Effet des apports en protéines métabolisables et en
énergie de rations à base de luzerne sur les
performances de vaches en lactation
Mémoire
Jean-Philippe Laroche
Maîtrise en sciences animales - avec mémoire
Maître ès sciences (M. Sc.)
Effet des apports en protéines métabolisables et en
énergie de rations à base de luzerne sur les
performances de vaches en lactation
Mémoire
Jean-Philippe Laroche, agr.
Sous la direction de :
Édith Charbonneau, agr., Ph. D., directrice de recherche
Rachel Gervais, agr., Ph. D., codirectrice de recherche
Résumé
La luzerne contient des fibres peu digestibles et une forte proportion de protéines dégradables. Les vaches laitières pourraient donc bénéficier d’une augmentation de la digestibilité des fibres de la luzerne, ou d’une meilleure adéquation entre l’apport en azote et en énergie disponible dans le rumen. Les cultivars de luzerne présentant une meilleure digestibilité des fibres sont une opportunité d’augmenter l’efficacité de la production laitière, mais aucune étude indépendante n’a testé ces cultivars dans les conditions agroclimatiques du Canada. Par ailleurs, la diminution de l’apport en protéines métabolisables est souvent associée à une diminution de la sécrétion de protéines dans le lait, possiblement causée par un apport réduit en acides aminés essentiels. Les objectifs de l’étude étaient (1) de comparer les performances de vaches laitières nourries avec des rations à base de luzerne, en utilisant un cultivar régulier et un cultivar sélectionné pour sa concentration réduite en lignine ; et (2) d’évaluer l’effet d’une variation de l’apport en énergie sur les performances de vaches laitières alimentées de rations ayant un apport réduit en protéines métabolisables, mais équilibrées pour les principaux acides aminés essentiels limitants. Les cultivars de luzerne n’ont eu aucun effet significatif sur l’ensemble des paramètres mesurés. La diminution de l’apport en protéines métabolisables et le maintien du même bilan énergétique ont permis de maintenir la prise alimentaire et la production laitière corrigée pour l’énergie. Cette stratégie alimentaire a également amélioré l’efficacité d’utilisation de l’azote tout en maintenant la production des composantes du lait et en diminuant l’excrétion azotée. Il est possible de réduire l’apport en protéines métabolisables sans avoir d’effet négatif sur la performance des vaches laitières, à condition de combler leurs besoins en énergie et en acides aminés essentiels. De tels changements permettraient de diminuer l’impact environnemental de la production laitière, en plus de réduire potentiellement les coûts d’alimentation.
Abstract
Alfalfa contains poorly digestible fiber and a high proportion of degradable protein. For these reasons, dairy cows could benefit from an increased digestibility of alfalfa’s fibers, or from a better match between nitrogen and energy supply in the rumen. Alfalfa cultivars with improved fiber digestibility represent an opportunity to increase milk production efficiency, but no independent studies tested these cultivars under the agroclimatic conditions of Canada. Also, decreasing metabolizable protein supply is often associated with a decrease in milk protein yield, possibly caused by a reduced supply of essential amino acids. The objectives of the study were (1) to compare the performance of dairy cows fed alfalfa-based diets, using a regular cultivar and a cultivar selected for its reduced lignin concentration; and (2) to assess the effect of a variation in energy supply on the performance of dairy cows fed diets with a reduced supply of metabolizable protein, but balanced for the main limiting essential amino acids. Alfalfa cultivars had no significant effect on all measured parameters. Decreasing metabolizable protein supply and maintaining the same energy balance allowed cows to maintain their dry matter intake and energy corrected milk production. This feeding strategy also improved the nitrogen use efficiency while maintaining milk component production and reducing nitrogen excretion. Reducing metabolizable protein supply without negative effects on dairy cow performance is possible when energy and essential amino acids requirements are met. Such changes would decrease the environmental impact of milk production, in addition to potentially reduce feeding costs.
Table des matières
Résumé ... ii
Abstract ... iii
Table des matières ... iv
Liste des tableaux ... vi
Liste des figures ... viii
Liste des abréviations françaises ... ix
Liste des abréviations anglaises ... x
Remerciements ... xi
Avant-propos ... xii
Introduction ... 1
Chapitre 1 Revue des travaux antérieurs... 3
1.1 Amélioration de l’utilisation énergétique de la luzerne par l’augmentation de sa digestibilité ... 3
1.1.1 Digestibilité des fourrages ... 3
1.1.1.1 Analyses chimiques corrélées à la digestibilité ... 5
1.1.1.2 Analyses de digestibilité ... 6
1.1.2 Façons d’accroître la digestibilité de la luzerne par la sélection génétique ... 7
1.1.2.1 Approche conventionnelle ... 8
1.1.2.2 Approche transgénique ... 9
1.1.3 Intérêts agronomiques d’une luzerne plus digestible ... 10
1.1.4 Caractéristiques des luzernes plus digestibles ... 12
1.1.5 Effets de la digestibilité des fourrages sur la vache en lactation ... 16
1.1.5.1 Consommation ... 17
1.1.5.2 Production et composition du lait ... 21
1.1.5.3 Santé ... 23
1.1.5.4 Performances économiques et environnementales de la ferme ... 25
1.2 Amélioration de l’utilisation protéique de la luzerne par une diminution de l’apport en protéines métabolisables de la ration ... 28
1.2.1 Besoins azotés des vaches laitières ... 28
1.2.1.1 Protéines non dégradables ... 30
1.2.1.2 Synthèse de protéines microbiennes ... 31
1.2.1.3 Protéines endogènes ... 34
1.2.1.4 Besoins en acides aminés ... 34
1.2.2 Diminution de l’apport en protéines métabolisables ... 38
1.2.2.1 Fermentation ruminale et digestibilité des nutriments ... 39
1.2.2.3 Utilisation de l’azote ... 48
1.2.2.4 Santé et fertilité ... 51
1.2.2.5 Interaction avec l’apport en énergie de la ration ... 52
1.2.2.6 Performances économiques et environnementales de la ferme ... 53
1.3 Hypothèses et objectifs de recherche ... 54
Chapitre 2 Milk production, nutrient digestion, and nitrogen utilization efficiency as affected by metabolizable protein and energy supply in dairy cows fed alfalfa-based diets ... 57
2.1 Résumé interprétatif ... 57
2.2 Interpretive Summary ... 57
2.3 Abstract ... 57
2.4 Introduction ... 58
2.5 Materials and Methods ... 60
2.5.1 Haylage Production ... 60
2.5.2 Cows, Diets and Experimental Design ... 60
2.5.3 Measurements and Sampling... 61
2.5.4 Chemical Analyses ... 62
2.5.5 Statistical Analysis ... 64
2.6 Results and Discussion ... 64
2.6.1 Forages and Diets ... 64
2.6.2 Animal Performance ... 66
2.6.3 Apparent Digestibility and N Balance ... 67
2.6.4 Plasma Metabolites and AA, and Milk Fatty Acid Composition ... 69
2.7 Conclusions ... 70
2.8 Acknowledgments ... 71
2.9 References ... 71
Conclusion ... 89
Liste des tableaux
Tableau 1.1 Valeur nutritive moyenne de trois cultivars de luzerne pendant six dates d’échantillonnage
durant chacun des deux cycles de repousse de l’année d’implantation (2015) à travers six états. ... 13
Tableau 1.2 Valeur nutritive moyenne de quatre cultivars de luzerne pendant six dates
d’échantillonnage durant chacun des deux cycles de repousse de l’année d’implantation (2015) à travers deux états. ... 13
Tableau 1.3 Différentes mesures sur différents cultivars de luzerne cultivés à Becker (BE), Rochester
(ROC), Rosemount (ROS) et Saint-Paul (SP) au Minnesota en 2015 et en 2016. ... 15
Tableau 1.4 Caractéristiques comparées de la luzerne Europe (témoin) et la luzerne
expérimentale 6328P, n=66. ... 16
Tableau 1.5 Sommaire des expériences ayant porté sur la digestibilité de la luzerne à différents stades
de maturité. ... 18
Tableau 1.6 Sommaire des expériences ayant porté sur la digestibilité de cultivars de luzerne. ... 20 Tableau 1.7 Production et composition du lait selon différentes expériences ayant porté sur la
digestibilité de la luzerne à différents stades de maturité. ... 22
Tableau 1.8 Production et composition du lait selon différentes expériences ayant porté sur la
digestibilité de cultivars de luzerne. ... 23
Tableau 1.9 Mesures reliées à la santé des vaches selon différentes expériences ayant porté sur la
digestibilité de cultivars de luzerne. ... 25
Tableau 1.10 Simulation du modèle DAFOSYM sur les résultats économiques reliés à l’augmentation
