Développement d'une méthode analytique pour
l'évaluation des propriétés de déformabilité des
globules rouges
Mémoire
Jonathan Robidoux
Maîtrise en biochimie - avec mémoire
Maître ès sciences (M. Sc.)
Développement d'une méthode analytique pour
l'évaluation des propriétés de déformabilité des
globules rouges
Mémoire
Jonathan Robidoux
Sous la direction de :
Steve Charette, directeur de recherche
Danny Brouard, codirecteur de recherche
Résumé
La médecine transfusionnelle est un rouage important du système de santé canadien. Au Québec, la banque de sang Héma-Québec gère l’ensemble des étapes de la collecte de sang et la livraison des produits sanguins aux hôpitaux. Depuis quelques années, l’altération de la déformabilité des globules rouges observée durant leur entreposage dans les banques de sang est revenue à l’avant-plan des discussions en raison de l’accessibilité grandissante de nouvelles technologies permettant de mieux caractériser cette propriété. L’objectif de ce projet, réalisé au sein d’Héma-Québec, consistait à développer une approche microfluidique novatrice permettant la mesure de l’évolution de la déformabilité des globules rouges durant leur entreposage dans une banque de sang. Le suivi de la déformabilité de six concentrés de globules rouges entreposés à T = [2-8°C] durant 42 jours a permis de déterminer que les performances de la méthode microfluidique surpassent celles d’une méthode comparative également développée dans le cadre de ce projet. Certaines inconsistances observées lors de l’application de la méthode microfluidique ont subséquemment mené à une phase d’optimisation. Bien que certains aspects de la méthode doivent encore être améliorés, les résultats de ce projet laissent présager que la déformabilité pourrait éventuellement servir de marqueur de qualité des globules rouges au même titre que les marqueurs les plus éloquents actuellement pris en compte à Héma-Québec.
Abstract
Transfusion medicine is an important part of the Canadian health care system. In Quebec, the Héma-Québec blood bank manages all stages from blood collection to delivery of blood products to hospitals. In recent years, the alteration of red blood cells deformability observed during storage in blood banks has returned to the forefront of discussions because of the increasing accessibility of new technologies to better characterize this property. This project objective was to develop, within Héma-Québec, a novel microfluidic approach to measure the evolution of red blood cells deformability during blood bank storage. Monitoring the deformability of six red blood cells concentrates stored at T = [2-8 °C] for 42 days determined that the performance of the microfluidic method surpassed that of a comparative method, which was also developed as part of this project. Some inconsistencies observed during the application of the microfluidic method have subsequently led to an optimization phase. Although some aspects of the method still need to be improved, the results of this project suggest that deformability could possibly serve as a quality marker of red blood cells in the same way as the most eloquent markers do.
Table des matières
Résumé ... ii
Abstract ... iii
Table des matières ... iv
Liste des figures ... vi
Liste des tableaux ... vii
Liste des abréviations ... viii
Remerciements ... ix
Avant-propos ... x
Introduction ... 1
Chapitre 1 – Bases théoriques et pratiques de la physiologie du globule rouge ... 4
1.1 Physiologie du globule rouge in vivo ... 4
1.1.1 Transport de l’oxygène ... 5
1.1.2 Déformabilité du globule rouge ... 7
1.1.3 L’érythrophagocytose ou la mort régulée des globules rouges ... 10
1.2 Physiologie du globule rouge in vitro ... 11
1.2.1 Causes des lésions d’entreposage ... 12
1.2.2 Lésions d’entreposage connues... 14
1.2.3 Impact transfusionnel des lésions d’entreposage ... 15
1.3 Méthodes de caractérisation des propriétés de déformabilité des globules rouges ... 17
1.3.1 Méthodes d’analyse unicellulaire ... 17
1.3.2 Méthode d’analyse de masse... 19
1.3.3 L’analyse unicellulaire à haut rendement ... 19
1.3.4 Méthodes d’analyse microfluidique ... 20
Chapitre 2 – Prévenir la variation instrumentale liée à l’utilisation du dispositif MVA ... 30
2.1 Nature du problème ... 30
2.2 Solutions envisageables ... 30
2.3 Développement d’un étalon de référence ... 31
2.4 Preuve de concept : Étalon de référence ... 34
Chapitre 3 – Développement d’une méthode comparative de mesure de la déformabilité des globules rouges ... 36
Chapitre 4 - Development of a Flow Standard to Enable Highly Reproducible Measurements of Deformability
of Stored Red Blood Cells in a Microfluidic Device ... 39
4.1 Résumé ... 39
4.2 Abstract ... 40
4.3 Introduction ... 41
4.4 Material and method ... 43
4.4.1 Blood donation and transformation ... 43
4.4.2 The MVA method ... 43
4.4.3 Measurement of RBC deformability over storage... 45
4.4.4 Measurement of biochemical and hematological parameters ... 45
4.4.5 Statistical analysis ... 46
4.5 Results ... 46
4.5.1 MVA method characterization ... 46
4.5.2 Deformability analysis – NMF and MMFI ... 47
4.5.3 Biochemical and hematological analysis ... 49
4.6 Discussion ... 50
4.6.1 Velocity measurement to interpret deformability ... 50
4.6.2 Storage lesions ... 52
Chapitre 5 – Discussion ... 54
5.1 Limites de l’étude... 54
5.1.1 Différence entre les culots globulaires de l’étude et ceux transfusés cliniquement. ... 54
5.1.2 L’implication de la valeur mesurée ... 55
5.2 Amélioration de l’étalon de référence ... 56
5.2.1 Limites de l’étalon de référence #1 ... 56
5.2.2 Développement et caractérisation d’un nouvel étalon de référence ... 59
5.2.3 Limites du nouvel étalon de référence ... 64
5.3 Perspectives ... 68
Conclusion ... 70
Bibliographie ... 71
Liste des figures
Figure 1 – Schématisation de l’hémoglobine. ... 5
Figure 2 – Courbe de dissociation de l’hémoglobine et de l’oxygène. ... 6
Figure 3 – Structure du cytosquelette et de la membrane cellulaire d’un GR. ... 8
Figure 4 – Formation de microvésicules. ... 11
Figure 5 – Évolution morphologique d’un GR durant l’entreposage. ... 15
Figure 6 – Méthodes de mesure unicellulaire de la déformabilité des GR. ... 18
Figure 7 – Principe de la photolithographie. ... 20
Figure 8 – Réseaux de déformation en parachute. ... 21
Figure 9 – Réseau de goulots d’étranglement. ... 24
Figure 10 – Configuration de la puce microfluidique AMVN. ... 25
Figure 11 – Représentation de GR en rouleau par adhésion au niveau de leur surface plane. ... 26
Figure 12 – Influence de l’aggrégabilité et de la déformabilité des GR sur la viscosité d’un échantillon. .... 28
Figure 13 – Relation entre pression et débit relatifs... 35
Figure 14 – Hémolyse selon les conditions de microfiltration. ... 37
Figure 15 – MVA microfluidic network. ... 42
Figure 16 – MVA signal and analysis. ... 45
Figure 17 – IS ability to correct for method variability. ... 47
Figure 18 – RBCs raw and relative data analysis. ... 48
Figure 19 – Deformability measurements. ... 48
Figure 20 – Effet de l’obstruction partielle du réseau du MVA sur le débit mesuré. ... 57
Figure 21 – Effet des agrégats stationnaires sur le débit mesuré. ... 58
Figure 22 – Comparaison de l’oscillation du débit de l’ER1 et d’un échantillon de GR. ... 58
Figure 23 – Effet de la raréfaction des microsphères. ... 61
Figure 24 – Correction du signal analytique à l’aide de l’ER2. ... 63
Figure 25 – Variations ponctuelles et majeures du signal durant une acquisition. ... 64
Figure 26 – Augmentation du débit lors de l'utilisation de puces n'ayant pas tempéré. ... 65
Figure 27 – Obstruction partielle causée par des microsphères de taille non désirées. ... 67
Liste des tableaux
Tableau 1 - Détermination de la concentration en microsphères et en glycérol nécessaire pour obtenir un
ER dont le débit est représentatif d’un échantillon de GR. ... 33
Tableau 2 – RCCs quality markers evolution during storage. ... 49
Tableau 3 – NMF week-to-week significant differences. ... 49
Tableau 4 – NMF related markers ... 50
Tableau 5 – Caractéristiques et appréciation visuelle des solutions de microsphères testées lors du développement de l’ER2. ... 60
Liste des abréviations
2,3-DPG : 2,3-Diphosphoglycérate
AMVN : Artificial microvascular network (Réseau microvasculaire artificiel) ATP : Adénosine triphosphate
CG : Culot globulaire DésoxyHb : Désoxyhémoglobine DI : Indice de déformabilité EI : Indice d’élongation ER : Étalon de référence GR : Globule rouge Hb : Hémoglobine Htc : Hématocrite OxyHb : Oxyhémoglobine MétHb : Méthémoglobine
MS/V : Ratio de membrane cellulaire sur le volume cellulaire MVA : Microvascular analyzer (Analyseur microvasculaire) PBS : Phosphate buffered saline (Tampon phosphate salin) PDMS : Polydiméthylsiloxane
PE : Phosphatidyléthanolamine
PLM : Microsphère de polystyrène de latex PS : Phosphatidylsérine
ROS : Complexes réactifs oxygénés SAGM : Saline-adénine-glucose-mannitol VCM : Volume cellulaire moyen
Remerciements
Il est coutume de dire que l’important, ce n’est pas la destination, c’est le voyage. La réalisation de ce projet de maîtrise s’est effectivement avérée être un voyage des plus enrichissant et amusant. Il n’en reste pas moins qu’on apprécie arriver à destination.
