Ca²+ mechanisms of synaptic integration and plasticity in inhibitory interneurons
360
0
0
Texte intégral
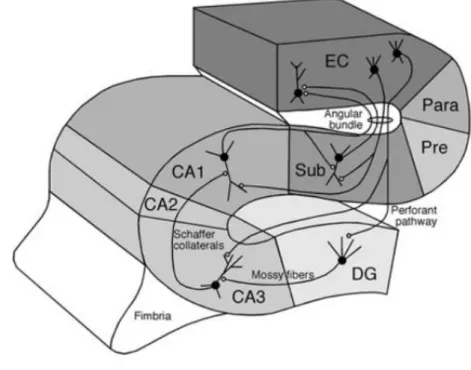
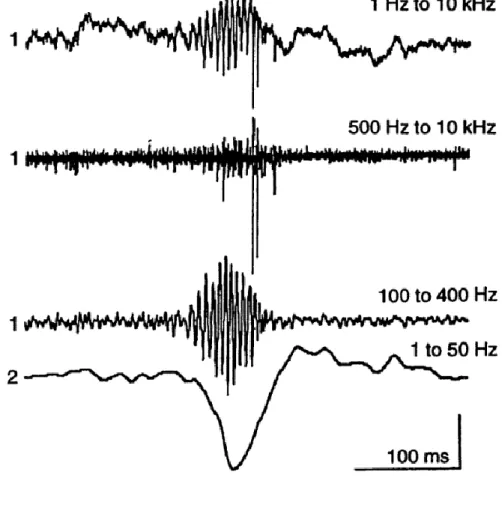
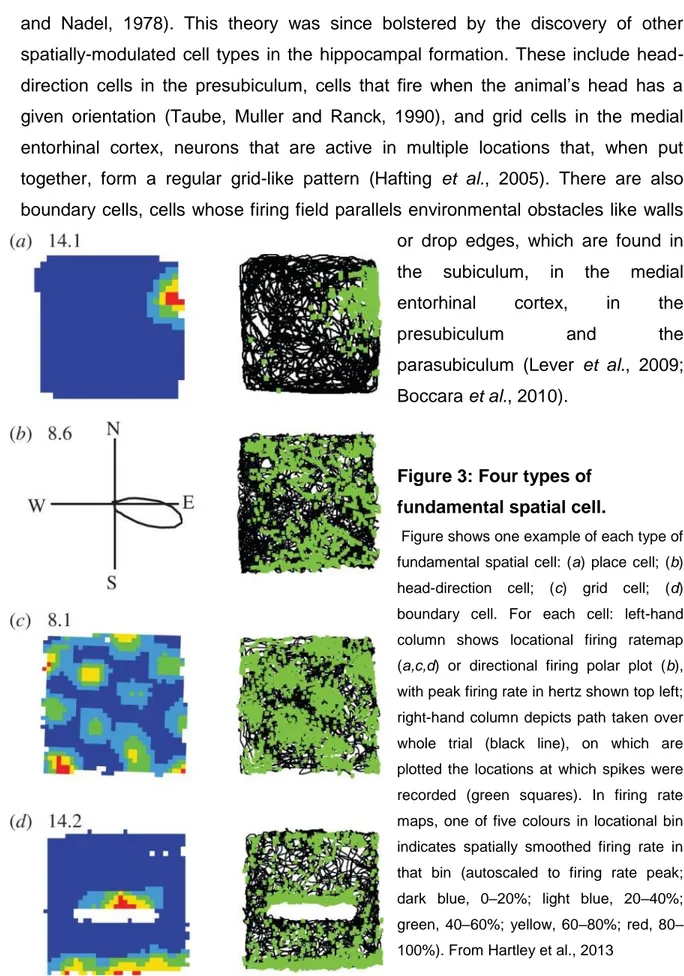
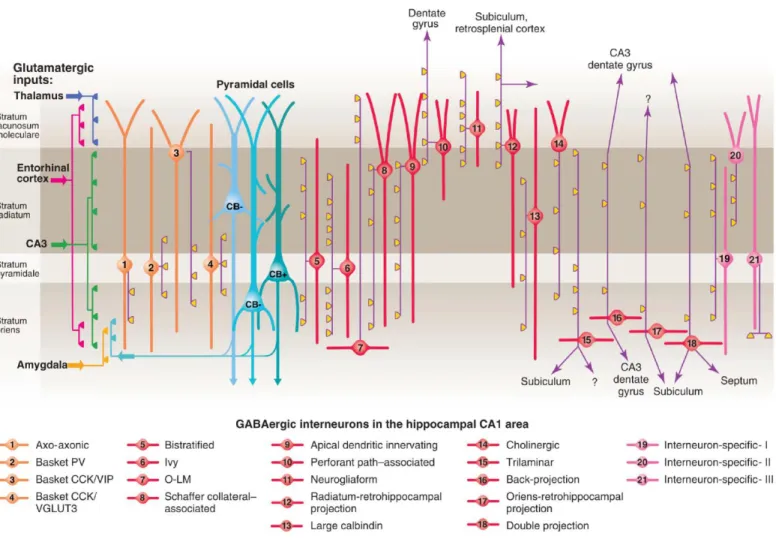