de la digestibilité de la NDF de la luzerne. ... 27
Tableau 1.11 Composition en acides aminés des bactéries et protozoaires du rumen chez les bovins (g
d’acides aminés/100 g d’acides aminés). ... 33
Tableau 1.12 Proportions optimales de Lys et de Met dans les protéines métabolisables pour maximiser
le rendement ou la concentration en protéine du lait avec le modèle de balancement des rations NRC. ... 37
Tableau 1.13 Effet des apports en protéines métabolisables et de la supplémentation en acides aminés
protégés sur la digestibilité apparente de rations par des vaches laitières. ... 40
Tableau 1.14 Effet des apports en protéines métabolisables, de la supplémentation en urée et en acides
aminés protégés sur la digestibilité apparente de rations par des vaches laitières. ... 41
Tableau 1.15 Effet des apports en protéines métabolisables et de la supplémentation en acides aminés
protégés sur la production et la composition du lait de vaches laitières. ... 46
Tableau 1.16 Effet des apports en protéines métabolisables, de la supplémentation en urée et en acides
Tableau 1.17 Effet des apports en protéines métabolisables et de la supplémentation en acides aminés
protégés sur la production et la composition du lait de vaches laitières. ... 48
Tableau 1.18 Effet des apports en protéines métabolisables et de la supplémentation en acides aminés protégés sur le bilan azoté de vaches laitières... 50
Tableau 1.19 Effet des apports en protéines métabolisables, de la supplémentation en urée et en acides aminés protégés sur le bilan azoté de vaches laitières. ... 51
Table 2.1. Chemical composition of alfalfa haylages ... 77
Table 2.2. Ingredients and chemical composition of experimental diets ... 78
Table 2.3. Estimated protein, EAA and energy balance1 in dairy cows ... 80
Table 2.4. Effect of MP and energy supply on DMI, milk yield and composition, and BW in dairy cows ... 82
Table 2.5. Effect of MP and energy supply on intake and total-tract apparent digestibility of nutrients in dairy cows ... 83
Table 2.6. Effect of MP and energy supply on milk N secretion, urinary and fecal N excretion, and urine purine derivatives (PD) excretion in dairy cows ... 84
Table 2.7. Effect of MP and energy supply on blood plasma metabolites in dairy cows ... 85
Table 2.8. Effect of MP and energy supply on blood plasma AA concentrations (μM) in dairy cows ... 86
Table 2.9. Effect of MP and energy supply on milk fatty acid composition (g/100 g of total fatty acid) in dairy cows ... 87
Liste des figures
Figure 1.1 Représentation graphique d’un modèle cinétique simple de la disparition des fibres dans
le rumen : Les fibres potentiellement digestibles sont représentées par D, le taux fractionnaire de digestion par Kd et le taux fractionnaire de passage par Kp. ... 4
Figure 1.2 Voie de biosynthèse de la lignine. ... 9
Figure 1.3 Qualité et rendement relatif de la luzerne à différents stades de croissance. ... 10
Figure 1.4 Masse de MS (alfalfa forage mass DM) prédite et intervalles de confiance à 95 % (plage
ombragée) pour le cultivar de luzerne réduit en lignine (54HVX41 ; y1) et les cultivars de référence (y2) en réponse aux degrés-jours cumulés (cumulative growing degree days). ... 11
Figure 1.5 Qualité fourragère relative (alfalfa relative forage quality; Moore et Undersander, 2002)
prédite et intervalles de confiance à 95 % (plage ombragée) pour le cultivar de luzerne réduit en lignine (54HVX41 ; y1) et les cultivars de référence (y2) en réponse aux degrés-jours cumulés (cumulative growing degree days). ... 11
Figure 1.6 Modèle de préestomac. Fx, flux d’N au site x ; Fy, x, flux d’N vers le pool y depuis le pool x. ... 29
Figure 1.7 Rendement en protéines vraies (true protein yield) en fonction de l’apport en protéines
métabolisables (MP supply). Concentrations en acides aminés déterminées : * —— * = principalement sur des échantillons regroupés analysés par des analyseurs d’acides aminés ; ● ______ ● = sur des échantillons individuels analysés par dilution isotopique avec spectrométrie de masse par chromatographie en phase gazeuse. ... 42
Figure 1.8 Relation entre l’azote uréique du lait (MUN) et l’excrétion d’azote urinaire (UN). ... 43 Figure 1.9 Effet de l’ajout de Met et de Lys protégées sur le lait corrigé pour la matière grasse (kg/d)
pour des rations ≤15,0 % PB/kg (base MS). Le graphique présente des différences normalisées de moyennes avec un IC à 95 %. Hétérogénéité (Q-statistique, χ2) entre les études = 62,1 (15 dl), P <0,001 (I2 = 75,8 %). Valeurs moyennes (■) proportionnelles à la taille de chaque étude. ... 44
Figure 1.10 Effet de l’ajout de Met et de Lys protégées sur la quantité de protéines produite dans le
lait (kg/d) pour des rations ≤15,0 % PB/kg (base MS). Le graphique présente des différences normalisées de moyennes avec un IC à 95 %. Hétérogénéité (Q-statistique, χ2) entre les études = 63,5 (15 dl), P <0,001 (I2 = 76,4 %). Valeurs moyennes (■) proportionnelles à la taille de chaque étude. ... 44
Figure 1.11 Relation entre l’apport (N intake) et la production (N output) d’azote dans les fèces, le lait,
l’urine et l’excréta. Les symboles représentent les valeurs observées et les lignes pleines sont des prédictions du modèle. ... 49
Liste des abréviations françaises
ADF Fibres insolubles dans un détergent acideADL Lignine brute
CH4 Méthane
CVMS Consommation volontaire de matière sèche
His Histidine
Lys Lysine
Met Méthionine
MS Matière sèche
NDF Fibres insolubles dans un détergent neutre
Liste des abréviations anglaises
AA Amino acids TTNDFD Total tract neutral detergent
BCAA Branched-chain amino acids fiber digestibility
BCS Body condition score
BW Body weight
CP Crude protein
DIM Days in milk
DM Dry matter
DMI Dry matter intake
EAA Essential amino acids ECM Energy corrected milk FCM Fat corrected milk MP Metabolizable protein MUN Milk urea nitrogen
NDFD Neutral detergent fiber digestibility NEAA Non-essential amino acids NEFA Non-esterified fatty acids NEL Net energy for lactation NFC Non-fiber carbohydrates NPN Nonprotein nitrogen
OM Organic matter
RDP Rumen degradable protein
RP Rumen-protected
RUP Rumen undegradable protein SCC Somatic cell count
SD Standard deviation
SEM Standard error of the mean
Remerciements
L’accomplissement d’un projet de recherche et la rédaction d’un mémoire apportent certes du mérite à l’auteur, mais également à tous les gens ayant contribué à cette aventure. L’aide précieuse dont j’ai bénéficié durant ma maîtrise m’a non seulement permis de mener à terme ce projet, mais m’a également fait évoluer en tant qu’agronome. J’aimerais donc prendre le temps de remercier l’équipe de professionnels qui m’a supporté durant ce parcours.
Tout d’abord, j’aimerais exprimer ma sincère reconnaissance à Édith, ma directrice de recherche, qui m’a offert un encadrement exceptionnel. Merci pour ton professionnalisme, tes précieux conseils et ta confiance. Travailler avec toi m’aura permis d’accumuler des expériences professionnelles et personnelles qui me seront très utiles dans ma carrière. Je me considère chanceux d’avoir pu profiter de ton expertise pour développer ce bagage. Merci aussi à Rachel, ma codirectrice, pour son soutien et son efficacité, particulièrement lors de la phase expérimentale du projet. Je me souviendrai longtemps des soirées passées en ta compagnie en train de finaliser le traitement des échantillons ! Ta passion et ton dévouement sont pour moi une source d’inspiration.
Merci également à Yolaine et Nancy pour leur support technique et professionnel lors de mes analyses aux laboratoires de l’Université Laval. Merci à l’équipe d’Agriculture et Agroalimentaire Canda de Québec, Gaëtan, Geneviève et Camille, pour leur précieuse aide lors de la récolte des fourrages et pour leur support au niveau des analyses. Merci aussi à l’équipe d’Agriculture et Agroalimentaire Canada de Sherbrooke, Hélène, Daniel, Danielle et Mario, pour leur accueil chaleureux, leur support professionnel et leur temps. Merci à l’équipe du Centre de recherche en sciences animales de Deschambault, Annie, Emmy et Jonathan, pour leur aide et leur compréhension. Votre bonne humeur a grandement contribué à rendre mes nombreuses visites à l’étable des plus agréables.
J’aimerais aussi souligner l’importante contribution de plusieurs étudiants m’ayant aidé lors de la phase expérimentale. Catherine, Florencia, Jennifer, Marie-Soleil, Sabrina et Sophie, merci pour votre rigueur, votre disponibilité et votre bonne humeur. Votre aide était essentielle à la réussite de ce projet (et au maintien de ma santé mentale) !