Je tiens tout d’abord à remercier mon codirecteur de recherche, Danny Brouard, pour m’avoir soutenu durant ce périple, mais également pour m’avoir encouragé à sortir de ma zone de confort dans la réalisation de tâches diverses et complexes, le tout m’ayant permis de développer de solides compétences. Merci de m’avoir permis de prendre part à la vie de professionnel de recherche en m’accordant ta confiance dans la gestion de nos collaborations et en me permettant d’assister notamment au congrès de l’AABB. Vous m’avez encouragé, Marc Cloutier et toi, à emprunter la voix de la recherche scientifique, et je vous en serai éternellement
reconnaissant.
Je tiens également à remercier tous les membres de l’équipe de recherche opérationnelle qui m’ont offert un support inconditionnel tout au long de ce processus. Vos compétences dans le domaine de la recherche scientifique n’ont d’égal que la chaleureuse ambiance régnant au sein de votre équipe. Vous m’avez accueilli à bras ouverts et j’ai été choyé d'avoir pu passer deux belles années à vos côtés.
Mention honorable également à Steve Charette, directeur de maîtrise, qui aura toujours su se rendre rapidement disponible malgré les nombreux chapeaux qu’il peut arborer dans une même journée. Ce fut un réel plaisir d’avoir la chance de te côtoyer.
Et finalement, on ne saurait rester sain d’esprit dans le monde de la recherche sans l’aide d’une muse pour détourner nos rêves des réflexions scientifiques qui hantent nos nuits. Merci, Gabrielle Nolet, de m’avoir soutenu émotionnellement, psychologiquement et financièrement durant ces longues années. Tu as porté ta croix, il est maintenant l’heure pour moi de rapporter le bacon à la maison.
Avant-propos
Ce mémoire contient, entre autres, un article scientifique, rédigé en anglais, se trouvant au Chapitre 4. L’article a été soumis en ligne au journal Transfusion en date du 15 août 2019 (ID : Trans-2019-0652). L’article a été accepté conditionnellement à l’ajout de modifications suggérées par les réviseurs en date du 7 octobre 2019. La version retrouvée dans ce mémoire correspond à la version révisée en date du 9 janvier 2020, version qui sera soumise incessamment. Les numéros des références, de figures et de tableaux ont été adaptés à la facture de ce mémoire. Je, Jonathan Robidoux, suis premier auteur de cet article. J’ai rédigé l’entièreté de l’article qui a été par la suite révisé par Audrey Laforce-Lavoie1, Steve J. Charette2, Sergey S. Shevkoplyas3,
Tatsuro Yoshida4 et Danny Brouard1.
1) Héma-Québec, Affaires Médicales et Innovation, Québec (QC), Canada, G1V 5C3
2) Université Laval, Département de Biochimie, Microbiologie et Bioinformatique, Québec (QC), Canada, G1V 0A6
3) Université de Houston, Département d’Ingénérie Biomédicale, Houston (TX), USA, 77204 4) Hemanext, Lexington (MA), USA, 02421
Introduction
Le terme « Banque de sang » (Blood Bank), initialement utilisé par l’américain d’origine hongroise Bernard Fantus en 1937, est aujourd’hui universellement utilisé pour identifier les entités assurant la gestion des produits sanguins devant servir à la transfusion sanguine humaine.1 Au Québec, c’est l’organisme à but non
lucratif Héma-Québec qui voit à répondre avec efficience aux besoins de la population québécoise en produits sanguins de qualité. Pendant la dernière année, ce sont un peu plus de 350 000 dons qui ont été effectués à travers les divers sites de collecte organisés et administrés par Héma-Québec.2
Il est généralement bien compris qu’un don de sang prélevé en collecte permettra la transfusion de produits sanguins à une personne dans le besoin. Dans les faits, il existe cependant de nombreuses étapes entre le don de sang et la transfusion de produits sanguins. Le sang se compose principalement de plasma, de globules rouges (GR), de plaquettes et de cellules immunitaires. Par des procédés de centrifugation et de filtration, il est possible d’isoler les composants sanguins en produits sanguins labiles.3 La transfusion de l’un
ou l’autre des produits sanguins labiles permet de répondre aux besoins spécifiques des différents receveurs tout en diminuant les risques de réactions transfusionnelles indésirables par l’élimination notamment des cellules immunitaires présentes dans le sang total. La séparation du sang total permet l’entreposage de chacun des produits sanguins en conditions optimales. Le plasma, dépourvu de cellules, peut être congelé et conservé pendant une année. Les concentrés de GR, également appelés culots globulaires (CG), demeurent propres à la transfusion pour une période de 42 jours s’ils sont conservés à une température qui avoisine T = 4 °C. Les plaquettes, qui ont tendance à s’agréger lorsque soumises à des variations de température, sont regroupées pour former le concentré plaquettaire qui peut être conservé sept jours, lorsque maintenu à température ambiante sous agitation continue.
En plus d’assurer la collecte et la distribution de produits sanguins, Héma-Québec a également le mandat de mener des travaux de recherche scientifique, ce qui en fait un acteur important du système de santé québécois. À cette fin, la direction de la recherche opérationnelle à la vice-présidence aux affaires médicales et à l’innovation remplit le rôle d’évaluateur des nouvelles pratiques et des technologies émergentes de la médecine transfusionnelle dans le but d’améliorer la productivité de l’organisme de même que la qualité et la sécurité des produits. Par exemple, un nouveau procédé de séparation du sang total en produits sanguins labiles doit être évalué avant d’être entériné par Héma-Québec puis intégré au niveau des opérations courantes. L’évaluation vise le suivi de différents marqueurs de qualité durant l’entreposage des produits de manière à vérifier le respect des critères de qualité définis par Santé Canada. Dans le cas des CG, les marqueurs sont mesurés à différents moments pendant la période d’entreposage de 42 jours. À terme, il doit être démontré, entre autres, que le taux d’hémolyse, définit comme étant la fraction de globules rouges (GR)
détruits ayant déversé son contenu au niveau du plasma (in vivo) ou du milieu d’entreposage (in vitro), doit être inférieure à 0,8%, et ce pour 95% des CG analysés.3 Bien qu’ils ne figurent pas parmi les critères de
qualité émit par Santé Canada, d’autres marqueurs de qualité peuvent également être mesurés, toujours dans le but d’optimiser les méthodes existantes de traitement et de proposer des produits de meilleure qualité. Aujourd’hui, il n’est pas rare d’observer plusieurs procédés de traitement différents coexister au sein d’une même banque de sang. Ceux-ci permettent l’obtention de CG de qualité équivalente selon les marqueurs de caractérisation actuels. Par souci d’amélioration continue de la qualité des produits, le développement d’outils novateurs pour mieux caractériser la qualité des CG figure parmi les objectifs de recherche de la direction opérationnelle.