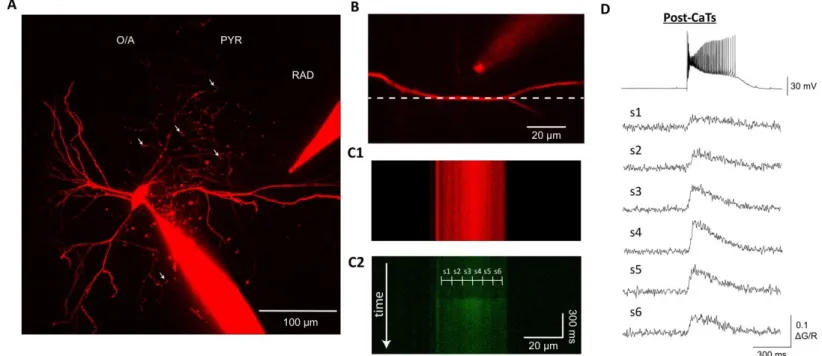
Figure
+7
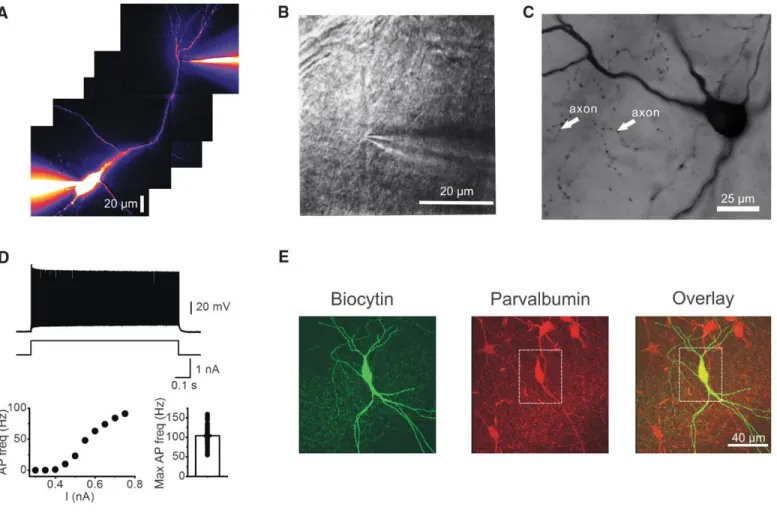
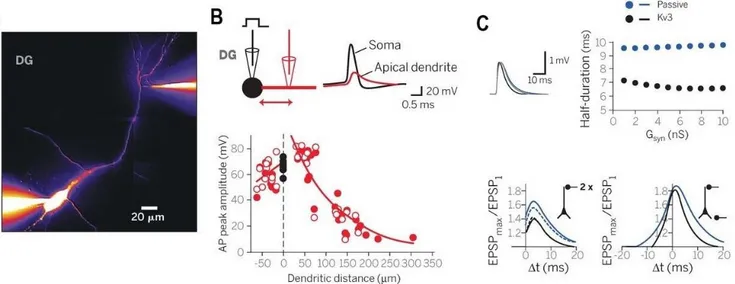

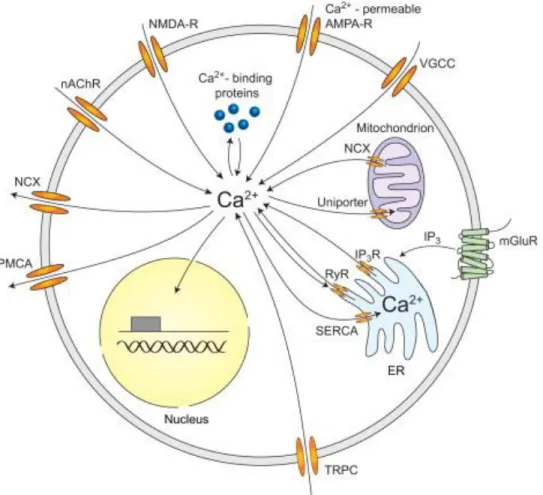
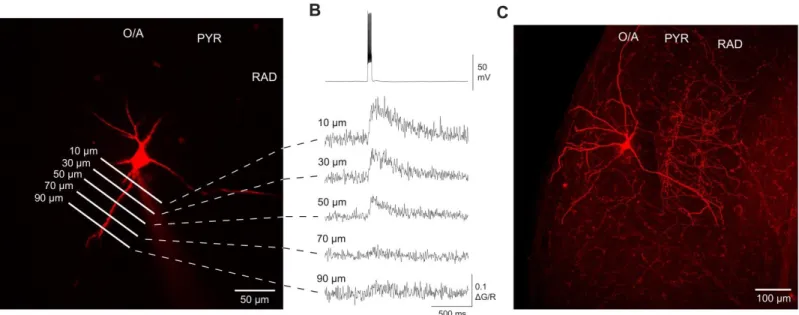
Documents relatifs