyse sur les effets de la fauche sur la diversité végétale des prairies 13
a. Schéma Prisma 13
b. Résultats pondérés et non pondérés 13
c. Modèles à effet aléatoire et à effet mixte 14
B. Méta-analyse effets du réchauffement sur la diversité végétale des prairies 16
a. Schéma Prisma 16
b. Modèles à effet aléatoire et à effet mixte 17
V. DISCUSSION 18
A. Effets de la fauche 18
B. Effets du réchauffement 20
C. Analyses de l'hétérogénéité 22
VI. CONCLUSION 22
VII. REFERENCES BIBLIOGRAPHIQUES 23
ANNEXES RESUME
4
LISTE DES ABREVIATIONS
LRR : Logarithme du ratio de réponse MEA: millenium ecosystem assessment r : coefficient de corrélation linéairer² : coefficient de détermination (coefficient de corrélation élevé au carré) SD: écart-type
𝑆𝐸: 𝑒𝑟𝑟𝑒𝑢𝑟 𝑠𝑡𝑎𝑛𝑑𝑎𝑟𝑑 = 𝑠𝑑 √𝑛
1
I.
INTRODUCTION
Les prairies permanentes sont essentielles à la production de fourrage et permettent un ensemble de services écosystémiques indispensables à nos sociétés. Par exemple, elles contribuent au stockage du carbone dans les sols et limitent les émissions de gaz à effet de serre. Les prairies permanentes sont caractérisées par une biodiversité élevée composée de végétaux, mammifères, arthropodes et micro-organismes. Cette biodiversité permet la pollinisation et le maintien de l’esthétique générale des paysages, mais également est reconnue comme étant une assurance écologique et évolutive grâce aux changements phénotypiques au sein des espèces végétales et aux changements d’abondance de celles-ci. La biodiversité rendrait les prairies plus résilientes face aux aléas et aux évènements climatiques extrêmes (comme les sécheresses de longue durée) et serait à même de stabiliser la production de fourrage et le maintien de l’ensemble des services écosystémiques. Il est donc essentiel de conserver ces espaces ouverts afin de préserver cette biodiversité et les services associés mais aussi de les étudier pour mieux apprécier leurs évolutions. Certains mécanismes écologiques sous-jacents au fonctionnement des prairies sont pourtant encore mal compris. Ainsi, déterminer la nature et la valeur des biens et services que les prairies peuvent fournir dans des conditions environnementales et de gestion différentes reste un domaine de recherche ouvert et encore peu exploité.
Les services pouvant être rendus par les prairies dépendent en partie de la diversité de leurs cortèges floristiques. Les changements de la diversité végétale reflètent l’évolution de deux facteurs principaux : les conditions environnementales (pédo-climat) et les pratiques de gestion (fauche, fertilisation, pâturage,). Le développement des prairies est donc influencé par de nombreux éléments et il est ainsi difficile de prévoir avec précision les dynamiques d’évolution des espèces végétales composant les couverts prairiaux. Le projet MODIPRAS (modélisation des relations entre la diversité spécifique, le fonctionnement des systèmes prairiaux et leur capacité à fournir des services écosystémiques) faisant partie du métaprogramme INRA EcoServ (services rendus par les écosystèmes), s’inscrit dans cette optique avec comme axe de « comprendre et modéliser le fonctionnement des agroécosystèmes à différentes échelles spatiales et temporelles en lien avec les services fournis ». Ce projet vise à mieux évaluer les fonctions de réponse des espèces face aux conditions environnementales changeantes et de pouvoir intégrer ces informations aux modèles de prédiction existants. En effet, les modèles actuels simulant les phénomènes de compétition entre espèces (tels que GEMINI, Soussana et al., 2012), ne sont pas utilisables à l’échelle des communautés du fait que l’influence des processus biophysiques et biogéochimiques deviennent trop importante à ce niveau.
L’un des principaux objectifs est donc d’améliorer les prédictions des modèles afin de pouvoir en tirer un meilleur parti. Pour cela, il faut comparer les observations faites au cours de différentes études scientifiques et les résultats proposés par les modèles. Il sera ainsi possible de vérifier la précision des prédictions d’une part et d’autre part de calibrer les modèles pour affiner les calculs. Il est donc nécessaire d’identifier les facteurs influençant le développement des prairies de manière précise afin d’être en mesure d’améliorer les prédictions des modèles. Pour cela, il est intéressant de comprendre comment les écosystèmes prairiaux soutiennent les services et comment ils répondent aux changements. Ces informations permettront de mieux comprendre l’impact de la diversité végétale sur les services. Elles pourront également aider à mieux évaluer le maintien de la stabilité des systèmes fourragers en quantifiant plus précisément la résilience aux perturbations à court terme et à plus long terme.
Ce rapport ne traitera néanmoins que la diversité végétale tandis que les autres aspects de la diversité (animale, arthropodes, polinisateurs, faune su sol, micro-organismes etc.) seront exclus.
2
II.
ETAT DE L'ART
A. Définitions
Les prairies : définition et types
Selon l’ouvrage de Gibson (2009), plusieurs définitions sont possibles pour décrire une prairie. La plus simple est ‘un habitat dominé par l’herbe’ cependant elle reste succincte. On peut aussi parler de ‘communauté de plantes constituée d’une couverture basse et sans espèces ligneuses’ (Milner et Hughes, 1968) ou encore ‘un milieu avec une prévalence des poacées, une présence sporadique de ligneux et en général un climat aride’ (Risser, 1988). Il s’agit du biome le plus important, une grande source de biodiversité et les prairies fournissent un ensemble important de biens et de services. Un total de onze types de prairies à utilisations agricoles variées a été défini en 2012 pour l’Union Européenne par 22 experts de 13 pays membres (Peeters et al., 2014). Cette classification est basée sur une étude menée au niveau mondial incluant une trentaine de types (Allen et al., 2011). En Europe les prairies sont gérées de manière à rester au stade herbacé afin de pouvoir être utilisées pour le pâturage. Les types de gestion dépendent de quatre facteurs principaux (Huyghe, 2014) : les caractéristiques géographiques (accessibilité de la parcelle…) ; la production animale associée (type de production : lait, viande, type d’herbivores…) ; l’histoire de la région ; le potentiel de production, défini par le sol et le climat. Les itinéraires culturaux sont donc variables.
Surfaces occupées par les prairies
Selon les statistiques de la FAO (Food and Agriculture Organization; FAOSTAT, 2015), la surface des prairies permanentes représente 67% de la surface agricole utile (environ 3275 millions d’hectares). Cette proportion est de 57% pour l’Amérique du Nord (266 millions d’hectares). Dans l’Union Européenne, cette surface est estimée à environ 57 millions d’hectares soit environ 13% de la surface totale et 33% de la surface agricole utile (Eurostat, 2010 ; Huyghe, 2014), mais pouvant atteindre 65 millions d’hectares soit 35% de la surface agricole utile (FAOSTAT, 2015). En France, la surface occupée par les prairies permanentes est d’environ 9 millions d’hectares soit 32% de la surface agricole utile (FAOSTAT, 2015). Cependant les surfaces prairiales ont largement baissé depuis le siècle dernier dû à la fois à une intensification dans l’utilisation des sols (conversion en cultures) et à une déprise agricole conduisant à l’abandon et l’enfrichement des prairies. Entre 2000 et 2015, on estime leur diminution de 4% à l’échelle mondiale, 8% dans l’Union Européenne et 10% en France (FAOSTAT, 2015). Le changement d’utilisation des sols conduisant à une perte de biodiversité pourrait être néfaste aux services et fonctions que les prairies fournissent (Huyghe, 2014). Les services rendus par les prairies
Les biens et services rendus par les écosystèmes ont été introduits dans le Millenium Ecosystem
Assessment (MEA, 2005). Il s’agit de faire reconnaître à la société, que les écosystèmes rendent
de nombreux services aux activités humaines et à leur bien-être (Haines-Young et Potschin, 2010 ; fig. 1). Une définition possible d’un service peut être : « l’ensemble des propriétés des écosystèmes permettant de produire le bien-être de l’homme ou de la nature » (Fisher et al., 2008). La classification internationale CICES considère que les bénéfices sont définis en quatre catégories : l’approvisionnement en ressources, la régulation des systèmes, le maintien des systèmes et le service culturel (MEA, 2005). Cependant, il n’y a pas de consensus sur la définition d’un service écosystémique (Plantureux et al., 2016) notamment sur la question d’attribuer ou non de la valeur monétaire sur certains services et de considérer qu’il y a les services d’une part et les biens retirés (pour le bien être humain) de l’autre (Bommarco et al., 2013).