Finalement, je ne peux passer outre ma gratitude envers mes proches. J’aimerais d’abord remercier mes parents, Mario et Nathalie, qui m’ont toujours supporté tout au long de mes études. Venir décrocher à la maison les fins de semaine m’a permis de recharger mes batteries, ce qui était parfois plus que nécessaire ! Je voudrais également offrir un remerciement spécial à mon amoureuse. Pénélope, merci pour ton soutien, tes conseils et tes encouragements. T’avoir dans ma vie me rend tout simplement meilleur. Je vous aime !
Avant-propos
Ce mémoire contient un chapitre rédigé sous forme d’article scientifique, pour lequel je suis le principal auteur. Les coauteurs sont les chercheurs R. Gervais, H. Lapierre, D. R. Ouellet, G. F. Tremblay, C. Halde, M.-S. Boucher et E. Charbonneau. Toutes ces personnes se sont impliquées dans les travaux de recherche. L’article portant le titre « Milk production, nutrient digestion, and nitrogen utilization efficiency as affected by metabolizable protein and energy supply in dairy cows fed alfalfa-based diets » sera soumis pour publication dans la revue « Journal of Dairy Science ».
Introduction
La luzerne (Medicago sativa L.) est la légumineuse fourragère la plus cultivée au Canada (Statistique Canada, 2018). Cette légumineuse a un contenu élevé en protéines, un bon rendement, une bonne persistance, la capacité d’améliorer la structure et la fertilité du sol, une bonne résistance à la sécheresse, l’habileté à piéger profondément l’azote incrusté et la capacité de fixer l’azote atmosphérique de façon symbiotique (Émile et al., 1997; Julier et Huyghe, 1997), ce qui a contribué à sa popularité.
Toutefois, malgré une concentration en fibres plus faible, les légumineuses sont généralement moins digestes que les graminées (Hoffman et al., 1998). Cela peut notamment s’expliquer par une digestibilité inférieure des fibres chez cette famille de plante (Hoffman et al., 1998). L’amélioration de la digestibilité des fibres serait bénéfique pour les ruminants (Jung et Allen, 1995) et représenterait une opportunité pour le secteur de la production laitière, tant au niveau économique qu’environnemental (Hatfield et al., 1999). Selon une méta-analyse (Oba et Allen, 1999), l’augmentation d’une unité de pourcentage de la digestibilité des fibres insolubles dans un détergent neutre (NDF), soit in vitro ou in situ, est associée à une augmentation de 0,25 kg par jour de la production de lait corrigé pour la matière grasse (4 %). Plusieurs cultivars de luzerne plus digestible ont récemment fait leur apparition sur le marché canadien. Étant donné l’absence d’études indépendantes testant ces luzernes sous les conditions agroclimatiques du Canada, il serait intéressant de vérifier l’intérêt d’utiliser une luzerne plus digestible cultivée au Canada dans les rations de vaches en lactation.
De plus, malgré sa concentration élevée en protéines brutes, une grande partie (de 31,1 à 61,6 %) des protéines de la luzerne sont formées soit d’azote non protéique, soit de protéines vraies hautement dégradables dans le rumen (NRC, 2001). L’utilisation de ces protéines brutes hautement dégradables par les micro-organismes du rumen pourrait être améliorée par une meilleure adéquation entre l’apport en azote et en énergie disponible dans le rumen. Par conséquent, augmenter la digestibilité des parois cellulaires des légumineuses serait bénéfique pour les ruminants en augmentant la synthèse de protéines microbiennes dans le rumen (Jung et Allen, 1995). De plus, les vaches laitières sont régulièrement alimentées de rations ayant un apport excessif en protéines alors que la production de lait et des composantes peut être maintenue, ou même augmentée, tout en réduisant la concentration en protéines brutes des rations (Broderick, 2018). Réduire la concentration en protéines de la ration pourrait améliorer la rentabilité, en améliorant l’efficacité d’utilisation de l’azote tout en réduisant son excrétion dans l’environnement (Holt et al., 2013). Actuellement, les besoins azotés des vaches laitières sont exprimés en termes de protéines métabolisables plutôt qu’en acides aminés essentiels individuels (NRC, 2001). Avec des rations fournissant un apport adéquat en protéines métabolisables, les vaches sont probablement suralimentées en certains acides aminés, ce qui entraine une faible efficacité d’utilisation de
l’azote (Arriola Apelo et al., 2014). Cependant, une diminution de l’apport en protéines métabolisables est souvent associée à une production laitière réduite, probablement parce que les besoins en certains acides aminés essentiels ne sont pas satisfaits (Arriola Apelo et al., 2014). Il serait donc intéressant de tenter de maximiser l’utilisation protéique de la luzerne par une diminution de l’apport en protéines métabolisables de la ration tout en comblant les besoins en acides aminés.
Le premier chapitre de ce mémoire présente l’état des connaissances actuelles nécessaires pour améliorer l’utilisation énergétique et protéique de la luzerne. Le deuxième chapitre sera formé de deux parties. Dans un premier temps, l’évaluation de la performance de vaches laitières alimentées d’un cultivar de luzerne sélectionné pour une faible teneur en lignine sera présentée. Ensuite, l’évaluation de l’effet des apports en protéines métabolisables et en énergie sur la production laitière, la digestion des nutriments et l’efficacité d’utilisation de l’azote des vaches laitières sera exposée. Les objectifs de ce chapitre sont de vérifier l’intérêt d’utiliser un cultivar de luzerne plus digestible sur les fermes laitières québécoises, et de vérifier s’il est possible de diminuer l’apport en protéines métabolisables sans affecter négativement les performances zootechniques des vaches laitières.
Chapitre 1 Revue des travaux antérieurs
1.1
Amélioration de l’utilisation énergétique de la luzerne par l’augmentation
de sa digestibilité
Les glucides représentent la principale source d’énergie des ruminants. Ils peuvent être divisés en deux catégories : les glucides non structuraux et les glucides structuraux. Les glucides non structuraux sont plus facilement digestibles. Les glucides structuraux apportent tout de même une part importante d’énergie à l’animal grâce à la fermentation ruminale, mais cela varie en fonction de leur digestibilité. La luzerne a un contenu élevé en glucides structuraux (Émile et al., 1997), ce qui explique sa faible valeur énergétique (Julier et Huyghe, 1997). Plusieurs moyens ont été utilisés pour accroître la digestibilité de la luzerne, et quelques études ont évalué la performance de cultivars plus digestibles semés en parcelles. Toutefois, très peu d’études ont testé ces cultivars dans la ration de ruminants.
1.1.1 Digestibilité des fourrages
Les fourrages sont définis comme étant des aliments composés de tiges, de feuilles et possiblement de grains, qui sont servis frais, en foin ou en ensilage (NRC, 2001). Ils représentent la plus importante ressource alimentaire en nutrition des ruminants, puisqu’ils sont leur première source de nutriments (Jung et Allen, 1995). Même s’il est possible d’élever des ruminants avec des rations contenant relativement peu de fourrages, il y a plusieurs raisons de maintenir les fourrages à de hauts niveaux dans les rations (Jung et Allen, 1995). En effet, les fonctions ruminales et la santé de l’animal sont améliorées avec des régimes à base de fourrages (Jung et Allen, 1995). De plus, leurs coûts de production sont généralement inférieurs à ceux des cultures céréalières, et les cultures fourragères vivaces sont considérées comme étant plus respectueuses de l’environnement, entre autres grâce à la protection de l’érosion des sols et la réduction de la pollution de l’eau par les pesticides et les engrais, utilisés en plus petites quantités dans ces cultures (Jung et Allen, 1995). Toutefois, les fourrages ont souvent une faible digestibilité et une haute teneur en parois cellulaires, ce qui limite l’énergie disponible pour les animaux consommant une ration contenant beaucoup de fourrages (Jung et Allen, 1995). L’amélioration génétique pour la production et la croissance des vaches laitières va d’ailleurs impliquer une réduction de la capacité des fourrages à fournir une part importante d’énergie de la ration, à moins que la disponibilité de l’énergie des fourrages soit augmentée (Jung et Allen, 1995).
Les fourrages sont constitués de divers nutriments : protéines, lipides, glucides non structuraux, glucides structuraux, vitamines et minéraux. La plupart des nutriments de la ration qui ne sont pas des fibres sont pratiquement tous digérés ou fermentés durant le passage dans le système digestif des animaux (Allen et Mertens, 1988). Les fibres, quant à elles, représentent une partie significative de la diète des herbivores, dont
la productivité est limitée par leur capacité à les digérer (Allen et Mertens, 1988). Elles représentent de 30 à 80 % de la matière sèche (MS) des fourrages, et constituent une source majeure d’énergie pour les ruminants via les acides gras volatils produits par les micro-organismes du rumen (Hatfield et al., 1999).
