Depth-resolved particle-associated microbial respiration in the northeast Atlantic
Texte intégral
Figure
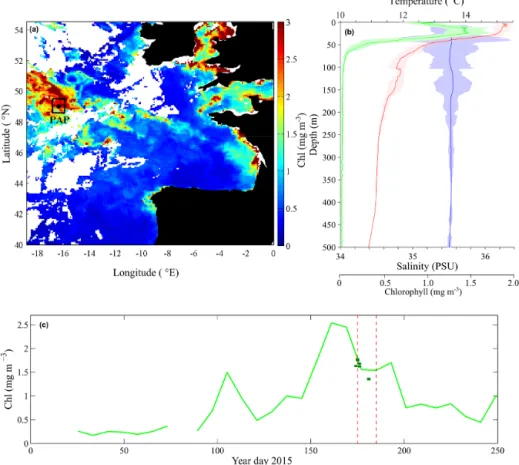

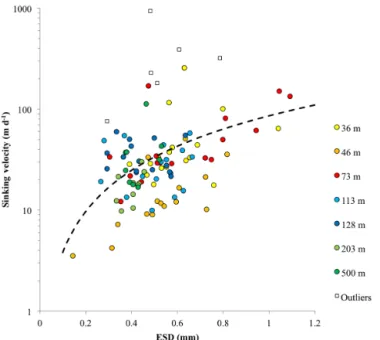
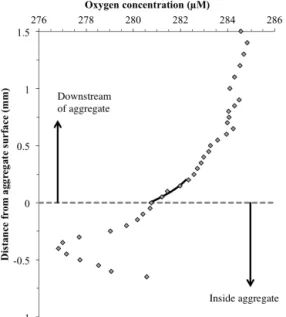
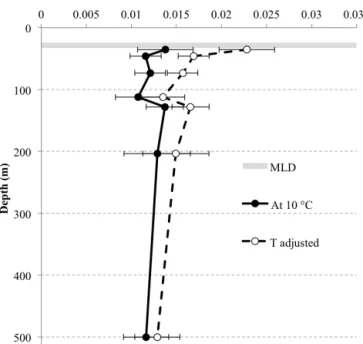
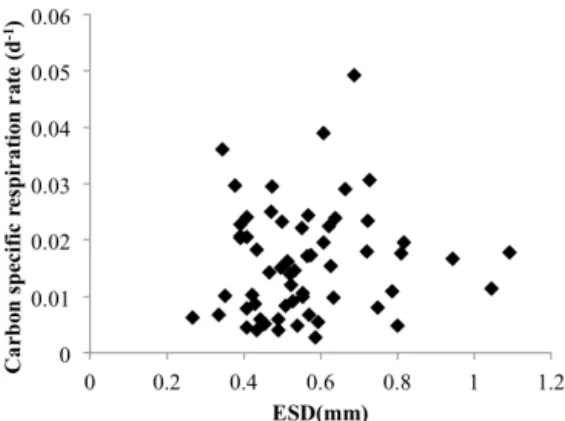
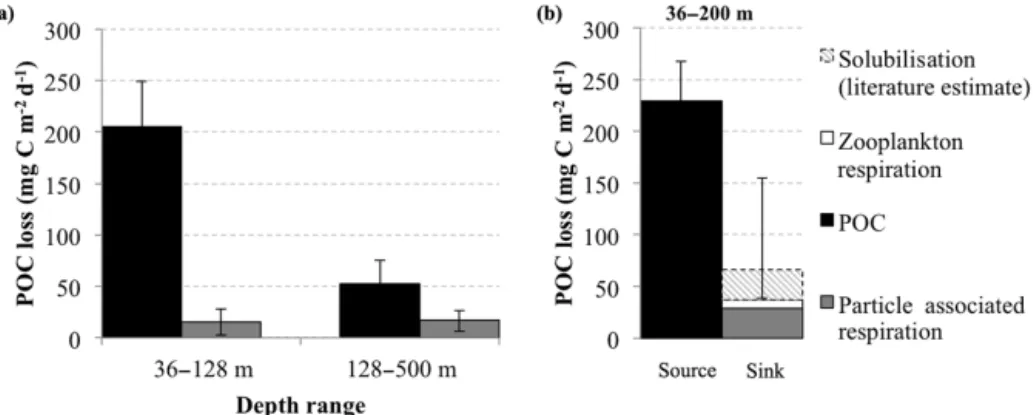
Documents relatifs
Fifty percent of labile organic carbon is exported by the biological pump and 50% by subduction; 98% of the total carbon flux below the mixed layer is due to carbon
Section four is devoted to the numerical simulations, with the impact of the light model on the singular control structure, the optimization of a periodic 1-day cycle, optimization
- Temperature increased respiratory activity until optimum temperature then declined - More decomposed peat decreased the amount of microbes but not respiratory activity - Q 10
SST: Sea-surface temperature; MLD: mixed layer depth; ELD: euphotic layer depth; Tchl a: total chlorophyll a; TOC: total organic carbon; GCP: gross communityproduction; DCR:
Il est donc difficile d’imaginer, dans le cas de productions de verre beaucoup plus importantes (de quelques kilos à une tonne) comme celles des verres anciens, que l’ensemble du
Une thyroïdectomie totale a été associée à un curage ganglionnaire du groupe VI chez 2 patients (patients n°1 et 2), et une lobectomie thyroïdienne a été associée à un
Recopie puis complète les pyramides suivantes afin que le nombre contenu dans une case soit le produit des nombres contenus dans les deux cases situées en dessous de lui..
Je suis un enfant terrible ou pas éduqué Maman est dans sa carrière, émancipée Papa aussi bien préoccupé par soi Et moi je suis là, sous leur toit. Je les aime, comme