3
La fonction agricole principale des prairies est la fourniture de fourrage pour les herbivores domestiques. En plus de produire de la biomasse végétale pour les animaux, les prairies rendent de nombreux services : elles font aussi office de réservoirs de biodiversité.
Figure 1 : Relations entre la biodiversité, les fonctions de l’écosystème et le bien-être humain (Haines-Young etet Potschin, 2010)
Elles ont aussi été reconnues comme pouvant agir comme puits de carbone et il a été montré que les sols des prairies sont plus riches en carbone que les sols cultivés (Amiaud et Carrère, 2012). De ce fait il est très important de conserver ces sols pour limiter les pertes de carbone stocké qui peuvent se produire lors du labour, ou d’événements de canicule par exemple. Les prairies permettent également de protéger les sols en limitant les risques d’érosion du fait de la présence du couvert végétal permanent associé à un réseau racinaire maintenant le sol. L’activité biologique des prairies permet aussi une dégradation plus rapide des amendements organiques et des polluants. La nécessité de conserver ces milieux semble donc évidente pour atténuer les effets du réchauffement climatique, mais aussi pour conserver la diversité des espèces végétales et animales.
Liens entre biodiversité et services écosystémiques des prairies
Les prairies hébergent un grand nombre d’espèces végétales et animales différentes qui ont un rôle plus ou moins important et direct dans la réalisation des services écosystémiques (Amiaud et Carrère, 2012). Chercher à comprendre les liens entre la diversité biologique et le fonctionnement des écosystèmes est donc pertinent. En général, l’impact de la richesse spécifique des communautés végétales sur les services est étudié afin de déterminer les effets de la biodiversité sur les fonctions de l’écosystème (Fridley, 2001 ; Zavaleta et al., 2010). Cette relation entre biodiversité et services n’est que partiellement connue (fig. 1). D’une part, on sait que l’augmentation de la biodiversité est bénéfique pour les services fournis par les écosystèmes (Hervieu, 2002 ; Milne, 2004). D’autre part, les mécanismes liant ces deux notions sont encore méconnus (Hayne-Young et Potschin, 2010 ; Marquard et al., 2009). De plus, d’autres phénomènes comme les stratégies de fonctionnement des plantes (Grime, 1979 ; Tilman 1990) et la plasticité physiologique (Sultan, 2000) font que différentes communautés ayant le même nombre d’espèces n’auront pas forcément la même influence sur les services. De ce fait, le fonctionnement des écosystèmes et leur aptitude à fournir des services dépendent du nombre d’espèces, de leur nature mais aussi des liens qu’elles entretiennent (par exemple la complémentarité) (Maire, 2009).
Diversité taxonomique et fonctionnelle des plantes prairiales
Les prairies permanentes des zones tempérées nord-américaines sont principalement dominées par des espèces de poacées à physiologie C4 (ex : Andropogon gerardii, Sorghastrum nutans,
Bouteloua gracilis, Bouteloua dactyloïdes) (Woodward, 2008). Les prairies permanentes des
zones tempérées européennes sont quant à elles plutôt dominées par des graminées à physiologie C3 (ex : Lolium perenne, Dactylis glomerata).
4
La dominance d’une espèce sur les autres au sein des communautés dépend de ses stratégies de développement mais aussi de ses capacités d’adaptations au milieu dans lequel elle se développe. Pour les plantes prairiales, Grime (1979, 2001) a décrit trois grands types de stratégies et les a positionné sur un triangle appelé CSR : les plantes compétitrices (C), tolérantes au stress (S) et rudérales (R). Un autre auteur a décrit l’adaptation au milieu et les préférences écologiques de plus de 2000 espèces végétales d'Europe Centrale à partir de 6 paramètres environnementaux (lumière, température, continentalité, humidité, acidité du sol, richesse) (Ellenberg, 1988). Ces données sont accessibles notamment via le catalogue des milieux naturels (CATMINAT1) (Julve, 1998).
La biodiversité représente l’hétérogénéité des organismes ou des traits portés par ceux-ci à toutes les échelles (gènes, parcelle, paysage etc.). La diversité biologique pouvant être exprimée par bien des aspects différents, il existe un grand nombre d’indices permettant de la représenter. Il y a donc plusieurs moyens de caractériser la biodiversité d’une prairie. Les principaux indices utilisés pour décrire la diversité alpha sont la richesse spécifique (S), soit le nombre d’espèces ; l'indice de Shannon-Weaver (H’), l’équitabilité (E), la diversité de Simpson (D’) et l’indice de dominance de Berger-Parker (Morris et al., 2014). Dans notre étude, nous avons utilisé l’indice de Shannon qui mesure l’entropie et l’équitabilité variant de 0 à 1 (1 étant le cas où il y a le plus de dominance d’une espèce sur un milieu). Pour la diversité beta (entre deux communautés), les indices les plus couramment utilisés sont ceux de Jaccard et Sorensen. Ces deux indices sont similaires et servent à comparer le niveau de ressemblance de communautés de deux échantillons. Plus la valeur de l’indice est proche de 1 et plus les deux communautés auront un grand nombre d’espèces en commun. Pour obtenir les relevés d’espèces, il existe différentes méthodes d’échantillonnage afin d’évaluer la biodiversité et les communautés d’espèces (quadrats, transects etc.). Il n’existe pas de méthode plus précise que les autres et le choix doit se faire en fonction des objectifs recherchés (Vittoz et al., 2007). En termes du nombre d’espèces, plusieurs typologies ont été proposées pour établir une classification (Scheiner, 2003 ; Dengler, 2009). Une idée assez rependue est celle statuant que la richesse spécifique (S : nombre d’espèces) augmente à mesure que la surface d’échantillonnage augmente (Dengler, 2009). Néanmoins ces résultats ne sont pas toujours évidents. Dans leur publication, Vittoz et al. (2007) n’ont pas trouvé de lien significatif entre la surface d’observation et le nombre d’espèces trouvées. Le concept reste vrai pour les petites surfaces. Les observateurs ont trouvé plus d’espèces dans les zones de 4m² que de 0.4m² cependant ce n’était pas le cas en comparant les résultats des placettes de 4m² et 40m².
La biodiversité peut aussi être définie de manière non taxonomique, via par exemple la détermination des traits des espèces présentes dans la communauté et les fonctions associées. La diversité fonctionnelle fait référence aux différents éléments de la biodiversité qui influence de manière opérationnelle et fonctionnelle les écosystèmes (Violle et al., 2007). Nous n’avons pas travaillé sur cet aspect donc nous ne le développerons pas d’avantage.
B. Réponses de la diversité végétale aux facteurs de gestion
De nombreux facteurs influent sur la diversité des prairies. Ceux-ci sont de natures diverses : biotiques, abiotiques, gestion etc., mais aussi agissent à des échelles diverses (parcelle, paysage). La compréhension et les prédictions de la diversité des prairies permanentes nécessitent donc de prendre en compte de nombreux facteurs et processus (Gaujour et al., 2011). Les effets de l’azote sont assez bien connus et de nombreux travaux ont montré que la fertilisation azotée réduit la richesse spécifique des prairies permanentes (Gough et al., 2000 ; Jacquemyn et al., 2003 ; Rajaniemi, 2002 ; Humbert et al., 2016), quel que soit le type d’N
5
minéral (ammonium, nitrates, urée) (Galka et al., 2005). Les effets du phosphore sur la richesse en espèces sont moins clairs, avec soit des effets plus importants que l’azote (Gaujour et al., 2011), soit n’ayant pas d’effet (Soon et al., 2017). Il existe de nombreux travaux sur les effets de la fertilisation minérale mais étant donné que nous ne les avons pas intégrés à nos recherches, leur influence ne sera pas plus développée.