Les fibres sont un facteur important pour déterminer la qualité d’un fourrage (Hatfield et al., 1999). Elles sont constituées de polysaccharides structuraux ayant des compositions et des structures variées, d’acides hydroxycinnamiques, de lignine, de protéines (métaboliques et structurelles), d’ions et d’eau (Hatfield et al., 1999). L’interaction de ces composants contrôle l’organisation des parois cellulaires et leur intégrité structurelle, mais pourrait aussi avoir un effet sur leur dégradation par les herbivores (Hatfield et al., 1999). Étant donné que les fibres sont résistantes à la digestion par les enzymes produites par les ruminants, les contraintes affectant la digestion des fibres par les microbes du rumen sont des facteurs critiques qui limitent la productivité des ruminants (Allen et Mertens, 1988). La fermentation de ces fibres est effectuée dans un écosystème complexe qui est influencé par les interactions dynamiques de la ration, de la population microbienne et de l’animal (Allen et Mertens, 1988). Selon Nocek et Russell (1988), la digestibilité ruminale de la NDF des fourrages peut varier de 18,9 à 77,1 %.
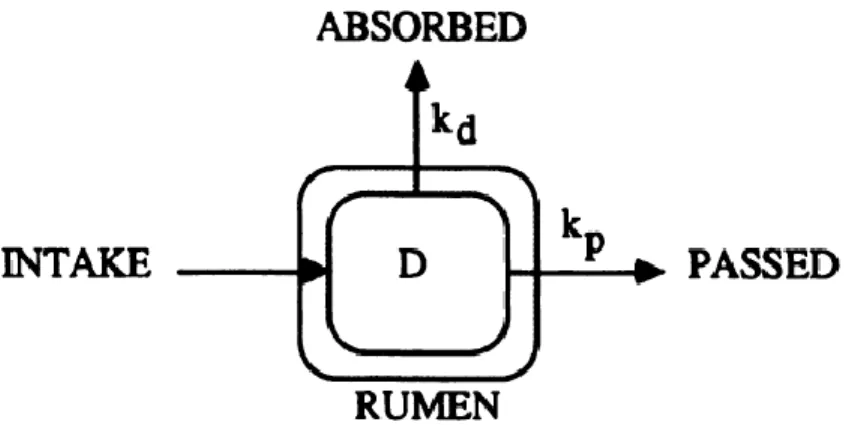
La digestibilité est définie comme étant le ratio entre l’absorption et la consommation de nutriments. Selon Allen et Mertens (1988), la digestibilité des fibres est fonction de la proportion de fibres qui est potentiellement digestible, mais également du taux de digestion et du taux de passage des fibres dans le rumen (Figure 1.1). N’importe quel facteur qui influence à la hausse la fraction indigeste des fibres peut devenir une contrainte à la digestion de celles-ci dans le rumen (Allen et Mertens, 1988). D’autres facteurs ayant un effet négatif sur le taux de digestion ou le temps de rétention des fibres potentiellement digestibles peuvent également devenir une contrainte (Allen et Mertens, 1988).
Tiré de Allen et Mertens (1988).
Figure 1.1 Représentation graphique d’un modèle cinétique simple de la disparition des fibres dans le
rumen : Les fibres potentiellement digestibles sont représentées par D, le taux fractionnaire de digestion par K et le taux fractionnaire de passage par K.
La plus grande contrainte indépendante à la digestion reste la fraction des fibres qui est indigeste, celle-ci représentant jusqu’à un tiers et la moitié de la fraction des fibres totales des graminées et des légumineuses, respectivement (Allen et Mertens, 1988). La contrainte du taux de digestion va dépendre des caractéristiques intrinsèques de la matrice chimique des parois des cellules végétales, des facteurs microbiens qui modifient le taux maximal de digestion et du temps de rétention des fibres dans le rumen (Allen et Mertens, 1988). Pour ce qui est du taux de passage, plusieurs facteurs entrent en jeu comme la rétention sélective et le tapis ruminal. La rétention sélective dans le rumen permet une fermentation plus extensive des fibres potentiellement digestibles en diminuant le taux de passage (Allen et Mertens, 1988). En effet, selon Lund et al. (2007), les particules alimentaires contenant plus de NDF digestibles ont un temps de rétention plus long, ce qui serait attribuable aux différences dans la distribution de ces fibres plus digestibles dans la plante, la taille des particules ou la flottabilité. Pour ce qui est du tapis ruminal fibreux, la formation de celui-ci permet de retenir les particules de fibres qui pourraient potentiellement quitter le rumen, ce qui augmente le temps de digestion de ces particules (Allen et Mertens, 1988).
La digestibilité des fibres potentiellement digestibles varie de 40 à 86 % en utilisant différents taux de digestion (0,04-0,12/h) et taux de passage (0,02-0,06/h) observés chez les animaux (Allen et Mertens, 1988). N’importe quel facteur ayant pour effet d’augmenter le taux de digestion ou le temps de rétention peut donc avoir un effet fortement positif sur la digestibilité (Allen et Mertens, 1988). L’apport en nutriments est fonction à la fois de la consommation et de la digestibilité de ces nutriments (Allen et Mertens, 1988). Même si l’apport en nutriments et la digestibilité sont positivement liés au taux de digestion, ils ont des relations opposées avec le temps de rétention dans le rumen ; une augmentation de la consommation est associée avec un taux de passage accéléré, ce qui diminue la digestibilité des fibres (Allen et Mertens, 1988).
La digestibilité est hautement corrélée avec certains composants de la paroi cellulaire qui peuvent être analysés de façon chimique, soit les fibres insolubles dans un détergent acide (ADF), la lignine brute (ADL) et la NDF (Allinson et al., 1969; Hill et Barnes, 1977; Buxton et al., 1987; Julier et al., 1996). Toutefois, aucune mesure chimique actuelle n’est reliée au taux de digestion des fibres (Allen, 1996b). La digestibilité de l’aliment entier est influencée par la fraction entièrement digestible des constituants alimentaires non fibreux (Allen et Mertens, 1988). La grande variation de cette fraction dans les aliments pour ruminant est responsable de la mauvaise relation entre la digestibilité des fibres et de la MS (Allen et Mertens, 1988).
1.1.1.1 Analyses chimiques corrélées à la digestibilité
La NDF et l’ADF font partie des mesures les plus courantes pour les fibres dans les analyses alimentaires de routine (NRC, 2001). La NDF est principalement composée d’hémicelluloses, de cellulose et de lignine. Les proportions de ces constituants affectent la digestibilité de la NDF, ce qui veut dire que plusieurs aliments ou
rations ayant le même niveau de NDF n’auront pas nécessairement un niveau d’énergie similaire (NRC, 2001). La NDF est utilisée pour la formulation des rations des vaches en lactation parce que la concentration en NDF est reliée positivement avec le remplissage du rumen et négativement avec la densité énergétique de l’aliment (Mertens, 1987). La NDF représente le meilleur prédicteur chimique unique de la consommation volontaire de matière sèche (CVMS) (Allen, 1996a). Pour ce qui est de l’ADF, elle est principalement composée de cellulose et de lignine. La NDF représente la meilleure expression des fibres actuellement disponible, mais il existe aussi des recommandations pour l’ADF (NRC, 2001).
La lignine, liée à d’autres composants de la paroi, a un rôle pivot dans la dégradation des parois cellulaires (Hatfield et al., 1999). La lignine est indigeste, et se lie avec la cellulose et l’hémicellulose, ce qui diminue également la digestibilité de ces composants de la paroi cellulaire (McCaslin et al., 2014). Les tissus de plante hautement lignifiés traversent le système digestif de l’animal et ne sont pas utilisés pour la croissance et le développement de celui-ci (Sulc et al., 2016). La lignine limite donc la digestibilité, le potentiel d’ingestion et la disponibilité de l’énergie, ce qui finit par limiter les performances et la production du ruminant (Sulc et al., 2016).
1.1.1.2 Analyses de digestibilité
La digestibilité est une mesure courante de la qualité fourragère qui estime le contenu énergétique du fourrage (Allen, 1996b). Cette mesure est typiquement prédite à partir du contenu en fibres (Allen, 1996b). Plusieurs équations ont été développées pour prédire la digestibilité à partir des différentes teneurs en fibres, ces dernières étant moins digestibles que les composants non fibreux (Allen, 1996b). Par exemple, il est possible de prédire la digestibilité de la NDF avec les teneurs en NDF, en protéines liées à la fibre NDF et en ADL. Toutefois, Robinson et al. (2004) ont observé une faible relation entre la digestibilité in vitro mesurée avec une incubation de 48 h et cette méthode. En effet, puisque la digestibilité des fibres est fonction de la fraction potentiellement digestible, du taux de digestion et du temps de rétention dans le rumen, elle est mal prédite par les mesures chimiques (Allen, 1996b). Une autre façon de déterminer la digestibilité des fibres est de la mesurer à l’aide d’incubations dans le liquide ruminal.