La déformabilité des GR a longtemps été considérée comme une propriété ayant le potentiel d’être révélatrice de l’état général d’un CG durant l’entreposage. Cependant, les conclusions discordantes d’études sur le sujet semblent suggérer que les méthodes de mesures actuelles ne sont pas adaptées au suivi des variations de la déformabilité des GR durant l’entreposage. De récentes avancées dans la fabrication de dispositifs microfluidiques ouvrent à nouveau le débat sur la possibilité de mesurer et de considérer la déformabilité des GR comme marqueurs de qualité des CG. Devant pareille opportunité, un projet de recherche ayant comme principal objectif le développement et la caractérisation d’une méthode microfluidique de mesure de la déformabilité des GR a été élaboré.
Hypothèse du projet :
Un outil microfluidique adéquat permettra d’interpréter la déformabilité des GR et de déceler des variations quantifiables de cette propriété durant l’entreposage de ses GR sous les conditions retrouvées dans une banque de sang.
Objectif principal du projet :
Développer et optimiser une approche microfluidique pour la caractérisation des propriétés de déformabilité des globules rouges.
Objectifs secondaires du projet :
1. Développer un étalon de référence permettant un étalonnage de l’approche microfluidique.
3. Comparer le comportement de certains marqueurs de qualité des globules rouges reconnus par Santé-Canada aux résultats de déformabilité obtenus à partir de l’approche microfluidique.
Dans ce mémoire, les bases théoriques de la physiologie des GR et les méthodes existantes pour la caractérisation de leurs propriétés de déformabilité sont d’abord exposées. Ensuite, les travaux préliminaires ayant mené à la réalisation des objectifs secondaires 1 et 2 sont détaillés. Le chapitre 4 traite, au moyen d’un article scientifique, des performances de l’approche microfluidique et de son rapport aux autres marqueurs de qualité des GR suivis durant leur entreposage. Finalement, les travaux ayant suivi ceux rapportés dans l’article scientifique sont présentés avant de conclure avec une discussion portant sur les perspectives d’avenir de la technologie développée.
Chapitre 1 – Bases théoriques et pratiques de la
physiologie du globule rouge
L’étude des GR dans un milieu contrôlé nécessite la compréhension des mécanismes fondamentaux régulant leur vie et leur survie dans leur milieu naturel. De ce fait, la physiologie du GR in vivo est abordée avec une attention particulière aux propriétés physiologiques qui lui confèrent ses remarquables propriétés de déformabilité. Il est ensuite question des changements physiologiques observés pendant la période d’entreposage des CG dans les banques de sang. Les forces et limitations des méthodes permettant la mesure de la déformabilité des GR sont discutées avant la présentation de l’approche novatrice développée dans le cadre du projet.
1.1 Physiologie du globule rouge in vivo
Le système circulatoire sanguin se constitue d’un réseau de vaisseaux sanguins qui permet notamment aux nutriments, aux déchets métaboliques, aux intervenants du système immunitaire et aux messagers du système endocrinien de circuler dans le corps humain.4 Il permet également aux GR de circuler et
d’acheminer l’oxygène des poumons vers les tissus, puis au retour, le dioxyde de carbonedes tissus vers les poumons. Un être humain adulte possède approximativement 25 milliards de GR en circulation qui représentent, en moyenne, entre 40% et 50% du volume sanguin.5-8 Ces GR, également connus sous le nom
d’hématies et d’érythrocytes, ont une durée de vie moyenne de 120 jours, à la suite de laquelle l’accumulation de nombreuses altérations physiologiques mène à leur élimination. Ce phénomène permet le remplacement quotidien des GR désuets par des GR fraîchement formés par érythropoïèse. Ceci signifie qu’un individu moyen présente une population hétérogène de GR âgés entre 1 et 120 jours.
L’érythropoïèse est définie comme étant le processus de différenciation des GR à partir des cellules souches de la moelle osseuse.9 Au cours de ces étapes de différentiation, les cellules souches contenant tous les
organites généralement retrouvés dans une cellule (noyau, mitochondries, réticulum endoplasmique, cytosquelette, etc.) se transforment en une cellule privée d'organites à l’exception d’un cytosquelette retrouvé uniquement sous la surface interne de la membrane cellulaire. Le cytoplasme est principalement constitué d’hémoglobine (Hb), et dans une moindre mesure, d’une variété de protéines enzymatiques nécessaires au fonctionnement du GR désormais incapable d’en synthétiser davantage étant dépourvu de noyau. Il est important de noter que les mitochondries, grandes productrices d’énergies par voie aérobie, sont également absentes des GR matures. Ceci prévient une utilisation significative de l’oxygène transporté à des fins métaboliques. Les composantes du GR seront décrites en fonction de leur rôle respectif au niveau du
1.1.1 Transport de l’oxygène
L’oxygène est une molécule réactive qui est grandement utilisée dans le corps humain aux fins de la respiration aérobie, permettant la formation d’adénosine triphosphate (ATP) par la conversion de l’énergie obtenue de la dégradation des nutriments absorbés. Cependant, sous certaines conditions, l’oxygène peut s’associer à d’autres molécules et former des complexes réactifs oxygénés (ROS, reactive oxygen species) qui peuvent causer d’importants dommages cellulaires.10 Afin d’éviter les dommages oxydatifs, 98% de
l’oxygène présent dans le sang est confiné à l’intérieur des GR alors qu’environ 2% se retrouvent sous forme dissoute dans le plasma sanguin.11 C’est grâce à leur impressionnante quantité d’Hb cytoplasmique, celle-ci
constituant près de 95% de leur volume cellulaire5, que les GR parviennent à assurer un transport efficace et
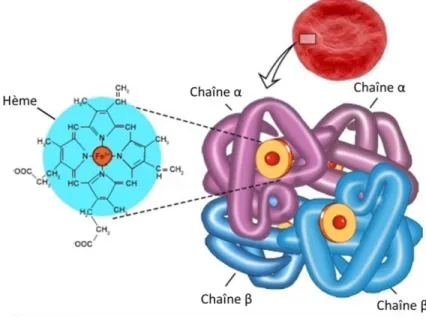
contrôlé de l’oxygène. L’Hb est une protéine tétramérique formée généralement de deux sous-unités α et de deux sous-unités β (Figure 1).12 Des variantes de la sous-unité β peuvent être retrouvées chez l’humain, mais
elles ne seront pas décrites dans le cadre de ce mémoire. Chacune des sous-unités porte un cofacteur hème, caractérisée par la présence d’un ion de fer généralement sous forme oxydée (Fe2+). Ce sont ces ions ferreux
qui permettent la liaison et la rétention de l’O2. La majorité du CO2 est également transportée en faisant
intervenir l’hème, bien qu’une partie considérable soit également dissoute dans le plasma sanguin.13
Figure 1 – Schématisation de l’hémoglobine. Une molécule d’Hb se compose de
deux chaînes α et de deux chaînes β portant chacune un groupement hème. Chaque hème est porteur d’un atome de fer permettant la liaison et le transport de l’oxygène. La capacité du fer à porter une molécule d’O2 dépend de son état
ionique, lui qui peut être retrouvé sous forme Fe2+, Fe3+ ou Fe4+ selon l’état du
cytoplasme et l’intégrité de la molécule d’Hb. Figure adaptée de https://bodytomy.com/structure-of-hemoglobin.