Fauche
Les dates de fauches sont un élément important pour la composition floristique car couper trop tôt peut limiter la production de graines des plantes non clonales et les coupes tardives peuvent conduire à des parcelles à couverture végétatives denses qui peuvent inhiber l’installation de nouveaux individus (Smith et Haukos, 2002 ; Humbert et al., 2012). La fauche peut également influer sur les transports de graines via les machines pouvant transporter de grandes quantités de graines sur de longue distances de prés en prés (Strykstra et al., 1996 ; Strykstra et al., 1997). Dans le cas des plantes à reproduction sexuée, les coupes avant la floraison empêchent la reproduction. Si la coupe est effectuée juste après la dispersion des graines, une certaine quantité est dispersée par les machines. Dans les deux cas, la richesse spécifique diminue (Smith et al., 2002).
Des fréquences de coupe élevées (trois ou quatre par an) favorisent les espèces avec des feuilles près du sol, celles tirant un avantage de l’augmentation de la lumière au niveau du sol (Carlen et al., 1998). De ce fait, les espèces ayant des feuilles hautes sont désavantagées. La fauche influence les espèces, notamment vis-à-vis de leur capacité à tolérer les coupes et à la vitesse de régénération des feuilles après la coupe (Schipper et Joenje, 2002). La fauche augmente la quantité de lumière disponible dans les couches basses de la végétation, réduisant la compétition pour la lumière et facilitant ainsi la coexistence d’espèces ayant des capacités de développement différentes (Kull et Zobel, 1991). En général, la végétation est plus diversifiée dans les prairies fauchées que pâturées (Gaujour et al., 2011). Principalement parce que les conditions en milieu fauché sont plus favorables pour la dispersion des graines et l’installation de nouvelles espèces. Dans le cas où les banques de graines et la dispersion des plantes ne sont pas des facteurs limitants, la hauteur de la pelouse a un effet important sur la germination et la croissance des plantes (Plantureux et al., 2005). Ainsi, une gestion de coupe plus ou moins haute influera sur la disponibilité de la lumière et donc sur les espèces pouvant se développer ou non, et sera aussi fonction de la position des méristèmes localisés plus ou moins haut sur les tiges où les talles pour les poacées.
Pâturage
Le pâturage affecte la composition floristique des prairies permanentes et dépend du chargement animal (Diaz et al., 2001), des saisons (Sternberg et al., 2000) et du type d’animaux (Huntly, 1991). L’augmentation du chargement animal favorise les plantes rudérales et compétitives (sensu Grime et al., 1988), en particulier les poacées très compétitives (Dumont et al., 2009), la régénération annuelle via les graines, la floraison et la dispersion précoce de graines, la morphologie en rosette, et les espèces héliophiles à hauteur faible (Pakeman, 2004). Des chargements animaux faibles favorisent les espèces herbacées tolérantes au stress (sensu Grime et al., 1988) et les autres espèces diverses (Dumont et al., 2009). L’abondance en dicotylédones croît avec l’augmentation du chargement en ovins (Bullock et al., 2001). Deux mécanismes sont mis en avant pour expliquer les effets du pâturage : le prélèvement de la végétation et le piétinement. Le pâturage conduit à une hétérogénéité spatiale du couvert végétal (Wallace 1987) par défoliation sélective et cela ouvre des niches de régénération (sensu Grime 1979). Le fait d’enlever la végétation permet à la lumière d’atteindre les couches de végétation les plus basses jusqu’à la surface du sol, ce qui réduit la compétition pour les ressources (les plantes rampantes comme Trifolium repens se développent très bien dans ces espaces ouverts)
6
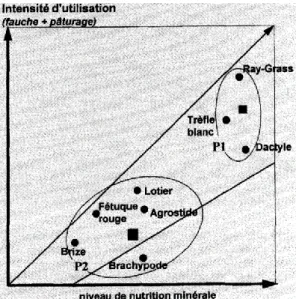
(Pavlu et al., 2007 ; Pavlu et al., 2006). D'après la figure 2, on observe que selon les intensités de fauche, de pâturage et de fertilisation, les communautés sont très différentes. On remarque par exemple que le trèfle blanc peut mieux se développer dans les prairies utilisées intensivement, du fait de canopée plus basse. Le ray-grass et le dactyle sont des poacées assez nitrophiles donc compétitives dans les milieux amendés en nitrates.
Le motif peut être régulier (bovins, ovins) ou différencié (chevaux) (Gaujour et al., 2011). Les herbivores pâturent de préférence les espèces de haute qualité fourragère préalablement broutées dans de petites zones (Adler et al., 2001). Dans le cas de mélanges d’animaux, les effets peuvent être additionnés ou compensés selon si les habitudes alimentaires sont similaires ou non (Loucougaray et al., 2004).
Un effet positif sur la richesse en espèces est globalement observé en faveur du pâturage plutôt que de la fauche. Les effets du pâturage/fauche sont différents selon les continents, le type de prairies et l’altitude. La méta-analyse (Tälle et al., 2016) a montré un effet positif du pâturage sur l’abondance, la biomasse. Sur le court terme à la différence de la fauche qui aurait un effet bénéfique sur le long terme.
Figure 2 : Dominance moyenne de certaines espèces de prairies en fonction du niveau de l’intensité d’utilisation et du niveau de nutrition minérale (Duru et al., 1998).
C. Réponses de la diversité végétale aux facteurs climatiques
Il existe potentiellement quatre raisons pour lesquelles une communauté peut être plus résistante au changement climatique : antécédents d’exposition à des phénomènes climatiques extrêmes, statut de succession, état de la diversité et composition fonctionnelle (Carlyle et al., 2014).
Elévation des températures
D'après deux méta-analyses récentes, il a été mis en évidence que le réchauffement de l'air a des effets positifs, nuls ou négatifs sur la production de biomasse (Wang et al., 2012 ; Dieleman et al, 2012). Cantarel et al., 2012 ont aussi montré que le réchauffement avait un effet négatif sur la réserve des plantes et donc sur la production de biomasse. En effet, si dans un premier temps la photosynthèse peut être stimulée par le réchauffement, l’évapotranspiration augmente (du fait du réchauffement de l’air) conduisant à une diminution de l’eau du sol et donc à une baisse de productivité (Cantarel et al., 2012 ; Dai, 2011). Il est important de noter également que la réponse au réchauffement varie selon les types de métabolismes C3 et C4 ou encore les groupes fonctionnels des plantes (Wang et al., 2011).
Le réchauffement augmente la dominance liée à la baisse de diversité : changement de type de plantes. Par exemple, dans le cas des toundras, les plantes vasculaires vont remplacer les mousses et les lichens (Cornelissen et al., 2001 ; Walker et al., 2006 ; Lang et al., 2012). L’article d’Elmendorf et al. (2012) montre également que les effets de la température varient selon la température estivale. En effet, pour les températures les plus basses (environ 2°C), l’augmentation des températures augmente le nombre d’espèces.
Beaucoup d’articles ont montré que les remplacements induits par la compétition jouaient un rôle important dans la baisse de la richesse spécifique avec une accélération de la compétition
7
exclusive due au réchauffement (Klanderud et Totland, 2005 ; Limberger et al., 2014). La compensation des pertes en espèces locales pourrait se faire en dispersant sur de grandes distances des plantes adaptées, plus tolérantes, aux zones plus chaudes. La méta-analyse de Gruner et al. (2017) tend à montrer que globalement, la richesse spécifique locale (tout organisme confondu) baisse avec l’augmentation des températures. Cette baisse du nombre d’espèces peut être associée à une augmentation de la biomasse plus élevée (s’il n’y a pas de stress hydrique) entrainant plus de compétition pour la ressource. Les facteurs de réchauffement et d’augmentation de CO2 atmosphériques sont liés et cette relation n’est pas simplement additive mais plus complexe (Dieleman et al., 2012).
