THÈSE
Présentée pour obtenir le Diplôme de
Doctorat d’Etat Es-Sciences Biologiques
Présentée par
Nom et Prénom : Fassi Fihri Aicha
Discipline : Biologie
Spécialité : Biotechnologie
Titre :
Collecte et maturation des ovocytes bovins : effet de
l’état nutritionnel sur le rendement
et la qualité des ovocytes
Soutenue le 28 Mars 2006
Devant le jury,
Président :
Mr. Benjelloun W.
(Doyen, de la la Faculté des Sciences de Rabat)
Examinateurs :
Mme. Hajji Kh.
(Professeur à la Faculté des Sciences de Rabat)
Mr. Derqaoui L.
(Professeur à l’Institut Agronomique et Vétérinaire Hassan II, Rabat)
Mr. Lakhdissi H.
(Professeur à l’Institut Agronomique et Vétérinaire Hassan II, Rabat)
Mme. Kharbach A.
(Professeur, Directeur de la maternité du Centre Hospitalier et Universitaire du Souissi, Rabat)
Mr. Tahri-Joutei A.
Avant Propos
Le présent travail a été réalisé sous la codirection de Mme Khadija Hajji,
Professeur au laboratoire de Zoologie et de Biologie Générale de la Faculté des
Sciences de Rabat et de Mr Hassan Lakhdissi, Professeur au Département de
Reproduction Animale, d’Obstétrique et d’Insémination Artificielle de l’Institut
Agronomique et Vétérinaire Hassan II, Rabat.
J’adresse mes vifs remerciements à Mme le Professeur Khadija Hajji de
m’avoir introduit dans ce domaine passionnant. Elle a toujours porté un intérêt
particulier et suivi de près toutes les étapes de réalisation de ce travail.
C’est grâce au projet P.A.R.S. (Biol, 102) qui est sous sa direction que ce
travail a pu être lancé. Qu’il me soit permis de lui exprimer mes sincères et
profondes reconnaissances.
Monsieur le Professeur Hassan Lakhdissi a eu l’amabilité de me proposer ce
sujet et de m’accorder une place au sein du Département de Reproduction
Animale, d’Obstétrique et d’Insémination Artificielle de l’Institut Agronomique
et Vétérinaire Hassan II. Il a suivi pas à pas et avec beaucoup de patience
l’évolution de mon travail, évolution facilitée par ses conseils très utiles, ses
discussions enrichissantes.
Il m’a toujours accordé un encadrement attentionné. Sans son aide, sa
générosité, ses conseils, ses critiques permanentes lors de la rédaction des
articles et de la thèse, ce travail n’aurait pas pu voir le jour.
Je suis sensible au dévouement de Monsieur Lahsen Derqaoui, Professeur au
Département de Reproduction Animale, d’Obstétrique et d’Insémination
Artificielle de l’Institut Agronomique et Vétérinaire Hassan II.
Monsieur le Professeur Lahsen Derqaoui a porté un intérêt particulier pour
cette étude, ses efforts, ses directives et le temps précieux qu’il m’a consacré
pour la réalisation de ce travail malgré ses multiples occupations. Son
expérience, sa compétence et ses conseils très enrichissants m’ont énormément
facilité la tache. Qu’il me soit permis de lui exprimer mes sincères et profondes
reconnaissances.
J’adresse mes vifs remerciements au Professeur Wail Benjelloun, Doyen de la
Faculté des Sciences de Rabat, pour avoir accepté de juger ce travail.
Je suis très sensible à l’honneur que vous me faites en acceptant de présider
mon jury de thèse. Veuillez trouver ici, Monsieur le Doyen, l’expression de
notre sincère gratitude.
Je tiens aussi à remercier le Professeur Abderrafih Tahri-Joutei, Professeur à
la Faculté Des Sciences de Rabat pour avoir accepter de juger ce modeste
travail.
Mes remerciements s’adressent aussi au Professeur Aicha Kharbach, Directeur
de la maternité du centre Hospitalier et Universitaire du souissi, Rabat, qui a
accepté d’être parmi les membres de jury de cette thèse.
Ces presque huit ans furent comme toujours longues et courtes à la fois. De
nombreuses personnes ont, à un moment ou à un autre, intervenu dans le cours
des événements qui ont peu à peu conduit à l’élaboration de ce travail.
Un grand merci à Mme Mariam Naciri Professeur au laboratoire de Zoologie
et de Biologie Générale de la Faculté des Sciences de Rabat, puisse ce travail
être le témoignage de notre longue amitié.
J’exprime ma profonde gratitude et mon profond respect à Mr Mohamed El
Hamidi, Professeur au Département d’Histologie et Anatomie pathologique de
Mes remerciements à Messieurs Zaid Zouagui Professeur au Département de
Pathologie Médicale et Chirurgicale des ruminants et Mr. Abdelouahed Bassir
du même département.
Je remercie tous les membres du Département de Reproduction, d’Obstétrique
et d’insémination artificielle, particulièrement Pr. Mazouz A., Pr. Sghiri A.