La grande efficacité d’un GR pour réaliser le transport de l’oxygène réside, en partie, dans la dynamique particulière d’association et de dissociation de l’O2 et de l’Hb, un processus qui ne consomme pas d’énergie.12
L’Hb est une molécule allostérique, ce qui signifie qu’elle change de conformation lorsqu’elle se lie à l’oxygène. Lorsque la pression en oxygène du milieu est faible, l’Hb ne porte aucune molécule d’oxygène et se trouve sous forme désoxyhémoglobine (désoxyHb) en conformation tendue. Cette conformation est maintenue par des liaisons ioniques, des ponts hydrogènes et des interactions hydrophobes qui sont stabilisés par la présence de molécules de 2,3-diphosphoglycérate (2,3-DPG) entre les sous-unités β de l’Hb. Sous une plus forte pression en oxygène, l’affinité de l’Hb pour l’oxygène supplante les liens maintenant la conformation tendue, ce qui permet la liaison d’une première molécule d’oxygène. Cette liaison induit un changement conformationnel de l’Hb qui passe à l’état relâché. Sous cette forme, l’Hb peut se lier à de nouvelles molécules d’oxygène jusqu’à concurrence de quatre. L’Hb saturée en oxygène est appelée oxyhémoglobine (oxyHb). Ce phénomène est illustré par la courbe de dissociation sigmoïdale de l’oxygène à la Figure 2.
Figure 2 – Courbe de dissociation de l’hémoglobine et de l’oxygène. A) La forme sigmoïdale de la
courbe laisse transparaître; une réticence de l’Hb à lier l’oxygène à des pressions d’O2 faibles (PO2 <
10 mm de Hg), une aisance exponentielle à lier l’oxygène causée par le passage de la conformation tendue à relâchée (PO2 = 10 à 50 mm de Hg), une zone de plateau lorsque la majorité de l’Hb est déjà saturée en oxygène (PO2 > 50-60 mm de Hg). B) Visualisation de l’oxyHb et de la désoxyHb. Figure adaptée de https://www.medicalexamprep.co.uk/
Concrètement, les GR en circulation dans le sang possèdent à la fois des molécules d’oxyHb et de désoxyHb. Lorsque ceux-ci circulent dans un tissu, comme un tissu musculaire par exemple, la faible pression d’O2
Seuls les tissus pulmonaires possèdent une pression environnante en O2 suffisamment élevée pour induire la
fixation de l’oxygène par les désoxyHb. C’est donc la concentration en oxygène de l’environnement du GR qui influence la fixation et le largage de l’oxygène. Dans un même ordre d’idée, le pH peut également jouer un rôle au niveau de l’affinité de l’Hb pour l’oxygène. La forte concentration en ions H+ associée à un faible pH incite
l’Hb à adopter une conformation tendue et désoxygénée en favorisant la formation de liaisons ioniques entre certains de ses acides aminés. Par conséquent, la forte concentration en ions H+ observée au niveau des
tissus hypoxiques favorise le largage d’oxygène en facilitant la transition conformationnelle d’oxyHb à désoxyHb. La capacité du GR de libérer ou de fixer l’oxygène sans coût énergétique est donc modulée par l’environnement de l’Hb qui favorisera, par allostérie, la fixation dans un milieu riche en oxygène au pH normal tel que les poumons, et le largage dans un milieu pauvre en oxygène au faible pH comme un tissu musculaire au travail.
1.1.2 Déformabilité du globule rouge
Le GR présente une forme discoïdale caractérisée par une épaisseur comprise entre 2,5 et 3 μm et un diamètre de 7 à 8 µm.14 Afin d’acheminer l’oxygène aux tissus, les GR se doivent de traverser les capillaires
sanguins, qui peuvent présenter un diamètre contraignant pouvant atteindre 3 μm. Les GR parviennent à circuler par les capillaires grâce à leur grande capacité à se déformer qui résulte d’un amalgame de propriétés. De manière générale, on distingue ces propriétés en trois grandes catégories, soit le ratio de membrane de surface par rapport au volume cellulaire (MS/V), l’organisation du cytosquelette et la viscosité du cytoplasme.
1.1.2.1 Le ratio de membrane de surface par rapport au volume cellulaire
Le volume moyen d’un GR est d’environ 90 fL (1 fL = 10-9 µL) pour une surface membranaire totale d’environ
140 um2.14 S’il était de forme sphérique plutôt que discoïdale, un GR ayant toujours une surface membranaire
de 140 um2 aurait un volume de 155 fL. À cet égard, on pourrait comparer un GR à un ballon de plage
dégonflé, qui s’avère être beaucoup plus déformable et malléable que son homologue bien gonflé. Ce grand ratio de surface par rapport au volume (MS/V) permet au GR d’adopter sa forme discoïdale typique. Le ratio MS/V procure aux GR la résilience nécessaire pour répondre aux contraintes physiques rencontrées dans le système circulatoire. Lorsque soumise à une force orientée dans une seule direction, la membrane discoïdale du GR adopte une forme plutôt ovale. Par déplacement du surplus de membranes, le GR peut s’allonger pour atteindre plus du double de sa longueur initiale entraînant une diminution remarquée de sa largeur. Bien qu’un faible ratio MS/V soit nécessaire pour permettre au GR de se déformer, c’est l’organisation particulière de son cytosquelette qui lui confère tonus et flexibilité.
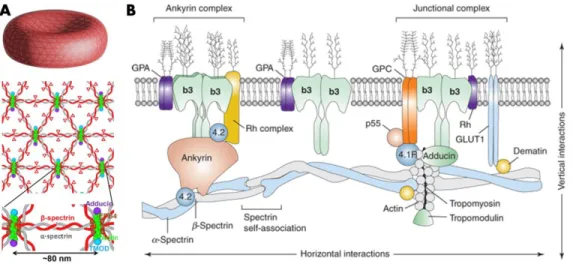
Figure 3 – Structure du cytosquelette et de la membrane cellulaire d’un GR. A) Le
cytosquelette du GR est aussi appelé squelette membranaire puisqu’il recouvre la surface interne de la membrane cellulaire du GR, mais est absent du cytoplasme. L’arrangement particulier de l’α-spectrine, de la ß-spectrine et des autres protéines du cytosquelette lui confère tonus et flexibilité. B) Le cytosquelette supporte la membrane et régule la distribution des protéines membranaires.12
L'interaction horizontale décrit l’arrangement des protéines du cytosquelette entre elles et l'interaction verticale décrit l'arrangement entre protéines du cytosquelette et protéines membranaires. Figure 3.A adaptée de https://www.cell.com/cms/attachment/ce7e45ef-39d4-47df-af4e-c1e9e69d62b7/fx1_lrg.jpg
1.1.2.2 L’organisation du cytosquelette
Le cytosquelette du GR recouvre la surface interne de la membrane cellulaire et est complètement absent du cytoplasme.15 Le cytosquelette se compose d’α-spectrine et de ß-spectrine qui s’associent en dimères
hélicoïdaux et en tétramères pour ensuite former une succession de segments d’hélices répétées. Ces tétramères sont liés entre eux par des protéines d’actines, qui forment ensemble une mosaïque flexible (Figure 3). Les ankyrines sont des protéines intermédiaires permettant la liaison entre les spectrines du cytosquelette et la protéine transmembranaire bande 3. En plus d’arrimer la membrane au cytosquelette, le complexe spectrine, ankyrine et bande 3 joue un rôle dans l’organisation latérale de la membrane cellulaire. Les glycophorines, de forme A, B, C ou D sont également des protéines transmembranaires pouvant servir de point d’ancrage pour le cytosquelette. C’est à l’aide de la protéine 4.1b et d’un complexe protéique inconnu que les glycophorines se lient aux actines du cytosquelette. Pour d’autres types de cellules, la présence d’organites cytoplasmiques requiert le soutien du cytosquelette omniprésent à l’intérieur de la cellule.16 C’est
donc l’absence de cytosquelette en son centre et l’arrangement particulier des hélices de spectrines qui, à l’image de ressorts, procurent aux GR leur flexibilité tout en soutenant la forme discoïdale observée au repos. En circulation dans le corps humain, le GR ne dispose que d’un bref instant pour répondre aux contraintes spatiales de même qu’aux collisions avec les autres cellules en circulation. L’arrangement du cytosquelette et
cellulaire défini de l’anglais comme étant du tank treading, ce qui augmente la résilience des GR face aux contraintes rencontrées dans la circulation sanguine.17
1.1.2.3 La viscosité du cytoplasme
Le ratio MS/V et l’organisation particulière du cytosquelette permettent à la membrane de se mouvoir dans toutes les directions en réponse aux contraintes externes, ce qui rend la déformabilité du GR vulnérable à la viscosité de son cytoplasme qui peut offrir une résistance supplémentaire.15 Outre l’eau, l’Hb est le principal
constituant cytoplasmique d’un GR. La concentration de l’Hb se situe normalement entre 27 et 35 g/dL et n’influence pas de manière considérable la viscosité du cytoplasme. Étant dépourvu de noyau, le GR n’a pas la capacité de produire de l’Hb au cours de sa vie. Par contre, un dérèglement homéostatique causé par des défaillances au niveau des pompes ioniques peut entraîner une déshydratation du GR. La quantité totale en Hb demeure stable, mais la diminution du volume du GR a pour effet d’augmenter sa concentration en Hb. Lorsqu’elle devient supérieure à 37 g/dL, la viscosité du cytoplasme augmente de manière exponentielle devenant un facteur important influençant les propriétés de déformabilité du GR.