Modification des précipitations
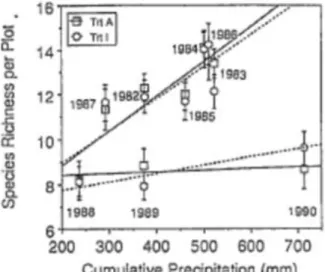
La disponibilité en eau dans le sol est un facteur impactant largement la production de biomasse et la sélection des espèces. En effet, le stress hydrique joue un rôle sur la composition du cortège floristique (Grime et al., 2008) en favorisant les espèces à stratégie de conservation (Picon-Cochard et al., 2013). Les effets sur la richesse spécifique varient selon les fréquences et intensité des épisodes de sécheresse. Une sécheresse ponctuelle forte peut induire de fortes mortalités (exemple de 20% de perte d’espèces : Tilman et Haddi, 1992; fig. 3). De même, d’après la figure 3, on observe que le nombre d’espèces est corrélé avec le taux de précipitation cumulées et que le nombre d’espèces augmente avec la disponibilité en eau pour la
Figure 3 : Richesse spécifique de chaque parcelle en fonction des précipitations cumulées entre 1982 et 1990 (Tilman et Haddi, 1992).
période 1967-1986. Cependant, une telle corrélation n'est pas constatée entre 1988 et 1990. Ce résultat est dû au fait que la disponibilité en eau des sols est moindre pour la deuxième période du fait d'une température ambiante plus importante.
Paysages
Les échelles spatiales sont importantes à prendre en compte dans la réponse de la diversité végétale car la structure des paysages influence d’abord les flux de graines donc par la suite la végétation des prairies. On distingue trois éléments : l’hétérogénéité, les éléments et la fragmentation. Cependant les effets sont moins clairs en ce qui concerne les prairies. La non corrélation entre l’hétérogénéité et la richesse en espèces pourrait être due aux pratiques agricoles qui modifient la colonisation et/ou les matrices de plantes (Dauber et al., 2003 ; Krauss et al., 2004).
8
D. La méta-analyse
Méthodologie statistique pour quantifier l’effet de facteurs environnementaux sur la diversité végétale en prairie : approche par méta-analyse
La méta-analyse a été développée en 1976 par Gene V. Glass en 1976 (Université du Colorado). Il s’agit de l’analyse statistique des résultats d’un nombre donné d’études scientifiques et connote d’une certaine rigueur méthodologique par rapport aux synthèses bibliographiques (i.e. reviews) (Glass, 1976). Les méta-analyses ont d’abord été utilisées en sciences médicales et sociales puis dans d’autres domaines scientifiques (biologie, écologie etc.) par la suite (Nakagawa et al., 2017).
Pour une majorité de méta-analyses, la phase de recherche des articles est systématique et bien documentée afin de pouvoir être reproductible.
Figure 4 : Nombre de méta-analyses publiées par an pour le domaine de l’écologie végétale (Koricheva et Gurevitch, 2014).
Il existe le protocole PRISMA (Preferred Reporting Items for Systematic Reviews and Meta-Analyses) permettant de systématiser les recherches de données (Nakagawa et al., 2017). Ces éléments à considérer (items) sont au nombre de 27 afin de rapporter au mieux les informations récoltées (exemples : objectifs de la recherche, méthode de sélection des données, risque de biais) (Liberati et al., 2009). En terme de nombre d’articles analysés, une méta-analyse avec moins de 20 publications sera plutôt considérée comme petite et plutôt grande au-delà de 50 articles.
Depuis vingt ans les méta-analyses sont de plus en plus utilisées. Dans le domaine de l’écologie, 322 ont été identifiées entre 1996 et 2013 dans 95 journaux différents (fig. 4). La généralisation de cette méthode statistique a conduit à la création de toute une littérature dédiée à la bonne utilisation des méta-analyses, notamment en écologie. A titre d’exemple, nous pouvons citer l’ouvrage de Koricheva, Gurevitch et Mengersen (2013).
E. Objectifs
Le but du stage est d’étudier l’influence des facteurs environnementaux et de gestion influençant la diversité végétale des prairies. L’approche via une méta-analyse permet de synthétiser les informations disponibles dans la littérature via une quantification des effets, ainsi que de regrouper les résultats observés dans les différentes expériences menées. Par la suite, les résultats obtenus pourront être utilisés pour être comparés avec les prédictions de modèles de simulation développés dans l’unité UREP et ainsi améliorer les calibrations des modèles. Pour cela il est important de définir les problématiques principales à savoir : i. influence de la fauche et du réchauffement sur la diversité des prairies préalablement abandonnées. ii. Est-ce que la hauteur, les surfaces et les durées d’expérimentation jouent un rôle sur la diversité de ces contextes ?
Sélection des facteurs
Le premier objectif a été d’abord de déterminer les facteurs dans la littérature via des expériences, des synthèses et des méta-analyses. Le deuxième objectif a été de déterminer quels facteurs nous allions étudier. En effet, tous les intégrer à notre méta-analyse n’était pas possible car cela nécessiterait trop de temps par rapport à la durée du stage. Les facteurs ont été choisis
9
selon trois contraintes. En premier lieu, le facteur considéré se devait d’avoir été suffisamment étudié afin que des données soient disponibles pour pouvoir être utilisées dans notre méta-analyse. La seconde contrainte était que le facteur devait aussi ne pas avoir été préalablement étudié de manière globale. Notre but n’étant pas de refaire des travaux déjà faits. C’est pour cela que nous n’avons pas considéré l’azote car beaucoup d’études avaient déjà été publiées, notamment la méta-analyse de Humbert et al. (2016). Le fait que le facteur choisi ai un effet clair et avéré ou non n’était pas un élément de décision car le but même d’une méta-analyse est de pouvoir étudier différentes études afin de mieux évaluer les résultats ayant été publiés et ensuite donner des pistes sur les points de recherche potentiellement prometteurs.
Nous avons choisi le nombre de fauches (zéro ou une par an) en relation avec les hauteurs de coupes et les surfaces des parcelles, car les données étaient disponibles dans une base de données. Nous avons aussi choisi le réchauffement climatique car ce facteur est largement étudié et les effets de l’augmentation des températures varient selon les milieux. De plus, peu de méta-analyses traitent du sujet. Nous comptions aussi étudier les variations de pluviométrie mais nous n’avons pas eu le temps de l'étudier dans la période impartie par le stage.
Sélection des indicateurs de biodiversité
Comme il a déjà été présenté dans l’état de l’art, il existe de nombreux indicateurs de biodiversité : taxonomiques (nombre d’espèces) ou non taxonomiques (traits). Le choix s’est basé sur la disponibilité des données. Nous avons sélectionné les indicateurs les plus utilisés : richesse spécifique (S), indice de Shannon-Weaver (H’) et d’équitabilité (E). Nous avons également considéré la biomasse car cette donnée est très disponible du fait que la production primaire est un service très important rendu par les prairies.
III. MATERIELS et METHODES
Le travail s’est partagé en deux parties principales : la première a consisté à reprendre et compléter un jeu de données puis d’étudier ses valeurs (impact de la fauche sur la diversité végétale); la seconde partie a été de faire un bilan des principaux facteurs influençant cette diversité des prairies, d’en sélectionner certains, de rechercher des articles, puis d’encoder les données pour les analyser. Dans ces deux parties, une méta-analyse a été utilisée pour quantifier des effets de changement de gestion et du changement climatique sur la diversité végétale de prairies issues de différents pédo-climats. Sur l’ensemble des facteurs impactant la diversité floristique des prairies ayant été étudiés et présentés dans l’état de l’art, deux ont été retenus : les changements de la fréquence de fauche et de changement de températures liés au réchauffement climatique.
