Production of xylanases by Bacillus polymyxa using lignocellulosic wastes
11
0
0
Texte intégral
Figure



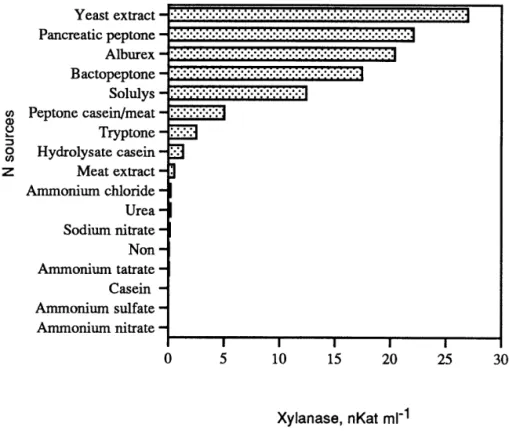
+2
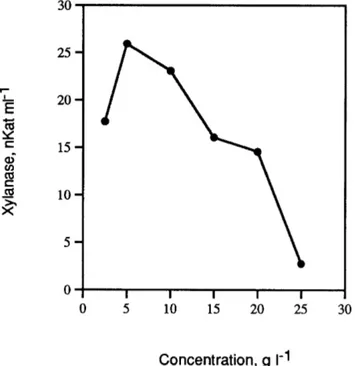
Documents relatifs