1.1.2.4 Le rôle indirect des réserves d’énergie
Bien qu’elles ne soient pas directement utilisées dans le processus de déformabilité, les réserves d’énergie du GR, principalement constituées d’ATP, assurent le maintien de l’organisation membranaire, de l’intégrité du cytosquelette et du fonctionnement des pompes ioniques.18,19 Une chute abrupte des réserves d’énergie
n’aurait donc pas un impact instantané sur la déformabilité du GR mais affecterait éventuellement chacune des trois propriétés abordées précédemment. Par conséquent, les réserves d’ATP ont un effet indirect sur la déformabilité des GR. Puisque les réserves d’énergie des GR évolueront durant leur entreposage, il est important d’en décrire la source et d’en comprendre les effets.
La glycolyse est le principal processus métabolique actif chez le GR. Ce processus permet la dégradation anaérobie du glucose et la production d’intermédiaires énergétiques.20 Chaque molécule de glucose permet
au GR de produire deux molécules d’ATP. En plus d’intermédiaires comme le 2,3-DPG, la dégradation du glucose produit le pyruvate qui peut être réduit en lactate par la nicotinamide adénine dinucléotide (NADH) si sa concentration cytoplasmique le permet. La concentration en NADH dépend de la présence de la méthémoglobine (métHb); une forme altérée de l’Hb portant le fer sous forme ferrique (Fe+3) plutôt que
ferreuse (Fe2+). Le NADH permet la réduction préférentielle de la métHb en Hb d’où l’incidence de la
concentration en métHb sur la quantité de NADH disponible pour la réduction du pyruvate en lactate. Ce concept est d’incidence modérée dans le cas d’un GR in vivo puisque le lactate et le pyruvate sont expulsés de la cellule pour être métabolisés ailleurs dans le corps humain. Il en est autrement dans le cas de l’entreposage, où les déchets métaboliques excrétés par les GR subsistent dans le milieu nutritif.
1.1.3 L’érythrophagocytose ou la mort régulée des globules rouges
Le concept de mort cellulaire programmée, nommé apoptose, est retrouvé chez un grand nombre de types cellulaires au niveau des organismes multicellulaires.21 Une cellule endommagée ou en fin de vie utile peut
enclencher un mécanisme d’autodestruction par la synthèse de protéines spécifiques ce qui permet un renouvellement cellulaire bénéfique à l’organisme. Les GR en circulation ne peuvent pas synthétiser de nouvelles protéines puisqu’ils sont privés de noyau. Leur élimination fait principalement intervenir des acteurs externes qui parviennent à reconnaitre les GR les plus altérés via leurs marqueurs de sénescence.22 Au cours
de sa vie de 120 jours, un GR est soumis à de nombreux stress tels que l’acidité élevée et les dommages oxydatifs. Ces stress affectent l’intégrité et l’organisation du cytosquelette et de la membrane cellulaire du GR. Il en résulte une désorganisation des points d’ancrage et de la distribution des composants membranaires qui causent de petits détachements localisés entre la membrane cellulaire et le cytosquelette.23 Ces
détachements membranaires représentés à la Figure 4 résultent en de multiples bourgeonnements, c’est-à-dire à la formation et au largage de microvésicules (MVs) par le GR. Les MVs sont définies comme étant formées de membranes cellulaires encapsulant du contenu cytoplasmique. Elles se forment lorsque le détachement membranaire produit une protubérance suffisamment grande pour permettre à la membrane de se refermer sur elle-même, se détachant ainsi du reste de la membrane cellulaire. Ce processus permet aux GR d’éliminer l’Hb dénaturée, les protéines endommagées et les sections de membrane portant un nombre considérable de marqueurs de sénescence. Cependant, la libération de MVs affecte le ratio MS/V nécessaire aux GR pour maintenir une bonne déformabilité et induit en partie la transition d’un érythrocyte de forme discoïdale à sphérique.19 L’altération du cytosquelette s’accentue avec le temps sous l’effet des dommages
oxydatifs alors que le processus de vésiculation perd son efficacité à larguer les segments cellulaires les plus altérés en raison du manque de surplus membranaire. Cela cause éventuellement l’accumulation de marqueurs de sénescence exposés à la surface du GR.
L’altération significative du ratio MS/V et de l’intégrité du cytosquelette affecte grandement la déformabilité des GR, ce qui mène à leur rétention dans les fentes endothéliales (1 µm × 0,5 µm) de la pulpe rouge de la rate.14
Les GR coincés dans la pulpe rouge de la rate sont exposés aux macrophages, c’est-à-dire à des cellules du système immunitaire dont le rôle principal est la dégradation de cellules ou de structures cellulaires. C’est la reconnaissance des marqueurs de sénescence, tels que la phosphatidylsérine18 (PS) et la
phosphatidyléthanolamine24 (PE), ainsi que l’absence du marqueur d’inhibition de la phagocytose CD4725 qui
permet aux macrophages de reconnaitre les GR altérés et de déclencher le mécanisme de phagocytose. La dégradation à l’intérieur des macrophages permet la récupération des hèmes et du fer des GR pour leur réutilisation lors de l’érythropoïèse.9 Ce processus d’élimination permet de retirer de la circulation les GR qui
pour les échanges gazeux.26,27 Le mécanisme d’érythrophagocytose permet donc d’assurer le retrait des GR
dont la fonction principale est altérée tout en recyclant les molécules de fer et d’hème.
Figure 4 – Formation de microvésicules. Toutes perturbations des interactions
entre les protéines du cytosquelette et les protéines transmembranaires sont susceptibles de causer la formation de microvésicules par altération directe des liens verticaux ou par regroupement d’éléments membranaires non liés au cytosquelette si l’altération affecte plutôt l’organisation horizontale du cytosquelette. Figure tirée de
https://www.frontiersin.org/files/Articles/366081/fphys-09-00703-HTML/image_m/fphys-09-00703-g001.jpg
1.2 Physiologie du globule rouge in vitro
Les banques de sang isolent les GR des autres composants sanguins de manière à favoriser leur préservation durant l’entreposage et à réduire le risque de réactions immunitaires lors des transfusions. À Héma-Québec, les pratiques d’entreposage actuelles permettent la conservation d’un CG pendant une période de 42 jours en conditions réfrigérées (T = 2-8 °C).3 Bien que la transfusion d’un CG en fin d’entreposage soit tout de même
bénéfique pour un receveur, il est reconnu que les GR puissent être de moins bonne qualité en raison des altérations subies durant cette période. L’ensemble des changements subits et exprimés par les GR sont regroupés sous l’étiquette de lésions d’entreposage. Certains facteurs causant ses lésions d’entreposage sont connus, bien que la ramification complexe des mécanismes impliqués rend difficile l’établissement de cause à
effet. Pour cette raison, les causes connues et les lésions d’entreposage répertoriées sont présentées séparément.