A. Documentation
a. Recherche des sources bibliographiques
Pour les deux méta-analyses, la recherche d’articles a été faite à partir de la base de données Web of Science entre mai 2014 et août 2016 (fauche) et en juin 2018 (température et précipitation). Les mots clés utilisés pour le facteur fauche étaient: “(cutting OR mowing OR clipping) AND (plant diversity OR evenness OR richness) AND (grass* OR prairie) ” ont permis d’obtenir 659 articles. Pour le facteur température, quatre ensembles de mots clefs ont été utilisés. Pour les types de milieux: “grassland OR meadow OR pasture OR pampa OR steppe OR prairie OR savanna OR tundra”. Pour la diversité : “(diversity OR richness OR evenness OR cover OR disturbance) AND plant* OR functional type”. Pour le réchauffement : “(temperature OR warm) AND air OR heat AND stress OR extreme temperature”. Ces
10
ensembles ont ensuite été combinés (via l’opérateur booléen AND) pour effectuer deux recherches. Pour cette recherche nous avons obtenu respectivement 424 résultats. Les listes ont été exportées sous forme de tableur (Excel) pour pouvoir trier les articles pertinents ou non. Aux articles traitant du réchauffement de l’air, 26 publications, issues de la base de données de l’article de Gruner et al. (2017) ont été ajoutées.
b. Sélection des articles pertinents
La sélection des articles s’est faite de la même façon pour les deux facteurs (réchauffement et fauche) et selon la méthode PRISMA (Liberati et al., 2009). Il s'agit d'un protocole permettant de sélectionner les articles selon différentes étapes et ensuite de pouvoir communiquer sur les choix de sélections sous forme d'un schéma. La figure pour la méta-analyse sur l'effet de la fréquence de fauche est présentée dans les résultats (fig. 5) et celle pour l'effet du réchauffement en annexe 1.
c. Création des bases de données
Pour mettre en forme le tableau de données, nous nous sommes basés sur deux tableaux préexistants, le premier défini par C. PICON-COCHARD pour la méta-analyse traitant de l’effet de la fauche et le second, publié dans l’article Gruner et al. (2017).
B. Analyses statistiques
a. Extraction des données
La base de données était déjà existante, la sélection des articles déjà faite ainsi que l’extraction d’un certain nombre de données (moyennes, tailles des échantillons etc.). Cependant, les écart-types et erreur standard n’avaient pas été reportés. L’un des premiers travaux a donc été d’intégrer ces données au tableau afin de pouvoir les traiter par la suite. Ces valeurs ont pu être récupérées pour 35 publications. Pour chaque article analysé, en cas de données multiples (plusieurs dates, hauteurs de coupes, traitements etc.) pour les indices biologiques ou la biomasse, le maximum de données a été renseigné. Cependant, une seule valeur moyenne du ratio a été calculée par article. C’est pour cela que nous cherchions en priorité les études pondérées afin de pouvoir mieux apprécier la variabilité intra-étude afin de la comparer avec la variation totale des modèles (inter-études). L’utilisation des modèles mixtes a permis aussi de mieux évaluer la variété de traitements parmi les études.
Les méthodes pour obtenir les moyennes, écarts-types et erreurs standards dépendaient de la nature des données publiées. Les cas les plus simples étant lorsque les données étaient présentées sous forme de tableau ou de texte. Dans ce cas, elles ont été recopiées dans les colonnes correspondantes. Lorsque les données étaient exposées sous forme de graphique, le logiciel WinDIG (développé par D. Lovy, Université de Genève2) a été utilisé afin de les récupérer (illustration 1). Ce logiciel permet d’extraire les moyennes et les écart-types. Il s'agit du logiciel permettant une lecture graphique plus précise.
11
Illustration 1 : Capture d’écran du logiciel WinDIG. La croix rouge représente la localisation du pointeur de la souris. Sa position est montrée de manière plus précise dans la fenêtre ‘WinDIG Zoom’. Le nombre encadré en rouge représente la valeur en ordonnée (richesse spécifique) du point cliqué selon l’échelle, soit 21.133. L’échelle est définie par l’utilisateur selon trois points. Les valeurs d’erreur standard ont été calculées en fonction des longueurs de barres ou, dans le cas des boxplots avec des valeurs de “range”(Weir et al., 2018).
Dans le cas de l’étude des effets du réchauffement les données de 29 publications ont pu être extraites. Les valeurs de moyennes, range, erreur standard ont été récupérées de la même façon que pour la première méta-analyse.
b. Traitement des données
La plupart des méta-analyses sont basées sur trois types de modèles statistiques de bases. Mais le raisonnement principal est toujours le même, déterminer si on observe un effet ou non. Sous forme mathématique, cela peut être exprimé par l’équation: yi = θi + ei. Avec : yi : effet observé
dans i études; θi : vrai effet (inconnu); ei : erreur d’échantillonnage (connu).
Les trois types de modèles sont les modèles à effet aléatoire, les modèles à effet mixte et les modèles à effet fixé. Ce dernier type de modèle n’ayant pas été utilisé, il ne sera pas détaillé ici. Taille d’effet (effect size)
Dans tous les cas, le but des méta-analyses est de déterminer la proportion de la taille d’effet. La taille d’effet (effect size) est un calcul de l’influence d’un facteur sur un autre pour une variable donnée. En méta-analyse, ces effets sont caractérisés principalement selon trois types : comparaison de deux moyennes (ex : response ratio, Hedges et al., 1999), analyse de corrélation (ex : r/r²) et comparaison deux à deux (ex : relative risk) (Nakagawa et Santos, 2012).
Ratio de réponse (response ratio)
Le ratio de réponse est fréquemment calculé dans les méta-analyses afin d’avoir une mesure des effets expérimentaux. Il s’agit du rapport de valeurs quantitatives mesurées dans le traitement utilisé sur le traitement témoin. Ce ratio permet de quantifier la proportion de changements due aux manipulations de l’expérience. Si on note la moyenne des valeurs contrôles Xc et les valeurs moyennes expérimentales Xe, ce ratio s’écrit : RR = Xe/Xc. Il est
préférable d’analyser ces valeurs de ratio via un logarithme népérien (ln(RR)). Cela peut être expliqué par deux raisons. La première est que le logarithme népérien linéarise les données en attribuant le même poids au numérateur et au dénominateur du ratio de réponse. Ainsi, le numérateur et le dénominateur auront la même influence sur la déviation des résultats. La seconde raison est que les valeurs de ln(RR) sont réparties de manière plus normale que les valeurs de RR pour les petits échantillons (Hedges, 1999).
𝐿𝑅𝑅 = 𝐿𝑛 ( 𝑚1𝑖 𝑛1𝑖 ⁄ 𝑚2𝑖 𝑛2𝑖 ⁄ )
avec m1i: moyenne du nombre d’espèces en condition une fauche par publication ; m2i : moyenne du nombre d’espèces en condition d’abandon par publication ; n1i : nombre de répétitions en condition une fauche par publication ; n2i : nombre de répétitions en condition d’abandon par publication ;
Modèle à effet aléatoire (random effect model)
En général les méta-analyses se basent sur des publications qui ne sont pas parfaitement identiques dans leurs méthodes d’échantillonnages. Ces différences de méthodes peuvent introduire de la variabilité au sein de l’effet réel. Ce type de modèle n’impose aucune contrainte
12
mathématique (i.e. aucun co-facteur est fixée) et donc on observe une variation libre de la réponse. On détermine alors un effet réel moyen (µ) et la part totale d’hétérogénéité (τ²) (Wolfgang, 2010).
Modèle à effet mixte (mixed-effect model)
Cette méthode dérive de la méthode à effet aléatoire et consiste à introduire un ou plusieurs modérateurs (facteurs d’études) afin d’expliquer une partie de l’hétérogénéité entre les études. Dans ce cas, τ² n’est plus l’hétérogénéité totale mais la partie résiduelle n’étant pas expliquée par les modérateurs (Wolfgang, 2010). L’intérêt principal de ce type de modèle est de tester des modérateurs de manière indépendante ou combinée avec d’autres co-facteurs afin de déterminer s’ils ont un effet ou non sur les variations de l’échantillon et dans quelle proportion. Trois valeurs sont importantes à relever à la sortie du logiciel R : le R² : pourcentage de l’hétérogénéité expliqué par le modérateur ; p-val QM : valeur p du test des modérateurs, s’il est significatif à un seuil défini alors le modérateur peut être jugé comme ayant un impact sur l’effet; p-val QE : valeur p du test de l’hétérogénéité des résidus, s’il est significatif, on peut considérer que d’autre modérateurs influent sur l’effet du modèle.