Il existe trois façons de mesurer la digestibilité des fibres NDF par le liquide ruminal, soit la méthode in vivo, in
situ (ou in sacco) et in vitro. Dans chacune de ces méthodes, la quantité de fibres restante après le processus
de digestion est comparée à la quantité initiale. La méthode in vivo est la plus complexe, car elle nécessite des vaches canulées au duodénum ou au rumen et requiert soit le prélèvement de digesta qui passe à l’omasum ou une technique d’évacuation ruminale qui consiste à retirer complètement le contenu du rumen à quelques reprises (Huhtanen et al., 2010). La méthode in situ est une des deux méthodes les plus couramment utilisées dans l’industrie, notamment en recherche (Berthiaume et al., 2015). Elle consiste à placer des échantillons de fourrages dans des sachets de tissus qui seront insérés dans le rumen par une canule (Ørskov et McDonald,
1979). Pour ce qui est de la méthode in vitro, celle-ci est largement utilisée dans l’industrie puisqu’elle est moins couteuse et nécessite moins de main-d’œuvre. Elle consiste à placer des échantillons de fourrages dans une solution composée de salive artificielle (substances tampons et minéraux) et de liquide ruminal à température corporelle (38,9 °C) en absence d’air pour un temps donné (Goering et Van Soest, 1970). Bien qu’il soit suggéré d’incuber les échantillons durant 48 heures (NRC, 2001), plusieurs recommandent de réduire ce temps à 30 ou 24 heures afin de mieux refléter la digestibilité potentielle dans le rumen d’une vache forte productrice (Berthiaume et al., 2015). Des équations de calibration ont été développées pour prédire la digestibilité in vitro de la NDF à partir de la spectroscopie de réflectance dans le proche infrarouge, ce qui rend cette analyse beaucoup moins couteuse et plus rapide. Il existe toutefois une grande variabilité intra et inter laboratoire pour la NDF digestible in vitro, à cause de l’absence de standards pour ce dosage (Hall et Mertens, 2012).
Selon Allen (2011), les taux de digestion déterminés in vitro surestiment la digestibilité in vivo des fibres et sont seulement utiles pour comparer des différences relatives de digestibilité entre les fourrages. Lopes et al. (2015) ont utilisé un modèle qui prédisait de façon in vitro la digestibilité des fibres du tractus total in vivo (total tract
neutral detergent fiber digestibility : TTNDFD) avec la proportion de la NDF totale potentiellement digestible
(pdNDF), le taux de dégradation de la NDF potentiellement digestible, et le taux de passage de celle-ci. Le modèle a réussi à bien prédire la TTNDFD de façon similaire aux mesures in vivo, la corrélation étant significative (R2 = 0,68). La relation entre la digestibilité in vitro 30-h de la NDF et la TTNDFD n’était pas significative, alors que la méthode in vitro 48-h était corrélée avec la digestibilité des fibres du tractus total in vivo, mais de façon moindre que le modèle (R2 = 0,30). Toujours selon Lopes et al. (2015), ce modèle pourrait améliorer notre habileté à optimiser l’utilisation des fourrages et la production laitière.
Malgré les nombreuses méthodes d’analyse de la digestibilité, il est important de reconnaître que peu importe la méthode, ces analyses décrivent uniquement une composante du système se traduisant par la consommation et la digestion des aliments par l’animal (Mertens, 1987). Il faut savoir que la consommation et la digestibilité dépendent de l’interaction entre la ration, l’animal et son environnement d’alimentation (Mertens, 1987).
1.1.2 Façons d’accroître la digestibilité de la luzerne par la sélection génétique
Plusieurs facteurs influencent la digestibilité des fourrages. Il existe bien sûr une variation de la digestibilité entre les différentes espèces de fourrages, les graminées ayant en général des fibres plus digestibles que les légumineuses (Messman et al., 1992; Hoffman et al., 1998), mais aussi au sein d’une même espèce. Chez une espèce fourragère en particulier, c’est le stade de maturité qui est le facteur le plus déterminant quant à la digestibilité des fibres (Llamas-Lamas et Combs, 1990). Par exemple, pour la luzerne, la digestibilité des fibres
diminue avec la maturité (Shaver et al., 1988; Llamas-Lamas et Combs, 1990). Cela peut s’expliquer par le fait que la digestibilité des tiges et le ratio feuilles/tiges diminuent lors de la croissance (Julier et Huyghe, 1997).
Le stress hydrique et la température peuvent aussi avoir un effet sur la digestibilité des fibres ou de la MS. En conditions contrôlées, des luzernes poussant avec un fort stress hydrique ou avec des températures plus fraiches ont une meilleure digestibilité totale de la MS, moins d’ADF et moins d’ADL (Vough et Marten, 1971). Les mêmes constats ont été observés en parcelles pour la température alors que des luzernes ayant poussé sur un site plus nordique avaient une digestibilité des fibres généralement supérieure à celle ayant poussé sur des sites plus au sud, et ce pour un même stade de développement (Lauzon et al., 2019). D’autres facteurs comme le cycle de repousse, le lieu ou la méthode de conservation influencent la digestibilité. En effet, Julier et Huyghe (1997) ont montré que la MS de la luzerne était en moyenne plus digestible à la première coupe qu’à la deuxième. Les mêmes auteurs ont aussi observé que le lieu (sans être associé à la température moyenne) influence la digestibilité totale de la luzerne (Julier et Huyghe, 1997). Cet effet serait potentiellement lié à la composition minérale du sol, qui pourrait affecter le contenu en cendres de la luzerne (Julier et Huyghe, 1997). Finalement, selon la méthode de conservation, de 88 à 94 % de la digestibilité potentielle de la matière organique des fourrages est conservée (Waldo et Jorgensen, 1981).
Une autre façon de faire varier la digestibilité des fourrages est par l’amélioration génétique. Chez les graminées, la concentration en parois cellulaires est élevée, mais une fraction importante de celle-ci est digestible. Une réduction de la concentration en paroi cellulaire serait donc plus efficace qu’améliorer la digestibilité pour cette famille de plante (Jung et Allen, 1995). C’est le contraire chez les légumineuses, celles-ci bénéficieraient donc plus d’une augmentation de la digestibilité de leurs parois cellulaires (Jung et Allen, 1995). Que ce soit par la sélection conventionnelle ou par l’approche transgénique, les techniques d’amélioration génétique peuvent être envisagées pour améliorer la digestibilité de la luzerne.
1.1.2.1 Approche conventionnelle
L’amélioration de la génétique par des croisements dans une espèce fourragère largement utilisée représente la façon la moins couteuse d’améliorer la valeur énergétique d’un fourrage (Émile et al., 1997). Il existe une variation génétique de la digestibilité chez la luzerne (Allinson et al., 1969; Heinrichs et al., 1969; Buxton et al., 1987; Julier et al., 1996) et il est possible d’améliorer les populations par la sélection génétique tant pour le rendement que pour la valeur nutritive (Demment et al., 1986). En effet, la corrélation négative entre le rendement et la qualité de la luzerne (Demment et al., 1986; Julier et al., 1996) serait suffisamment faible pour permettre un progrès (Shenk et Elliott, 1971). De plus, certains auteurs ne rapportent pas de corrélations significatives (Allinson et al., 1969; Hill et Barnes, 1977; Julier et Huyghe, 1997), ce qui indique qu’il est possible d’identifier des cultivars à la fois productifs et plus digestibles.
Plusieurs critères de sélection ont été identifiés comme ayant le potentiel d’accroitre la digestibilité de la luzerne, par exemple le ratio feuilles/tiges (Émile et al., 1997), la digestibilité de la tige (Terry et Tilley, 1964), la teneur en pectines (Tecle et al., 2006) ou la lignine des parois cellulaires (Buxton et al., 1987; Julier et al., 1996). Les sélectionneurs d’Alforex Seeds ont d’ailleurs utilisé la sélection classique pour réduire la lignine de la luzerne, ce qui a mené à la commercialisation du cultivar de luzerne Hi-Gest® (Sulc et al., 2016).
1.1.2.2 Approche transgénique
L’approche la plus expéditive pour améliorer la digestibilité des fourrages serait probablement une combinaison des approches moléculaires avec la sélection génétique traditionnelle (Hatfield et al., 1999). Le composé indigeste par les ruminants le plus important dans les tiges de luzerne est la lignine, qui est localisée dans les parois cellulaires (Albrecht et al., 1987). Ainsi, une augmentation de la valeur nutritive de la tige semble être une approche à favoriser pour améliorer significativement la qualité nutritive de la luzerne (Sulc et al., 2016). Plusieurs étapes sont nécessaires pour synthétiser la lignine chez les plantes, ces étapes impliquent douze enzymes différentes (Figure 1.2 ; Lee et al., 2011b). Il existe plusieurs façons de réduire le contenu en lignine de la luzerne par le génie génétique. Il est en effet possible de réduire à la baisse certaines enzymes comme celles du cytochrome P450 (C4H, C3H et F5H ; Reddy et al., 2005), l’HCT (Shadle et al., 2007), la COMT ou la CCoAOMT (Guo et al., 2001).