1.2.1 Causes des lésions d’entreposage
À première vue, il pourrait sembler logique d’estimer que la population hétérogène composée de GR âgés de 1 à 120 jours continue simplement de vieillir durant l’entreposage. Au bout des 42 jours d’entreposage, cette population soustraite au processus d'érythrophagocytose serait composée d’individus âgés de 43 à 162 jours, dont certains seraient fortement altérés. Il faut cependant prendre en considération que la vie d’un GR in vivo diffère de celle d’un GR entreposé dans le cadre des opérations d’une banque de sang.28 In vivo, les GR
vivent dans le plasma sanguin à température corporelle (T ≈ 37 °C) et sont contraints de circuler sans relâche dans les vaisseaux sanguins en côtoyant une panoplie d’autres cellules (endothéliales, immunitaires, sanguines). L’intégrité des GR est fréquemment contrôlée lors de leur passage dans le foie et dans la rate ce qui mène à l’élimination des GR altérés. Sous forme de CG dans les banques de sang, les GR ne sont pas soumis aux mêmes stress mécaniques et leur métabolisme est ralenti par les faibles températures d’entreposage (T = 2-8°C). Ils sont suspendus dans une solution nutritive et ne sont plus soumis au processus d’élimination des GR altérés, ce qui bouleverse le concept de durée de vie des GR. Ce nouvel environnement froid où les GR sont immobiles dans une solution nutritive non renouvelée n’est pas parfait, et cause sa propre ribambelle d’altérations physiologiques. L’ensemble de ses altérations serait causé par des dommages oxydatifs et/ou par l’altération des mécanismes métaboliques.
1.2.1.1 Les dommages oxydatifs
L’oxygène est une molécule réactive qui, bien qu’essentielle au fonctionnement du corps humain, peut réagir de manière incontrôlée avec un grand nombre de molécules.5 Les GR assurent essentiellement un transport
cloisonné de l’oxygène qui ne se retrouve autrement qu’en très faible quantité sous forme de gaz dissout dans le sang.11 Durant l’entreposage des CG, les GR sont scellés dans une poche de plastique qui n’est pas
totalement étanche à la diffusion des molécules d’oxygène. Le milieu nutritif d’entreposage des GR accumule l’oxygène à une concentration plus élevée que celle retrouvée dans le plasma sanguin, ce qui affecte la concentration intracellulaire en oxygène. Comme mentionné précédemment, l’Hb alterne entre la forme oxygénée et désoxygénée selon la concentration en O2 du milieu. Cependant, lorsque la concentration en
oxygène devient trop importante, l’oxyHb est oxydée pour former ce que l’on appelle la métHb, selon la réaction suivante :
2-Puisque la metHb ne possède pas la capacité de transporter l’oxygène, sa présence n’est pas souhaitable in vivo et cette molécule est généralement réduite en oxyHb par la « méthémoglobine réductase ». Durant l’entreposage, l’activité de cette enzyme est cependant ralentie par la basse température, ce qui favorise l’accumulation de metHb dans les GR entreposés. De plus, la production de la molécule O2- enclenche une
cascade métabolique de dommages oxydatifs qui mène à la production de molécules oxygénées hautement réactives (ROS) que sont la « ferryl hemoglobin » (porteuse de Fe4+) et le radical hydroxyle (•OH). Ces
réactions moléculaires s’accélèrent après la deuxième semaine d’entreposage. Le pouvoir oxydatif important des ROS affecte d’importantes protéines structurales telles que l’ankyrine et la spectrine ainsi qu’un grand nombre d’enzymes. Certaines des protéines ainsi oxydées auront désormais tendance à se lier au cytosquelette, troublant son association avec la membrane cellulaire et causant des changements morphologiques nuisibles à la déformabilité des GR.
1.2.1.2 L’entreposage en milieu froid et acide
La seconde grande cause derrière l’apparition des lésions d’entreposage est le changement d’environnement des GR.5 Ce nouvel environnement se caractérise notamment par une basse température, un milieu fermé
représenté par la poche d’entreposage et un milieu nutritif plus simple que le plasma sanguin. Durant l’entreposage réfrigéré, le métabolisme cellulaire des GR est ralenti. Les enzymes de la glycolyse anaérobie continuent de dégrader lentement le glucose retrouvé dans le milieu nutritif. L'apport énergétique de ce métabolisme résiduel ne suffit pas pour remplacer la totalité de l’énergie dépensée par la cellule, ce qui mène à une diminution de la concentration intracellulaire en ATP. La faible température d’entreposage réduit également l’activité d’autres enzymes importantes. C’est le cas de la méthémoglobine réductase, mais également celui de la scramblase et de la flippase qui permettent normalement le maintien de l’organisation des phospholipides membranaires. Ce maintien permet de prévenir le largage de vésicules par détachement localisé de la membrane et du cytosquelette. Ces deux enzymes, étant ATP-dépendantes, voient leur activité résiduelle diminuer davantage avec la déplétion des réserves d’ATP observée durant l’entreposage. La pompe ionique Na-K, le transporteur ionique ATP-dépendant le plus important chez le GR, est inactivée à la température d’entreposage ce qui perturbe l’équilibre ionique nécessaire au maintien de l’activité des autres transporteurs ioniques. La chute de la concentration en ATP affecte également la pompe de l’ion calcium (Ca2+). Le débalancement du gradient électrochimique qui en découle affecte l’organisation du cytosquelette
puisque les ions Ca2+ sont impliqués dans la stabilisation des liens entre actines et spectrines.15 Le
métabolisme résiduel du GR continu de produire des déchets métaboliques tels que le lactate et le pyruvate. Ces déchets sont normalement excrétés directement dans le sang et sont retirés de la circulation par d’autres organes du corps humain.20 Durant l’entreposage, ces déchets s’accumulent dans le système fermé de la
comme conséquence d’entraver graduellement l’activité enzymatique de protéines impliquées dans la production d’ATP. La solution nutritive la plus utilisée chez Héma-Québec et largement étudiée dans la littérature scientifique est le saline-adénine-glucose-mannitol (SAGM). Le SAGM se compose d’une solution saline isotonique, d’une source d’énergie (glucose), d’une purine pour prévenir la désamination néfaste à la glycolyse et d’une molécule permettant d’amenuiser l’effet de radicaux oxygénés (mannitol). Il diffère du plasma sanguin par l’absence de facteurs antioxydants tels que l’acide urique.29 Bien que la formule de la
solution nutritive puisse varier, c’est la diminution du pH et la concentration en oxygène qui jouent un rôle prépondérant sur la qualité des GR durant l’entreposage, ce qui n’est pas facile à prévenir par des changements au niveau de la solution nutritive. Il est pertinent de mentionner que le remplacement hebdomadaire de la solution nutritive permet d’amenuiser l’apparition des lésions d’entreposage. Il existe également des solutions nutritives plus complexes qui permettent de recouvrir certaines caractéristiques ayant été altérées durant l’entreposage.30 L’utilisation de ces solutions, dans leurs états actuels, ne représente
toutefois pas une avenue opérationnellement viable pour une banque de sang qui doit gérer un volume important de dons par année.