Biais
Il s’agit d’un point important de l’étude statistique. Il y a un risque de biais lié aux publications quand la probabilité de publication dépend de la signification statistique, la magnitude ou la direction de l’effet. La capacité à détecter ces biais est essentielle pour s’assurer de la force statistique d’une méta-analyse (Nakagawa et Santos, 2012). Il existe plusieurs méthodes pour évaluer ces biais : graphique (ex : funnel plot) ou via des tests statistiques (fail-safe number (Jennions et Moller, 2002). Cependant, ces méthodes peuvent se révéler insuffisantes et certaines (comme les funnels plots) ont été modifiées pour mieux correspondre à l’identification des biais de publication (Peters et al., 2008).
Les données ont été traitées avec le logiciel R (version 3.3.3 x64) en utilisant le package metafor (Viechtbauer, 2010), qui fournit un ensemble de fonctions dédié aux analyses statistiques à mettre en œuvre dans les méta-analyses. Le package boot a également été utilisé pour calculer le bootstrap pour l'intervalle de confiance pour les articles dont on ne disposait pas des écart-types (Canty et Ripley, 2017). L’étude des données s’est faite en suivant les exemples donnés dans la publication de Viechtbauer (2010). L’une des premières tâches a donc été d’adapter nos données aux exemples présentés.
Quatre modalités ont été testées pour affiner l’effet de la fauche sur la richesse spécifique : les hauteurs de coupes (séparées en deux classes), la surface d’observation (trois classes), la durée d’expérimentation et la date de publication des articles (test des biais). Nous n’avons considéré que les cas d’abandons (absence de fauche) ou d’une seule fauche par an car il n’y avait pas assez de données pour les autres fréquences de fauche. Quatre modalités ont été testées pour définir les effets du réchauffement sur le nombre d’espèce, l’indice de Shannon-Weaver et l’équitabilité : la différence de température entre le contrôle et le test, la durée d’expérimentation, la surface d’échantillonnage et l’année de publication des articles (test des biais).
Les biais de publication ont été testés via le traitement de l’année de publication comme une modalité dans les modèles à effet mixte et par l’utilisation de graphiques en entonnoir (funnel
13
IV.
RESULTATS
A. Méta-analyse sur les effets de la fauche sur la diversité végétale des prairies
a. Schéma Prisma
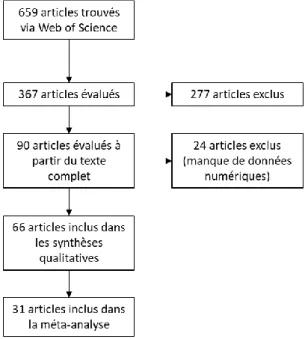
L’une des problématiques majeures pour l’analyse systématique est d’une part de trouver des données permettant d’apporter des informations sur le problème scientifique posé et d’autre part d’être en mesure d’utiliser ces données dans la méta-analyse. Le schéma Prisma (fig. 5) montre ainsi que de tous les articles repérés via la base de donnée Web of Science, seulement 31 (4.7%) ont été utilisé dans la méta-analyse (effet d’une coupe par an sur le nombre d’espèces par rapport à l’absence de coupe). Ce nombre baisse en considérant d’autre paramètres : biomasse, 18 articles ; hauteur de coupe, 19 articles. Enfin, l’influence de la surface d’échantillonnage a pu être étudiée sur 31 articles. L’une des causes principales de l’impossibilité d’utiliser certains articles a été le manque de valeurs de SE ou de SD. Ces valeurs étant essentielles afin de mieux pouvoir apprécier l’effet taille (hétérogénéité) dans les bases de données étudiées. La liste des articles est disponible en annexe 4a.
Figure 5 : Schéma Prisma (Liberati, 2009) : informations retenues au cours des différentes étapes de sélection des articles pour la méta-analyse sur l’effet de la coupe sur la biodiversité.
b. Résultats pondérés et non pondérés
La méta-analyse a été réalisée sur 31 articles qui présentaient des valeurs de moyennes de richesse spécifique et d’écart-type. 35 autres publications donnaient uniquement des informations de moyenne du nombre d’espèces. Ces publications ne donnant pas de valeurs de variabilité intra-étude, elles n’ont pas pu être intégrées dans la méta-analyse. Cependant, ce total de 51 publications a permis de réaliser un calcul de moyennes et d’intervalle de confiance via une technique d’inférence statistique : le bootstrap (fig. 6). Après calculs, on observe que les résultats entre les calculs à partir de moyennes pondérées (modèle à effet aléatoire) et non pondérées (bootstrap) sont relativement proches (0.01 points de différence). De plus, les intervalles de confiance se recoupent et sont du même ordre de grandeur bien que celui obtenu par inférence statistique soit plus petit (0.08 contre 0.1 pour le modèle à effet aléatoire). L’inférence statistique permet donc de montrer que l’échantillon utilisé dans la méta-analyse de 31 articles parmi les 66 initiaux est représentatif de l’ensemble des publications sélectionnées au départ.
Figure 6 : Ratio de réponse du nombre d’espèces prairiales à une coupe par rapport à l’absence de coupe calculée avec un jeu de données de 31 articles (orange : modèle à effet aléatoire) et avec un jeu de données de 51 articles (gris : bootstrap). La taille de l’effet est évaluée selon le logarithme népérien du ratio de réponse, selon deux méthodes avec les écart-types et sans, via une inférence statistique : bootstrap. Les barres verticales représentent les intervalles de confiance à 95%.
14
c. Modèles à effet aléatoire et à effet mixte
Effets sur le nombre d’espèces
Comme expliqué dans les parties précédentes, 31 articles ont permis le calcul d’un modèle à effet aléatoire. Le but étant : i. d’évaluer si le nombre d’espèces était différent entre les prairies fauchées une fois par an et celles non fauchées et ii. de quantifier cette différence si elle existe (fig. 7a).
Le funnel plot (annexe 2a), montre que la majorité des données sont dans le même ordre de grandeur. Cinq sont plus éloignées et une se démarque franchement avec ratio de réponse égale à environ 0.5 et une erreur standard de 0.5 environ. On retrouve cette publication dans le graphique en forêt (fig. 7a) et il s’agit de l’article de Truus et al. (2009). De manière générale, le funnel plot montre que nos valeurs sont assez variables, cependant il ne semble pas y avoir de biais de publication (par exemple, il n’y a qu’un seul nuage de points).
Effets sur la quantité de biomasse
Le modèle à effet aléatoire a été calculé à partir de données de 18 articles afin d’évaluer si la quantité de biomasse produite variait avec le nombre de fauches par an (0 ou 1) ainsi que de quantifier cet effet (fig. 7b).
Les indices statistiques et leurs valeurs chiffrées pour les deux modèles à effet aléatoire sont présentés dans le tableau 1a. Les résultats et valeurs d’indices statistiques obtenus pour les calculs de modèles à effet mixte (à partir de 16 articles) sont présentés dans le tableau 1b.