Tiré de Lee et al. (2011b).
Plusieurs types d’inhibition de l’expression des gènes ont été testées afin de réduire le contenu en lignine et d’augmenter la digestibilité de la NDF, mais une seule a réussi à le faire sans affecter simultanément des traits agronomiques clés comme le rendement, la résistance multiple aux ravageurs, la persistance ou la verse (McCaslin et al., 2014). La méthode ayant fonctionné est la modification génétique utilisant l’interférence d’ARN pour réguler à la baisse la CCoAOMT (McCaslin et al., 2014). En octobre 2014, l’agence canadienne d’inspection des aliments a déréglementé la modification génétique de la luzerne réduite en lignine au Canada (McCaslin et al., 2014), ce qui a permis son entrée sur le marché.
1.1.3 Intérêts agronomiques d’une luzerne plus digestible
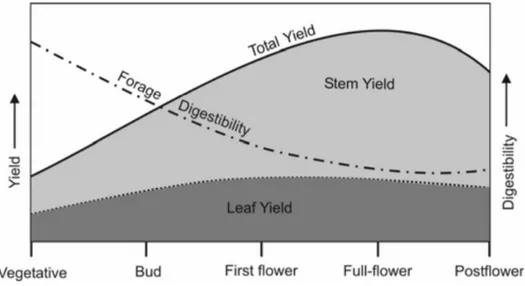
L’objectif lorsqu’on cultive de la luzerne, c’est de maximiser le rendement et la qualité fourragère sans compromettre la persistance (Orloff et Putnam, 2004). Toutefois, le rendement et la qualité sont inversement reliés (Blank et al., 2001). En effet, lorsque la luzerne gagne en maturité, le contenu en NDF augmente tandis que la digestibilité de la NDF diminue, ce qui peut être attribuable à la diminution du ratio feuilles/tiges et à l’augmentation conséquente du contenu en lignine (Figure 1.3 ; Orloff et Putnam, 2004).
Tiré de Orloff et Putnam (2004).
Figure 1.3 Qualité et rendement relatif de la luzerne à différents stades de croissance.
La réduction du contenu en lignine de la luzerne devrait permettre une période prolongée de récolte lorsque le fourrage a une valeur nutritive acceptable (Sulc et al., 2016), ce qui permettrait d’ajuster plus facilement le moment de récolte en fonction des aléas météorologiques. Cela pourrait aussi permettre d’avoir des rendements plus élevés en récoltant la luzerne plus tard (Sulc et al., 2016). Par exemple, Grev et al. (2017) ont démontré qu’avec un délai de coupe de 5 jours, un cultivar de luzerne à teneur réduite en lignine a permis d’avoir des rendements supérieurs de 21 % avec une réduction de seulement 3 % de la qualité fourragère relative. Cette
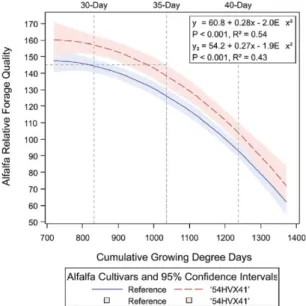
amélioration est possible grâce au fait que le rendement des cultivars de références et de la luzerne réduite en lignine est similaire (Figure 1.4), et que la qualité fourragère relative est plus élevée chez la luzerne réduite en lignine entre 772 et 1248 degrés-jours cumulés (Figure 1.5 ; Grev et al., 2017). De plus, une coupe plus tardive permet non seulement d’améliorer le rendement, mais aussi la persistance (Blank et al., 2001).
Tiré de Grev et al. (2017).
Figure 1.4 Masse de MS (alfalfa forage mass DM) prédite et intervalles de confiance à 95 % (plage ombragée) pour le cultivar de luzerne réduit en lignine (54HVX41 ; y1) et les cultivars de référence (y2) en réponse aux degrés-jours cumulés (cumulative growing degree days).
Tiré de Grev et al. (2017).
Figure 1.5 Qualité fourragère relative (alfalfa relative forage quality; Moore et Undersander, 2002) prédite et intervalles de confiance à 95 % (plage ombragée) pour le cultivar de luzerne réduit en lignine (54HVX41 ; y1) et les cultivars de référence (y2) en réponse aux degrés-jours cumulés (cumulative growing degree days).
Cette amélioration de la flexibilité de récolte facilite aussi la gestion des mélanges fourragers, puisque le producteur pourrait baser le moment de la récolte sur le stade de développement des autres espèces du mélange sans trop affecter la qualité de la luzerne.
Malgré ces avantages, les luzernes plus digestibles transgéniques font l’objet de controverse à trois niveaux. Tout d’abord, puisque ces cultivars ont aussi le gène de résistance au glyphosate, ils pourraient favoriser l’utilisation de cet herbicide qui est déjà souvent utilisé dans les rotations, notamment pour le maïs et le soya. Une absence de diversité d’herbicides causée par une rotation de plantes résistantes au glyphosate peut contribuer à l’apparition de mauvaises herbes résistantes, un problème en croissance (Bonny, 2016). Le gène de résistance au glyphosate pourrait aussi favoriser l’établissement de prairies de luzerne pures, puisque les autres plantes fourragères pérennes n’ont pas ce gène de résistance. L’utilisation de mélanges fourragers à base de luzerne, une pratique courante au Canada, a pourtant des avantages non négligeables entre autres au niveau du rendement (Bélanger et al., 2014), de la lutte aux mauvaises herbes (Sanderson et al., 2012; Bélanger et al., 2014), de la survie à l’hiver (Smith, 1962), du cycle de l’azote (Nyfeler et al., 2011) et de la valeur nutritive (Bélanger et al., 2014). Finalement, la commercialisation des luzernes modifiées génétiquement inquiète plusieurs acteurs de l’industrie canadienne. En effet, une contamination physique, une pollinisation croisée ou des graines dures germant après plusieurs années pourraient faire apparaître ces luzernes à des endroits imprévus, ce qui préoccupe grandement l’industrie des semences puisque l’Union européenne ne tolère pas les semences génétiquement modifiées (Yungblut et Jalbert, 2012). L’industrie des produits biologiques est également préoccupée par cette situation, étant donné que celle-ci ne permet pas l’utilisation de plantes génétiquement modifiées (Yungblut et Jalbert, 2012). Finalement, l’industrie du miel s’inquiète d’une contamination potentielle par du pollen provenant de plantes génétiquement modifiées (Yungblut et Jalbert, 2012).
1.1.4 Caractéristiques des luzernes plus digestibles
La digestibilité de la NDF de la luzerne peut varier de 30,9 % à 62,5 % (Nocek et Russell, 1988). Les luzernes plus digestibles pourraient permettre d’avoir une meilleure qualité fourragère pour un même rendement. Elles pourraient aussi avoir un avantage au niveau de l’ensilage. En effet, il a été proposé qu’un fourrage contenant plus de fibres digestibles avant l’ensilage pourrait perdre plus de fibres durant ce processus, ce qui pourrait favoriser la CVMS des animaux (Morrison, 1988; Dado et Allen, 1996).
Un essai réalisé en 2015 dans six états américains (Kansas, Michigan, Ohio, Pennsylvanie, Californie, Wisconsin) visait à évaluer le rendement et la valeur nutritive de plusieurs cultivars de luzerne (Sulc et al., 2016). Trois cultivars de luzerne ont été semés : HarvXtra-008 modifié génétiquement pour avoir moins de lignine,
54R02 ayant de hauts rendements, et WL 355R sélectionné pour une haute qualité nutritive. Comparativement aux cultivars de luzerne conventionnels, il a été démontré qu’à travers les six états durant l’année d’implantation, la luzerne HarvXtra-008 avait de façon constante une valeur d’ADL et de NDF inférieure, ainsi que des valeurs supérieures pour la digestibilité de la NDF (NDFD), la qualité fourragère relative et la teneur en protéines brutes (Tableau 1.1). La valeur nutritive a diminué pour toutes les variétés durant la repousse et les différences entre les variétés sont restées relativement constantes dans le temps.
Tableau 1.1 Valeur nutritive moyenne de trois cultivars de luzerne pendant six dates d’échantillonnage
durant chacun des deux cycles de repousse de l’année d’implantation (2015) à travers six états.