1.2.2 Lésions d’entreposage connues
Les causes pouvant altérer la qualité d’un CG ont une influence ramifiée et interconnectée.5,18,19,31 Il est de ce
fait difficile d’identifier une cause unique pour une altération particulière. Par exemple, l’inhibition partielle des enzymes de la glycolyse par le froid et par l’acidification du pH cause une diminution constante des réserves d’ATP des GR. Cette importante diminution affecte le travail de nombreuses enzymes ATP-dépendantes. Parmi le lot, les flippases et les scramblases deviennent moins efficaces ce qui accentue l’exposition de la PS, et l’activité réduite de la méthémoglobine réductase nuit à la prévention des dommages oxydatifs. Les pompes ATP-dépendantes sont également affectées, ce qui nuit à l’état d’hydratation du GR et à la conservation de l’équilibre ionique nécessaire au maintien de l’intégrité cytosquelettique. Un autre exemple de pluralité de conséquences est l’étendue des dégâts oxydatifs causés par la concentration élevée d’oxygène dissous. La génération de ROS affecte de nombreuses protéines enzymatiques et structurales ainsi que certains phospholipides. Dans l’ensemble, on considère que les lésions d’entreposage affectent la capacité des GR à maintenir un niveau optimal d’ATP et de 2,3-DPG. L’altération de l’intégrité cytosquelettique et de l’organisation membranaire induit des changements morphologiques graduels présentés à la Figure 5, la perte d’asymétrie membranaire ainsi que le rassemblement de PS et de protéines bande 3.32 Il en découle une perte
d’adhésion de la membrane au cytosquelette ce qui mène au largage de MVs par les GR. Cette perte de membrane amplifie le phénomène de changement morphologique et affecte le ratio MS/V.18
Figure 5 – Évolution morphologique d’un GR durant l’entreposage. Un GR sain présente une
forme discoïdale et porte le nom de discocyte. Les altérations physiologiques subies durant l’entreposage mènent généralement à des dissociations ponctuelles entre le cytosquelette et la membrane cellulaires des GR qui entraînent des détachements visibles par microscopie optique. Les GR sont catégorisés en echninocyte I, II et III et sphéro-echinocyte selon l’importance des spicules causés par les détachements membranaires et la morphologie générale du GR. La diminution du ratio MS/V causé par la formation de MVs mène les GR à adopter une forme sphérique. À ce stade, l'absence de surplus membranaire ne permet plus au GR de se déformer efficacement ou de larguer des MVs. Figure tirée de l’article de Sierra et coll.33
La déformabilité des GR semble être altérée par différents aspects de l’entreposage. La souplesse du cytosquelette est altérée par les dommages oxydatifs et les changements de concentration en Ca2+.19 Le
largage de vésicules induit une perte de membrane qui affecte le ratio MS/V, ce qui induit une transformation morphologique des GR qui passent d’une forme discoïdale à une forme sphérique peu déformable. La viscosité intracellulaire des GR ne change pas significativement durant l’entreposage et ne serait pas en cause dans l’altération de la déformabilité des GR durant l’entreposage. L’ATP n’est pas directement impliquée dans les processus de déformabilité des GR, mais joue un important rôle dans le maintien de l’intégrité cellulaire.14 Dans l’ensemble, la capacité de déformabilité des GR ne peut pas être déduite par la
mesure de marqueurs indirects puisque les relations de cause à effet sont multiples et ne sont pas entièrement connues à ce jour.
1.2.3 Impact transfusionnel des lésions d’entreposage
L’objectif de la transfusion de CG est de restaurer chez un individu l’oxygénation des tissus amenuisée par une variété de conditions, incluant un saignement dû à une blessure, une chirurgie importante ou une condition pathologique causant de l’anémie telle que les thalassémies α et β.34,35 Devant l’état des
y a lieu de se demander si la transfusion d’un produit entreposé depuis près de six semaines aura le même effet thérapeutique que la transfusion d’un produit frais.
L’impact transfusionnel de CG présentant certaines lésions d’entreposage n’est pas simple à établir. La transfusion de CG dans les hôpitaux est nécessaire pour les individus au profil physiologique affecté par la nature de leur condition, ce qui rend extrêmement difficile l’interprétation des indicateurs de santé mesurés des suites d’une transfusion. Par exemple, un individu souffrant d’hémorragie se trouve dans une fâcheuse posture qui pousse déjà son corps à produire une réponse inflammatoire. L’individu en anémie chronique, quant à lui, est souvent en proie à une élimination de GR plus rapide que la normale ce qui s’accompagne d’une hausse de facteurs sanguins liés à l’hémolyse. Il s’avère difficile de distinguer la réponse physiologique liée à la transfusion de GR altérés de la réponse physiologique causée par l’état de l’individu. Néanmoins, certains chercheurs ont tenté de déterminer l’effet de la transfusion de CG plus vieux par une étude statistique des cas de mortalité et de survie des suites de transfusions étant survenus dans les hôpitaux. Les résultats de ces études suggèrent qu’il n’y a pas de corrélation entre des évènements tragiques telles que la mort d’individu et la transfusion de CG plus âgés.36,37 D’autres études ce sont intéressées à des indicateurs moins
radicaux comme le temps passée en hospitalisation à la suite de la transfusion ce qui s’est soldé par une absence de corrélation.38 Il n’est pas étonnant de constater que la pratique transfusionnelle développée depuis
des décennies n’ait pas systématiquement d’impacts négatifs majeurs sur la santé des receveurs.
Des études ciblées ont toutefois permis de déceler certains indicateurs suggérant que la transfusion de CG altérés par l’entreposage pouvait s’avérer moins bénéfique pour le receveur que la transfusion d’un CG frais. Le principal indicateur utilisé pour caractériser le succès d’une transfusion de CG est le taux de GR transfusés ayant survécu dans le corps du receveur 24 heures après la transfusion sanguine. Le taux de survie après 24 heures est inversement corrélé avec le temps d’entreposage et peut atteindre des valeurs d’environ 75% après six semaines d’entreposage.5,39 C’est donc jusqu’à 25% d’un CG transfusé qui sera éliminé par le corps
dans les 24 premières heures suivant la transfusion. L’élimination de ces GR survient principalement par érythrophagocytose ce qui s’explique par l’exposition accrue de facteurs de sénescence et l’altération de la déformabilité des GR observés durant l’entreposage.39 Cette élimination rapide de GR peut causer une
surcharge de certains mécanismes de régulations du système circulatoire sanguin.40 Notamment, l’oxyde
nitrique permettant la vasodilatation de vaisseaux sanguins capillaires est accaparé par les microvésicules et le fer mis en circulation par la transfusion et par l’élimination rapide des GR transfusés.5 De plus, la surcharge
du système d’élimination des GR causée par la transfusion induit le maintien en circulation de GR ayant une déformabilité altérée.30 La capacité de vasodilatation amoindrie des capillaires et la circulation de GR ayant
individus devant recevoir plusieurs CG. Notamment, l’élimination accrue de GR peut provoquer une surcharge du fer dans le sang ce qui peut devenir néfaste pour les organes.42
Les conséquences exactes de la transfusion de CG en fin d’entreposage restent nébuleuses. L’élimination rapide de GR à la suite d’une transfusion est cependant un fait établi et il est probable que la déformabilité altérée des GR en soit partiellement responsable. C’est pourquoi le suivi des propriétés de déformabilité des GR lors de l’évaluation d’un nouveau procédé de séparation du sang total pourrait permettre aux banques de sang de mieux caractériser les culots globulaires et ultimement d’améliorer la qualité du produit.
1.3 Méthodes de caractérisation des propriétés de déformabilité
des globules rouges
N’étant que survolées dans l’article (Chapitre 4), les différentes méthodes de mesure de la déformabilité des GR sont ici reprises plus en détail, avec une attention particulière portée aux méthodes impliquant l’exploitation de concepts microfluidiques. Les méthodes de mesure de la déformabilité des GR sont classées comme étant des analyses unicellulaires, de masse (bulk analysis) ou unicellulaires à haut rendement. Le premier cas de figure regroupe des méthodes visant à quantifier la capacité de déformabilité d’un seul GR à la fois. Cela permet d’étudier la dynamique des membranes en mesurant, par exemple, l’effet d’une altération chimique du milieu sur la membrane d’un GR. L’analyse de masse permet d’étudier le comportement général d’un échantillon de sang ou de GR, et fournit une mesure unique par échantillon. L’analyse unicellulaire à haut rendement regroupe les méthodes permettant d’effectuer une mesure des propriétés de déformabilité de nombreux GR individuels à l’aide de systèmes automatisés. Elle permet ainsi l’obtention rapide de données pour un nombre appréciable de GR (typiquement, n > 1000).
1.3.1 Méthodes d’analyse unicellulaire
Ces méthodes demandent beaucoup de temps et d’habiletés techniques en plus d’être difficilement adaptées pour l’analyse de populations. Elles permettent toutefois de mesurer l’effet d’une modification précise de la composition membranaire ou cytosquelettique dans le cadre d’étude plus fondamentale portant sur les GR.43
L’aspiration à la micropipette est l’une des premières méthodes ayant été utilisée pour quantifier la
déformabilité des GR (Figure 6A).44 Elle vise à aspirer partiellement ou complètement un GR pour en étudier
la dynamique membranaire. L’aspiration partielle ou complète s’effectue dans des micropipettes de 1 à 5 µm de diamètre. D’autres applications consistent à mesurer le temps nécessaire pour qu’un GR retrouve sa forme initiale après avoir été aspiré.
fixée à la lame et au GR alors que la seconde n’est fixée qu’au GR. Le champ magnétique induit par un laser amovible permet de capturer et de déplacer la seconde bille de manière à étirer le GR. Le GR est étiré jusqu’à ce qu’il cherche à reprendre sa forme initiale et que la force de traction induite par sa membrane et son cytosquelette permette à la bille d’échapper au champ magnétique du laser. L’étirement maximal, mesuré par microscopie optique, correspond à l’étirement du GR au moment où la bille s’échappe du champ magnétique et permet l’étude comparative de l’étirement des GR.