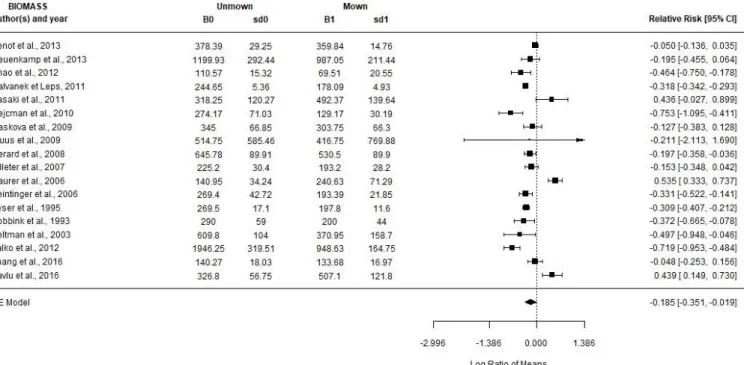
Figure 7a: Graphique en forêt (forest plot) montrant pour chaque étude des moyennes de logarithmes népériens du ratio de réponse (SR1/SR0) ainsi que les intervalles de confiance à 95% par étude et pour le modèle (RE model). La taille d'un point représente son poids dans l'étude. Species richness : richesse spécifique ; Authors(s) and year : auteur(s) et année de publication de l’article ; unmown : modalité aucune fauche par an; mown : modalité une fauche par an ; SR0 : moyenne de richesse spécifique pour la modalité aucune fauche ; SR1 : moyenne de richesse spécifique pour la modalité une fauche par an ; sd0 et sd1 : écart-types associés aux valeurs de SR0 et SR1 ; relative risk [95% CI] : valeur de ln(SR1/SR0) et valeurs de l’intervalle de confiance à 95%. ; RE Model : random effect model : moyenne et intervalle de confiance pour l’ensemble des articles.
15
Figure 7b : Graphique en forêt des moyennes de logarithmes népériens du ratio de réponse (B1/B0) ainsi que les intervalles de confiance à 95% par étude et pour le modèle (RE model). La taille d'un point représente son poids dans l'étude. Biomass : biomasse ; Authors(s) and year : auteur(s) et année de publication de l’article ; unmown : modalité aucune fauche par an; mown : modalité une fauche par an ; B0 : moyenne de biomasse (g.m-2) pour la modalité aucune fauche ; B1 : moyenne de biomasse (g m-2) pour la modalité une fauche par an ; sd0 et sd1 : écart-types associés aux valeurs de B0 et B1 ; relative risk [95% CI] : valeur de ln(SR1/SR0) et valeurs de l’intervalle de confiance à 95%. ; RE Model : random effect model : moyenne et intervalle de confiance pour l’ensemble des articles.
Modèle Moyenne LRR IC pval t² I² (%) H²
SR 0v1 0.2257 0.0927 <0.0001 0.0555 89.11 9.18
B 0v1 -0.1852 0.166 0.0288 0.1064 95.3 21.28
Tableau 1a : Valeurs d’indices statistique pour les modèles à effet aléatoire pour la richesse spécifique (SR 0v1) et pour la production de biomasse (B 0v1). Moyenne LRR : moyenne des valeurs de logarithme népérien de ratio de réponse : IC : intervalle de confiance ; pval : valeur de p ; t² : quantité d’hétérogénéité ; I² : pourcentage de variabilité dans la taille d’effet calculé via l’échantillon, pouvant être attribué à l’hétérogénéité de l’effet réel ; H² : ratio de la variabilité totale sur la variabilité de l’échantillon.
Le funnel plot pour le modèle aléatoire de la biomasse (annexe 2b) est assez semblable à celui du modèle aléatoire pour la richesse spécifique. Les données sont groupées mais avec une variabilité importante et une publication se démarque franchement (Truus et al., 2009).
Les analyses statistiques montrent qu’une fauche annuelle par rapport à une prairie non fauchée a un effet positif (+25%, exp(LRR) = 1.25) sur la richesse en espèces et un effet négatif (-17%, exp(LRR) = -0.185) sur la biomasse. Dans les deux cas, pval est inférieure à 0.05, indiquant donc des résultats significatifs. Ces résultats sont corroborés par les valeurs d’I² supérieures ou égales à 90% qui impliquent que 90% ou 95% de la variabilité observée peut être attribuée à la variabilité réelle de l’effet. Les valeurs de H² sont supérieures à 1 donc les effets observés ne sont pas uniquement dû à la variabilité des échantillons.
Dans le cas du graphique en forêt pour le cas de la richesse spécifique (fig. 7a), on observe qu’une seule étude montre un effet opposé aux autres : Huhta et al. (1998). On distingue également que trois études dénotent des effets deux fois supérieurs à la moyenne du modèle : Metsoja et al. (2013), Neuenkamp et al. (2013) et Fenner et Palmer (1998).
Dans le cas de la biomasse (fig. 7b), on distingue qu’une publication a une très forte variabilité : Truus et al. (2009). Trois publications ont un effet positif, donc opposé à la tendance générale :
16
Sasaki et al. (2011), Mauer et al. (2006), Pavlu et al. (2016). Enfin, deux publications ont des effets beaucoup plus négatifs que la moyenne : Hejcman et al. (2010) et Valko et al. (2012). Nous avons écrit dans l’état de l’art que la hauteur de fauche et la taille de la zone d’observation peuvent avoir un effet sur la diversité, c’est pourquoi nous avons réalisé un modèle mixte permettant de tester l’effet de variable influente sur la diversité. Suite aux analyses données par le modèle mixte, on observe que la modalité durée d’expérimentation (D) a un effet significatif sur la richesse spécifique (p-val QM < 0.05). Il est donc possible de rejeter H0 : SR0 = SR1 (tab. 1b). Pour les modalités Height et Plot, les valeurs de p-val QM sont supérieures à 5% (respectivement 0.6 et 0.7). Il est donc impossible d’affirmer que la hauteur de coupe et la surface d’observation jouent un rôle dans la variabilité de la richesse spécifique et de la biomasse (i.e. H0 ne peut pas être rejetée). A noter aussi que pour toutes les modalités, les valeurs de p-val QE sont inférieures à 5% donc il est sûr que d’autres variables que nous n'avons pas introduites dans nos modèles pourraient jouer un rôle dans la différence du nombre d’espèces.
Afin de tester les biais de publication, nous avons testé l’année de publication comme modalité. Via les modèles mixtes, il sort que cette co-variable n’a pas d’influence sur le nombre d’espèces (Y : p-val QM = 0.4214 ; tab. 1b). Par contre il y a un effet de cette modalité quand elle est combinée à la durée d’expérimentation (DxY : p-val QM = 0.0004). Il est donc important de prendre en compte le fait que l’année de publication joue un rôle sur la durée d’expérimentation (dans le logiciel R, on obtient une moyenne de LRR négative (-0.0143), donc on peut en déduire que la durée des études baisse avec les années. i.e. les durées d’expérimentation publiées dans les articles récents sont plus courtes que par le passé).
Modalité t² p-val QE p-val QM R² (%)
Height 0.012 0.0002 0.5844 0 Plot 0.123 <0.0001 0.704 0 HxP 0.132 <0.0001 0.8137 0 D 0.0244 <0.0001 0.0019 21.43 Y 0.0313 <0.0001 0.4214 0 DxY 0.0198 <0.0001 0.0004 36.46
Tableau 1b : Valeurs d’indice statistique pour les modèles à effet mixte calculés pour la richesse spécifique. Les modalités hauteur de coupe (Height), surface d’échantillonnage (Plot) ; combinaison des deux (HxP) ; D: durée d’expérimentation ; Y : date de publication ; DxY. t² : quantité estimée d’hétérogénéité résiduelle ; p-val QE : valeur p du test sur l’hétérogénéité résiduelle ; p-value QM : test sur les modérateurs ; R² : pourcentage de variance expliqué par la modalité.
B. Méta-analyse effets du réchauffement sur la diversité végétale des prairies
a. Schéma Prisma
Le schéma est disponible en annexe 1. Les articles pour la méta-analyse ont été récupérés à partir de deux bases de données : Web of Science (424 articles) et l’article de Gruner et al. (2017) (26 articles). L’article traitant de différents milieux, nous n’avons conservé que les données sur les systèmes herbacés. Au final, 37 articles ont pu être utilisés pour les synthèses qualitatives et les données de 29 publications ont été extraites pour les calculs de modèles. La liste des articles est disponible en annexe 4b.