Valeur nutritive HarvXtra-008 WL 355 RR 54R02
ADL (% MS) 4,0b 4,9a 5,0a
NDFD (% NDF) 55,5a 51,0b 50,1b
NDF (% MS) 26,7c 28,7b 30,5a
QFR1 297a 262b 243c
PB2 (% MS) 26,4a 25,8b 25,0c
a-cSur une même ligne, les moyennes sans lettres semblables sont significativement différentes à P = 0,05. 1Qualité fourragère relative.
2Protéines brutes.
Adapté de Sulc et al. (2016).
Le cultivar Hi-Gest 360 (seulement inclus dans les essais en Californie et Pennsylvanie), qui a été sélectionné de façon conventionnelle pour avoir moins de lignine n’était pas significativement différent du cultivar de luzerne sélectionnée pour une haute qualité nutritive (Sulc et al., 2016). Toutefois, ce cultivar a eu une NDFD plus élevée, une valeur fourragère relative plus élevée et une teneur en NDF plus faible que le cultivar de luzerne à haut rendement (Tableau 1.2 ; Sulc et al., 2016). Encore une fois, les différences entre les variétés sont restées relativement constantes dans le temps (Sulc et al., 2016).
Tableau 1.2 Valeur nutritive moyenne de quatre cultivars de luzerne pendant six dates d’échantillonnage
durant chacun des deux cycles de repousse de l’année d’implantation (2015) à travers deux états.
Valeur nutritive
HarvXtra-008 Hi-Gest 360 WL 355 RR 54R02ADL (% MS) 4,2b 4,8a 5,0a 5,1a
NDFD (% NDF) 56,2a 52,5a 51,5a 50,7b
NDF (% MS) 27,3c 28,3bc 29,1b 30,7a
QFR1 284a 265ab 254bc 237c
PB2 (% MS) 26,2a 26,0a 25,8a 24,9a
a-cSur une même ligne, les moyennes sans lettres semblables sont significativement différentes à P = 0,05. 1Qualité fourragère relative.
2Protéines brutes.
Adapté de Sulc et al. (2016).
Grev et al. (2017) ont réalisé au Minnesota entre 2015 et 2016 une étude comparant un cultivar modifié génétiquement pour avoir une teneur réduite en lignine (54HVX41) et plusieurs autres cultivars conventionnels.
Le cultivar faible en lignine avait de plus faibles rendements et une maturité moindre que certains cultivars à certains sites durant les deux années (Tableau 1.3). Pour ce qui est de la protéine et de la NDF, ces valeurs pouvaient respectivement être plus élevées et plus faibles chez le cultivar transgénique dépendamment du site et du cultivar de comparaison durant les deux années. L’ADL était plus faible, et ce indépendamment du cultivar de comparaison, du site et de l’année. Pour ce qui est de la digestibilité de la NDF, celle-ci était plus élevée que les autres cultivars dans tous les sites sauf Rosemount.
Tableau 1.3 Différentes mesures sur différents cultivars de luzerne cultivés à Becker (BE), Rochester (ROC), Rosemount (ROS) et Saint-Paul (SP) au Minnesota en 2015 et en 2016.
2015 2016
Traitement BE ROC ROS SP BE ROS SP
Index de maturité 54HVX41 2,5b 1,7b 1,9 2,6b 4,0 3,5b 4,6 54R02 2,9a 2,1a 2,1 2,8ab 4,0 3,6ab 4,5 DKA43-22RR 2,7ab 2,1a 2,1 3,0a 4,0 3,9a 5,0 WL 355. RR 2,9a 2,2a 2,1 2,8ab 4,0 3,7ab 4,9 Rendement accumulé, Mg ha-1 54HVX41 7,0b 5,8b 6,9b 8,6 15,0b 16,2 20,1 54R02 7,2b 6,8a 7,2ab 8,7 15,9a 16,8 20,5 DKA43-22RR 7,6ab 6,9a 7,4a 9,3 15,9a 16,5 20,8 WL 355. RR 7,9a 6,6a 7,3ab 8,8 15,9a 16,5 20,6 Protéines brutes, g kg-1 54HVX41 230a 218 226 234 187a 186 182 54R02 220b 211 221 226 180ab 182 178 DKA43-22RR 224b 212 224 228 179ab 186 175 WL 355. RR 225ab 215 227 230 178b 184 178 NDF, g kg-1 54HVX41 387c 388 351 351 441b 403 416 54R02 408a 403 359 364 456ab 414 424 DKA43-22RR 401ab 399 357 363 458a 405 430 WL 355.RR 396bc 389 346 356 452ab 407 419 ADL, g kg-1 54HVX41 74c 74b 67b 65b 81b 79b 77b 54R02 86a 83a 73a 74a 87a 85a 81a DKA43-22RR 84ab 83a 71a 73a 88a 84a 83a WL 355.RR 83b 81a 71a 73a 88a 85a 82a NDFD, g kg-1 54HVX41 443a 391a 447 453a 333a 333 339a 54R02 397b 353b 429 419b 288b 300 306b DKA43-22RR 404b 359b 428 418b 282b 305 305b WL 355. RR 402b 365b 436 419b 278b 301 306b
a-cSur une même colonne et une même section, les moyennes sans lettres semblables sont significativement différentes (P ≤ 0,05).
Adapté de Grev et al. (2017).
On peut donc constater que la digestibilité d’une luzerne ayant une teneur réduite en lignine n’est pas nécessairement plus élevée que les autres cultivars à tous les sites (Grev et al., 2017).
Dans une étude réalisée par Émile et Traineau (1993) en France, un cultivar expérimental (6328P) sélectionné pour avoir une digestibilité élevée a été comparé à un cultivar témoin (Tableau 1.4). Le cultivar sélectionné était plus digestible, tant de façon in vitro qu’in vivo, et permettait une meilleure ingestion lors d’un essai animal avec
des moutons (Émile et Traineau, 1993). Comparativement au cultivar témoin, le cultivar expérimental était aussi plus pauvre en cellulose brute et la digestibilité de cette cellulose était plus élevée (Émile et Traineau, 1993). De plus, celui-ci contenait davantage de matière azotée totale (Émile et Traineau, 1993). Il a également été noté que le cultivar à l’étude avait un port moins dressé, un rendement plus faible de 10 % et une plus grande proportion de feuilles à stade égal (Émile et Traineau, 1993).
Tableau 1.4 Caractéristiques comparées de la luzerne Europe (témoin) et la luzerne expérimentale 6328P,
n=66.
Témoin (Europe) Expérimental (6328P) Signifiance2
MS (%) 21,1 20,3 NS
Matière azotée totale (%) 20,1 21,1 **
Cellulose brute (%) 30,1 27,4 **
Digestibilité in vitro1 71,3 75,1 **
Ingestion (g/kg poids mort) 73,4 77,1 **
CUD3 matière organique 65,2 67,9 **
CUD cellulose brute 51,6 54,4 *
1n = 34.
2NS : Non significatif ; * : P ˂ 0,05 ; ** : P ˂ 0,01. 3Coefficient d’utilisation digestive.
Adapté de Émile et Traineau (1993).
1.1.5 Effets de la digestibilité des fourrages sur la vache en lactation
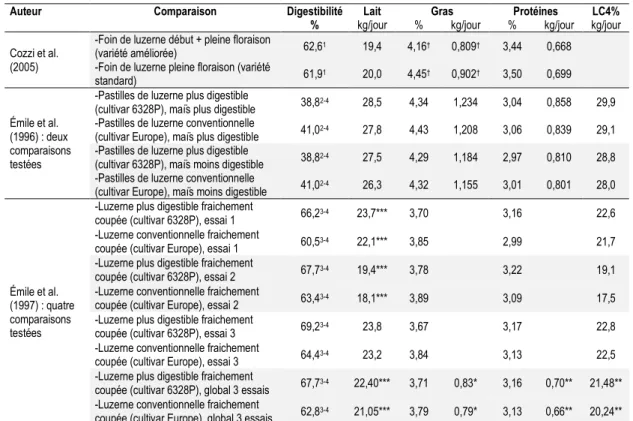
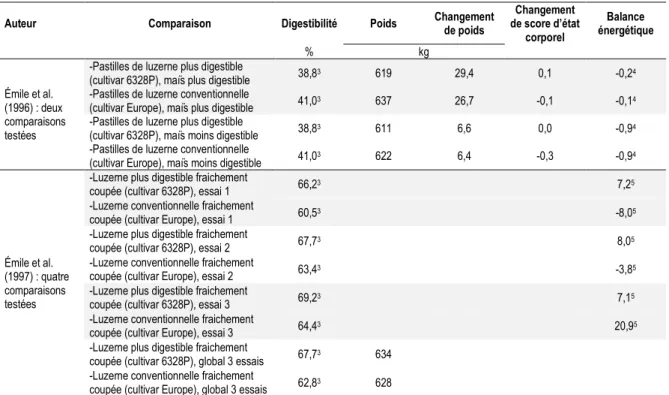
Les fourrages constituent de 35 à 100 % de la ration des vaches en lactation (Messman et al., 1992). La qualité fourragère peut donc avoir une grande influence sur la production laitière (Messman et al., 1992). Puisque la digestibilité des fibres semble être un facteur important influençant la CVMS et la production laitière (Ruiz et al., 1995), plusieurs expérimentations animales ont tenté de mesurer l’effet d’un fourrage plus digestible sur des vaches en lactation. Afin de simplifier l’analyse, seulement les expériences comparant des luzernes (Shaver et al., 1988; Llamas-Lamas et Combs, 1990; Dado et Allen, 1996) ou des cultivars de luzerne améliorés pour être plus digestibles (Émile et al., 1996, 1997; Cozzi et al., 2005) seront présentées.