Figure 6 – Méthodes de mesure unicellulaire de la déformabilité des GR. A) L’aspiration
à la micropipette peut consister à mesurer la longueur de la protubérance qui pénètre dans l’embout de la micropipette sous différentes pressions négatives. B) Les pinces optiques permettent d’étirer mécaniquement le GR. L’étirement maximal est mesuré au moment où la tension membranaire force la bille à quitter l’emprise du faisceau optique. C) La microscopie de force atomique permet de mesurer en plusieurs points la force nécessaire pour déformer la membrane du GR. Figure 6.A adaptée de https://www.dovepress.com, 6.B adaptée de https://ars.els-cdn.com/ et 6.C adaptée de https://content.iospress.com/
La microscopie de force atomique est une méthode de mesure quantitative de déformation ciblée d’un
matériel ou d’une cellule telle qu’un GR (Figure 6C).44 Ce dernier est fixé sur une lame de laboratoire et est
d’obtenir de l’information au niveau unicellulaire, mais également de distinguer différentes parties de la membrane d’un GR.
1.3.2 Méthode d’analyse de masse
La microfiltration est actuellement la seule véritable méthode d’analyse de masse. Plusieurs adaptations ont
été développées depuis son invention en 1976.45 Généralement, on mesure le temps de filtration d’un volume
précis d’échantillon de GR à un taux d’hématocrite prédéfini à travers un filtre de pores de diamètre inférieur à celui des GR (3 ou 5 µm), sous une pression faible et constante. Cette mesure permet le calcul de l’indice de déformabilité qui est défini comme le volume d’échantillon filtré par minute. La sensibilité du système est grandement limitée par l’obstruction rapide du filtre lorsque la déformabilité des GR est altérée de manière importante ce qui limite son utilité.46
1.3.3 L’analyse unicellulaire à haut rendement
Un rhéoscope se compose d’un microscope inversé muni d’une chambre rhéologique dans laquelle est déposé un échantillon sanguin en suspension dans un milieu de viscosité importante.44 La chambre
rhéologique d’un premier modèle est représentée par deux plaques circulaires transparentes tournant en sens opposés de manière à moduler la force de cisaillement (shear stress) désirée pouvant aller jusqu’à 30 Pa. Les GR adoptent ainsi une conformation ellipsoïdale influencée par leur déformabilité. Un logiciel permet l’analyse de l’indice d’élongation (EI) caractérisé par le rapport de la différence entre la longueur (a) et la largeur (b) sur la somme de la longueur et de la largeur du GR :
EI =(𝑎 − 𝑏) (𝑎 + 𝑏)
Les données d’EI proviennent de GR individuels et sont compilées en un graphique permettant d’apprécier la distribution des valeurs. Il est ainsi possible, par exemple, de raisonner sur la valeur moyenne de déformabilité entre un échantillon de GR en début et en fin d’entreposage. Celle-ci peut être due à une diminution modérée des propriétés de déformabilité pour l’ensemble des GR ou une forte diminution pour une fraction de ceux-ci. Ce type d’instrument peut servir d’outil diagnostique pour certaines pathologies sanguines puisque des GR en forme de faucille (anémie falciforme) ou en forme d’ellipse (elliptocytose congénital) n‘auront pas l’alignement de GR sains sous l’effet du cisaillement induit. D‘autres types de rhéoscopes utilisent un courant linéaire appliqué à une chambre rhéologique en forme de corridor. Une mesure de la déformabilité est obtenue à l’aide de la formule de mesure de l’EI. La principale limitation de ces méthodes est la mesure en deux dimensions d’un phénomène qui se déroule en trois dimensions.46 La force de cisaillement induite permet donc d’observer
1.3.4 Méthodes d’analyse microfluidique
1.3.4.1 Fabrication de dispositifs microfluidiques
Les méthodes microfluidiques requièrent l’utilisation de dispositifs comprenant des canaux dont les dimensions varient de trois à pas plus de quelques centaines de micromètres. Leur confection implique l’utilisation de moules précis fabriqués par photolithographie (Figure 7).47 Lors de l'application de ce procédé,
une plaque recouverte d’une ou plusieurs couches de matière photosensible est exposée à une lumière filtrée par un masque représentant le schéma microfluidique désiré. Le matériel photosensible se solidifie par exposition à la lumière et la partie n’ayant pas été exposée peut être dissoute afin d’obtenir un moule du schéma microfluidique aux dimensions voulues de précision micrométrique.
Figure 7 – Principe de la photolithographie. A) La lumière traverse un masque du réseau de
microcanaux désirés. Une lentille permet de focaliser la lumière filtrée par le masque afin qu’elle atteigne un substrat placé à une distance déterminée. À l’aide de calculs mathématiques, il est possible d’obtenir une image aux dimensions précises sur le substrat. B) La lumière qui traverse le masque atteint la résine placée sur le substrat. Selon la nature photosensible ou photorésistante de la résine, il est possible d’imprimer le schéma ou l’inverse du schéma sur le substrat et ainsi obtenir un moule aux dimensions micrométriques. Figure 7A adaptée de https://www.quora.com/What-are-the-limitations-of-optical-lithography et Figure 7B adaptée de https://en.wikipedia.org/wiki/Photoresist.
Le polydimétylsiloxane (PDMS) est un composé liquide qui est versé sur le moule porteur du schéma microfluidique. Les caractéristiques du PDMS, dont sa polymérisation par la chaleur, permettent le respect des dimensions micrométriques du moule.48 Le dispositif obtenu est transparent et de dimensions stables sur un
apposée sur une lame de microscope préalablement recouverte d’une fine couche de PDMS qui servira de revêtement favorisant l’adhésion du réseau de microcanaux. En effet, aucun adhésif n’est nécessaire puisque la fonctionnalisation de la surface de la lame et de la puce par un traitement au plasma d’oxygène permet d’obtenir deux surfaces fortement polarisées qui adhèrent ensemble.51 La transparence du PDMS permet le
suivi par microscopie du comportement de cellules circulant à travers le réseau de microcanaux. C’est la simplicité du procédé de fabrication des puces de PDMS qui, entre autres, a mené au développement de diverses méthodes microfluidiques pour l’analyse des propriétés de déformabilité des GR. Celles rapportées dans la littérature sont des méthodes d’analyses unicellulaires à haut rendement, et seront décrites plus en détail pour favoriser la compréhension du caractère unique du dispositif « Microvascular Analyzer » (MVA) développé dans le cadre de ce projet.
1.3.4.2 Mesure de la déformation en parachute
Deux méthodes similaires développées par Sun Y. et coll.52 et par Cluitmans et coll.53 permettent d’observer la
transition de passage des GR entre deux chambres vastes séparées par un canal de 8 µm de large pour la première étude et de 7 µm de large pour la seconde (Figure 8).
Figure 8 – Réseaux de déformation en parachute. A) Configuration du réseau microfluidique
développé par Sun et coll.52 Bien que le schéma ne soit pas présenté dans son ensemble, il est
possible d’apprécier le système permettant la circulation du GR et la mesure des paramètres D et L qui représentent respectivement la largeur et la longueur du GR tel que décrit dans la formule de calcul du DI. Les canaux perpendiculaires au canal de perfusion horizontal induisent un courant incitant l’alignement du GR au centre du canal. B) Configuration du réseau microfluidique développé par Cluitmans et coll.53 La principale différence avec le premier
dispositif réside dans l’absence de canaux d’alignement. Les paramètres de mesures B et A correspondent respectivement à la largeur et la longueur tel que décrit dans la formule de calcul du DI.