17
b. Modèles à effets aléatoire et à effet mixte
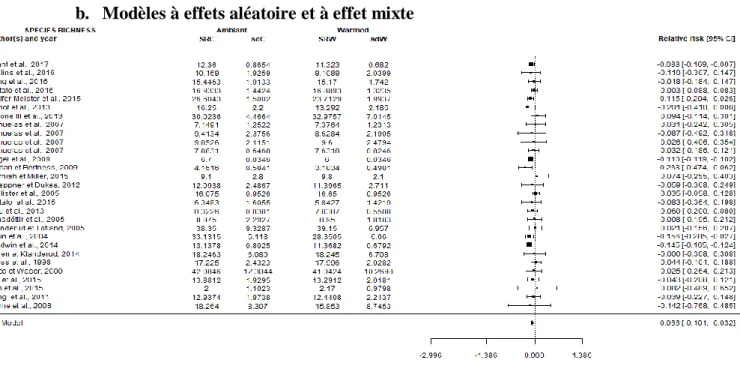
Figure 8 : Graphique en forêt (forest plot) des moyennes de logarithmes népériens du ratio de réponse (SRW/SRC) ainsi que les intervalles de confiance à 95% par étude et pour le modèle (RE model). La taille d'un point représente son poids dans l'étude. Species richness : richesse spécifique ; Authors(s) and year : auteur(s) et année de publication de l’article ; ambiant : modalité température ambiante ; warmed : modalité réchauffement ; SRC : moyenne de richesse spécifique pour la modalité température ambiante ; SRW : moyenne de richesse spécifique pour la modalité réchauffement ; sdC et sdW : écart-types associés aux valeurs de SRC et SRW ; relative risk [95% CI] : valeur de ln(SRW/SRC) et valeurs de l’intervalle de confiance à 95%. ; RE Model : random effect model : moyenne et intervalle de confiance pour l’ensemble des articles.
On observe d’après la figure 8, deux articles se démarquant des autres : Benot et al. (2013) et Gedan et Bertness (2009). On remarque également que pour 21 publications sur 29, l’intervalle de confiance s’étend entre des valeurs positives et négatives. Impossible donc de quantifier réellement l’effet. Néanmoins, on remarque que les études avec le plus de variabilité ont le moins de poids (plus le carré est grand plus le poids est important) dans le calcul de la moyenne totale. Cela explique pourquoi nous pouvons faire des conclusions quant à la moyenne globale. Les modèles à effet aléatoire ont également été calculés pour les valeurs d’indice de Shannon-Weaver (11 articles) et d’équitabilité (9 articles). Le nombre d’études étant limité pour ces deux indices, il est difficile de connaitre la véracité des effets observés. Les graphiques en forêt pour ces indices sont disponibles en annexe (3a et 3b). Le tableau 2a représente les valeurs d’indices pour ces modèles. Le tableau 2b, donnent les valeurs d’indices calculées pour les modèles à effet mixte.
Le funnel plot (annexe 2c) montre que les données des articles ont des erreurs standards assez hétérogènes comprises entre 0 et 0.16. Les articles sont répartis de manière homogène autour de la moyenne. Deux publications sont isolées avec des erreurs standard d’environ 0.3.
D’après le tableau 2a, on observe que le réchauffement induit une baisse significative de la richesse spécifique de 6.4% [exp(-0.0662)], mais n’a pas d’effet significatif sur l’indice de Shannon et l’équitabilité.
Les modèles à effets mixtes ont intégré les co-variables que sont la différence de température entre le traitement réchauffé et non réchauffé (T), la durée d’expérimentation (D) et la surface d’échantillonnage (P). En observant les modalités (tab. 2b), on constate que la durée d’expérimentation a un effet significatif (D : p-val QM = 0.0049) sur la différence de richesse spécifique. Le seul facteur « durée » pourrait même exprimer environ 54% de la variabilité du modèle. En combinant les modalités durée et surface d’expérimentation, on observe une
18
différence significative de richesse spécifique (DxP : p-val QM = 0.0002) et que ces deux modalités pourraient exprimer environ 87% du modèle. En combinant les trois modalités (ΔT, durée d’observation et surface d’expérimentation), on obtient également un résultat significatif (TxDxP : p-val QM = 0.0038) avec potentiellement plus de 78% de la variabilité expliquée. Il semble donc que ces facteurs aient effectivement un rôle dans la différence de richesse spécifique sous l’effet du réchauffement. Cependant, la différence de température et la surface d’observation n’ont pas été démontrées comme jouant un rôle à elles seules mais plutôt de manière combinée avec d’autres facteurs (respectivement p-val QE = 0.0313 et 0.164).
Pour les indices de Shannon-Weaver et d’équitabilité, du fait du faible nombre d’articles et de la forte variabilité des petits échantillons, les données sont beaucoup plus difficiles à interpréter.
Modèle Moyenne LRR IC zval pval t² I² (%) H²
SR CvW -0.0662 0.03 -3.7176 0.0002 0.0025 65.73 2.92
H CvW 0.0169 0.04 0.7578 0.4486 0.0006 11.01 1.12
E CvW -0.0333 0.10 -0.6594 0.5097 0.0159 73.58 3.78
Tableau 2a : Valeurs d’indices statistique pour les modèles à effet aléatoire pour la richesse spécifique (SR CvW), pour l’indice de Shannon-Weaver (H CvW) et pour l’indice d’équitabilité (E CvW). Moyenne LRR : moyenne des valeurs de logarithme népérien de ratio de réponse : IC : intervalle de confiance ; pval : valeur p ; t² : quantité d’hétérogénéité ; I² : pourcentage de variabilité dans la taille d’effet calculé via l’échantillon, pouvant être attribué à l’hétérogénéité de l’effet réel ; H² : ratio de la variabilité totale sur la variabilité de l’échantillon.
Indice éco. Modalité t² p-val QE p-val QM R² (%)
SR T 0.027 0.0313 0.7813 0 SR D 0.0012 0.1397 0.0049 53.76 SR P 0.0026 0.0164 0.3751 0 SR TxD 0.0014 0.1909 0.0247 45.86 SR TxP 0.0029 0.0559 0.6876 0 SR DxP 0.0003 0.6907 0.0002 87.47 SR TxDxP 0.0006 0.6386 0.0038 78.51 SR Y 0.0023 0.0260 0.2547 6.67 H' Y 0.0015 0.4172 0.7671 0 E Y 0.1419 0.0011 0.6684 0
Tableau 2b : Valeurs d’indice statistique pour les modèles à effet mixte. Indice éco. : indice écologique : SR : richesse spécifique ; H' : indice de Shannon-Weaver ; E : équitabilité. Modalités : T : différence de température entre l’ambiant et le réchauffement (i. e. ΔT); D : logarithme népérien de la durée d’observation en jours ; P : logarithme népérien de la surface d’échantillonnage en m² ; TxD : combinaison ΔT et durée d’observation; TxP : combinaison ΔT et surface d’échantillonnage; DxP : durée d’observation et surface d’échantillonnage ; TxDxP : ΔT durée d’observation et surface d’échantillonnage; Y: année de publication; t² : quantité estimée d’hétérogénéité résiduelle ; p-val QE : valeur p du test sur l’hétérogénéité résiduelle ; p-value QM : test sur les modérateurs ; R² : pourcentage de variance expliqué par la modalité.
V.
DISCUSSION
A. Effets de la fauche
Modèle à effet aléatoire
L’étude de prairies de différentes compositions, gestion et endroits du monde via la méta-analyse a donné des résultats significatifs. Les modèles à effet aléatoire ont démontré que la fauche avait un impact significatif sur le nombre d’espèces et la biomasse. Faucher une prairie par an peut induire dans la plupart des cas une augmentation de la richesse spécifique et en parallèle, une baisse de la biomasse. Ces tendances avaient déjà été montrées (Gaujour et al., 2011), donc nos résultats semblent cohérents. Cependant, il faut noter que les intervalles de confiances restent assez importants par exemple, si en moyenne le nombre d’espèces augmente de 25% avec une fauche, l’intervalle de confiance s’étend de 14% à 37%.