Il est extrêmement difficile de mener des essais dans lesquels les fourrages ne varient que par un facteur d’intérêt relatif à la qualité, ce qui fait en sorte que presque tous les essais dans la littérature sont confondus quand on essaie d’évaluer l’importance de facteurs spécifiques (Allen, 1996b). En effet, les études comparant des fourrages variant en maturité sont difficiles à comparer en ce qui a trait à l’effet de la digestibilité, car la maturité affecte plusieurs autres facteurs importants en lien avec la qualité (Allen, 1996b). Aussi, selon Oba et Allen (1999), bien que plusieurs expériences aient amassé des données sur la digestibilité de la NDF, l’interprétation des résultats est difficile en raison d’une variété de facteurs. Il existe très peu d’expériences ayant
comparé des fourrages qui différaient seulement au niveau de la digestibilité de la NDF, et il est difficile de confirmer qu’aucun facteur autre que la digestibilité des fibres n’a pu faire varier les performances animales (Oba et Allen, 1999). Par exemple, lorsqu’on compare deux fourrages inclus dans des rations dont les ratios fourrages : concentrés sont différents, d’autres facteurs, comme la digestion ruminale de l’amidon, peuvent affecter la CVMS ou la production laitière (Oba et Allen, 1999). Aussi, les études comparant des fourrages incorporés au même taux dans les rations, mais dont la concentration en NDF diffère ont des résultats biaisés par les différents effets de remplissage du rumen liés aux concentrations en NDF (Oba et Allen, 1999).
Il faut également considérer qu’une étude en particulier comparant des fourrages de qualité différente n’est que l’une des nombreuses situations possibles en matière d’alimentation (Allen, 1996b). L’extrapolation des résultats à différentes conditions peut être trompeuse en raison de la complexité des interactions entre le fourrage et la population microbienne du rumen, les facteurs animaux et les autres ingrédients de la ration (Allen, 1996b). En effet, les résultats expérimentaux peuvent varier en fonction du niveau de production animale, du stade de lactation, du niveau et du type de supplémentation en protéines, du niveau et du type de supplémentation en énergie, de la taille des particules du fourrage, de la génétique du fourrage, des conditions de croissances, de la méthode de conservation et de plusieurs autres facteurs (Allen, 1996b).
Afin de minimiser les risques de mauvaise interprétation, seulement les expériences ayant mesuré une différence significative de digestibilité in vitro ou in situ seront présentées. En effet, les digestibilités de la NDF
in vitro ou in situ sont de meilleurs indicateurs que la digestibilité de la NDF in vivo pour évaluer le potentiel des
fourrages d’augmenter la CVMS (Oba et Allen, 1999). Pour les expériences comparant des cultivars de luzerne, une différence significative de digestibilité ne sera pas nécessaire pour que l’étude soit incluse dans cette revue de littérature.
1.1.5.1 Consommation
Selon Allen et al. (2009), la CVMS dans les premières semaines de la lactation est contrôlée principalement par l’oxydation des carburants dans le foie qui envoie des signaux de satiété au cerveau. Lorsque la vache sort d’un bilan énergétique négatif, plusieurs semaines après le vêlage, la CVMS commence à augmenter tandis que la lipolyse et la concentration plasmatique en acides gras non estérifiés diminuent, ce qui diminue la disponibilité des acides gras non estérifiés pour l’oxydation dans le foie, entrainant une diminution du contrôle de la prise alimentaire par l’oxydation hépatique (Allen et al., 2009). La distension ruminale devient donc le mécanisme dominant du contrôle de la consommation dans la période entourant le pic de lactation (Allen et al., 2009).
Le remplissage du rumen est affecté par le contenu en NDF de la ration, mais aussi par la digestibilité de la NDF du fourrage (Oba et Allen, 1999). La digestibilité serait d’ailleurs le facteur le plus important influençant la
CVMS des fourrages ensilés (Huhtanen et al., 2007). En effet, augmenter la fraction des fibres qui est potentiellement digestible permettrait à une plus grande partie des fibres du rumen de s’éliminer par la digestion et le passage, ce qui entraine une consommation additionnelle d’aliments (Dado et Allen, 1996). Toutefois, même si un plus haut taux de digestion et de passage vont réduire l’effet de remplissage (Allen et Mertens, 1988), l’effet de la fraction potentiellement digestible de la NDF n’est pas encore clair (Allen, 2000). Alors qu’une plus grande fraction de NDF potentiellement digestible va résulter en une amélioration de la digestibilité de la NDF à un taux de rétention constant, le temps de rétention des particules dans le réticulo-rumen pourrait aussi être plus long en raison de la plus grande flottabilité des particules dans le temps (Allen, 1996a).
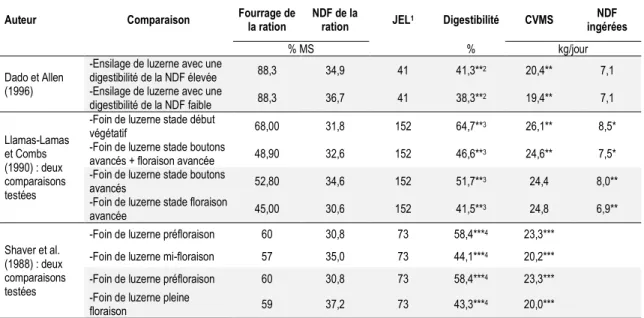
Certaines expérimentations animales avaient pour but de comparer l’utilisation de luzernes, sans toutefois que la différence de digestibilité soit liée au génotype (Tableau 1.5 ; Shaver et al., 1988; Llamas-Lamas et Combs, 1990; Dado et Allen, 1996). Tous les auteurs ayant mesuré l’effet d’une augmentation de la digestibilité de la luzerne (sans être associée au cultivar) ont noté une augmentation de la CVMS (Shaver et al., 1988; Llamas-Lamas et Combs, 1990; Dado et Allen, 1996), à l’exception d’une comparaison (Llamas-Llamas-Lamas et Combs, 1990). Il est toutefois important de rappeler que beaucoup d’autres variables peuvent aussi avoir eu un effet sur la consommation, puisque ce n’est pas seulement la digestibilité qui a changé dans ces luzernes.
Tableau 1.5 Sommaire des expériences ayant porté sur la digestibilité de la luzerne à différents stades de
maturité.
Auteur Comparaison Fourrage de la ration NDF de la ration JEL1 Digestibilité CVMS NDF
ingérées
% MS % kg/jour
Dado et Allen (1996)
-Ensilage de luzerne avec une
digestibilité de la NDF élevée 88,3 34,9 41 41,3**2 20,4** 7,1 -Ensilage de luzerne avec une
digestibilité de la NDF faible 88,3 36,7 41 38,3**2 19,4** 7,1 Llamas-Lamas et Combs (1990) : deux comparaisons testées
-Foin de luzerne stade début
végétatif 68,00 31,8 152 64,7**3 26,1** 8,5*
-Foin de luzerne stade boutons
avancés + floraison avancée 48,90 32,6 152 46,6**3 24,6** 7,5* -Foin de luzerne stade boutons
avancés 52,80 34,6 152 51,7**3 24,4 8,0**
-Foin de luzerne stade floraison
avancée 45,00 30,6 152 41,5**3 24,8 6,9**
Shaver et al. (1988) : deux comparaisons testées
-Foin de luzerne préfloraison 60 30,8 73 58,4***4 23,3***
-Foin de luzerne mi-floraison 57 35,0 73 44,1***4 20,2***
-Foin de luzerne préfloraison 60 30,8 73 58,4***4 23,3***
-Foin de luzerne pleine
floraison 59 37,2 73 43,3***4 20,0***
† : P ˂ 0,10 ; * : P ˂ 0,05 ; ** : P ˂ 0,01 ; *** : P ˂ 0,001. 1Moyenne des jours en lactation à la mi-expérience. 2NDFD in vitro 24 heures de la ration.
3NDF potentiellement digestibles des fourrages estimée in situ. 4NDF potentiellement digérées in situ.