Deep-sea corals : a new oceanic archive
234
0
0
Texte intégral
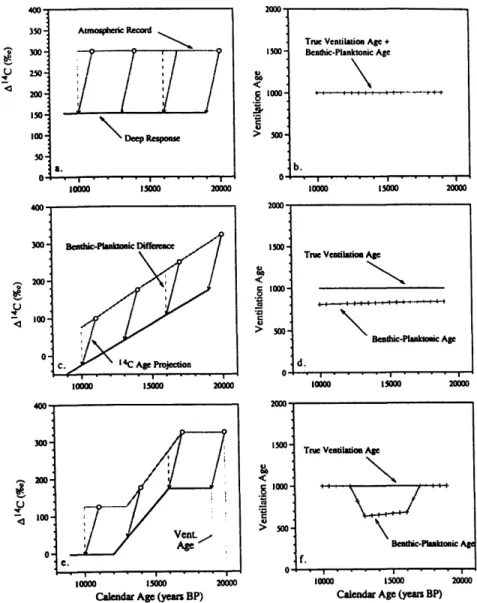
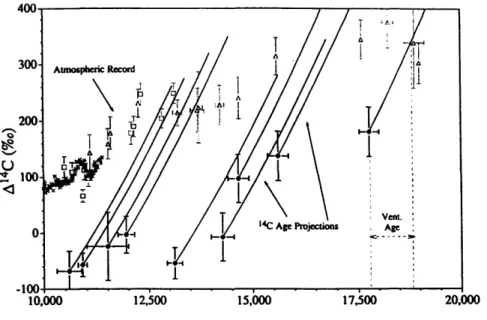
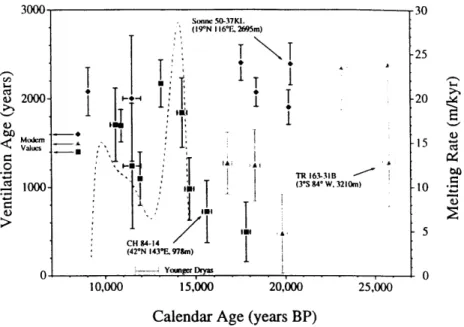
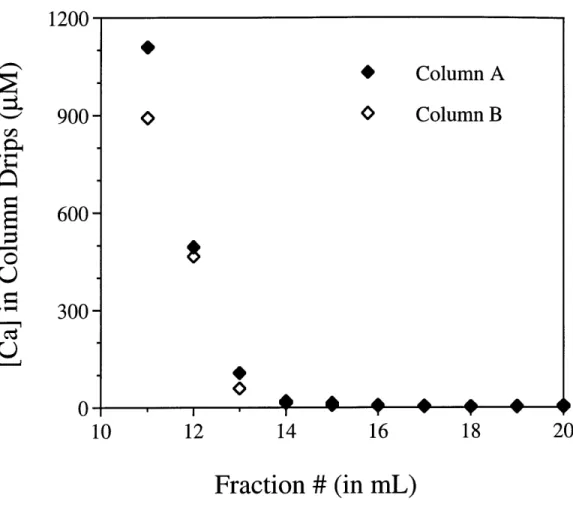
Figure


+7


Documents relatifs