Medames Faraj R., El Idrissi N. Messieurs, Zaizaa B., Bennani M. et Bouhila
A.
Un grand merci au professeur Mohamed Oukessou, chef du département de
Physiologie et Thérapeutique à l’IAV Hassan II (Rabat), pour avoir bien voulu
m’accueillir au sein de son département, pour ses conseils et ses directives qui
m’ont été d’un grand intérêt. Je remercie également tous les enseignants du
Département pour leur sympathie particulièrement le Professeur El Guerouali
A., Messieurs Suilah S. et Ablouh.
J’adresse mes vifs remerciements à Mr le Professeur Sefiani, chef du
Laboratoire de Génétique de l’Institut d’Hygiène, qui a bien voulu m’accueillir
au sein de son laboratoire et où j’ai pu faire les premières cultures cellulaires.
Je remercie également tous les chercheurs et les médecins du laboratoire pour
leur sympathie.
Mes remerciements s’adressent également à mes collègues et amies du
Laboratoire de Zoologie et de Biologie Générale de la Faculté des Sciences de
Rabat pour l’aide qu’ils m’ont apportée à divers titres, tout particulièrement,
Mmes Benazzou Touria, Kadiri Zineb, Zouiten habiba, Benmessaoud Fatma,
Kourradi Rhita et Mr Benhoussa Aziz.
Je ne saurai terminer ce préliminaire sans évoquer l’appui moral et la
sollicitude que j’ai trouvés auprès de ma mère, mon père, mon mari et mes
INTRODUCTION GÉNÉRALE ...13
Première Partie : SYNTHÈSE BIBLIOGRAPHIQUE Chapitre I : Caractéristiques de la reproduction chez la vache...17
I/ Développement embryonnaire des ovaires ...17
1/ Constitution et régulation du stock de follicules Primordiaux...18
2/ La folliculogenèse...18
3/ Aspects morphologiques de la croissance folliculaire...20
3.1/ Les follicules pré-antraux...21
3.2 / Les follicules à antrum...21
4/ Entrée en méiose...24
II / Puberté...24
III/ Cycle sexuel...25
1/ Composante anatomique...28
1.1/ Vagues folliculaires...28
1.2/ Croissance et maturation de l’ovocyte... 29
1.2.1/ Croissance de l’ovocyte...29
1.2.2/ Maturation de l’ovocyte...30
a/ Aspects cytologiques de la maturation des ovocytes...31
a.1/ Rupture de la vésicule germinale...31
a.2/ Condensation des chromosomes...31
a.3/ Formation du fuseau chromosomique...31
a.4/ Passage et arrêt en métaphase II...31
a.5/ Maturation cytoplasmique...32
b/ Aspects moléculaires de la maturation de l’ovocyte...32
1.3/ Ovulation... 33 1.4/ Corps jaune...34 2/ Composante hormonale...35 2.1/ Hormones hypothalamiques...35 2.2/ Gonadotrophines hypophysaires, (FSH et LH)...………...36 2.3/ Hormones ovariennes...37 2.4/ Autres hormones...37
a/ Prostaglandines...37 b/ Ocytocine...38 2.5/ Régulations hormonales ...38 a/ Contrôle de la sécrétion de la GnRH...38 b/ Contrôle de la sécrétion de LH et de FSH...39 3/ Composante comportementale...40 3.1/ Saisonnalité...40 3.2/ Œstrus.
...40
Chapitre II : Facteurs de variation de la production des ovocytes et de leur qualité...42 I/ Facteurs endogènes...42 1/ Race ...42 2/ Age ...42 3/ Etat physiologique...43 4/ Phase du cycle...43 II/ Facteurs exogènes :………...44
A. Nutrition...44
1/ Déficit énergétique...44
1.1/ Mode d’action du déficit énergétique...45
1.2/ Déficit énergétique et établissement de la puberté : Rôle de la leptine...46
1.3/ Déficit énergétique et croissance folliculaire...47
1.4/ Déficit énergétique et composante hormonale...48
1.5/ Indicateurs de la modification du statut énergétique...51
a/ B.C.S. (Body Condition Score ou note de l’état corporel)...51
b/ Paramètres métaboliques...52
2/ Excès protéiques...53
2.1. Mode d’action d’un excès protéique...54
2.2. Indicateurs sériques du métabolisme azoté...55
b/ L’albumine...55
c/ L’urée...55
3/ Vitamines et Minéraux...56
a/ Vitamines...56
b/ Minéraux...57
B. Stress et maladies intercurrentes...58
Chapitre III : Collecte et maturation des ovocytes...59
I/ Facteurs de variation du rendement de la collecte des ovocytes...59
1/ Disposition des follicules dans le cortex ovarien (facteur endogène) ...59
2/ Technique de collecte (facteur exogène) ...59
II/ Maturation des ovocytes in vitro...60
1/ Le milieu de base...60
2/ Hormones...61
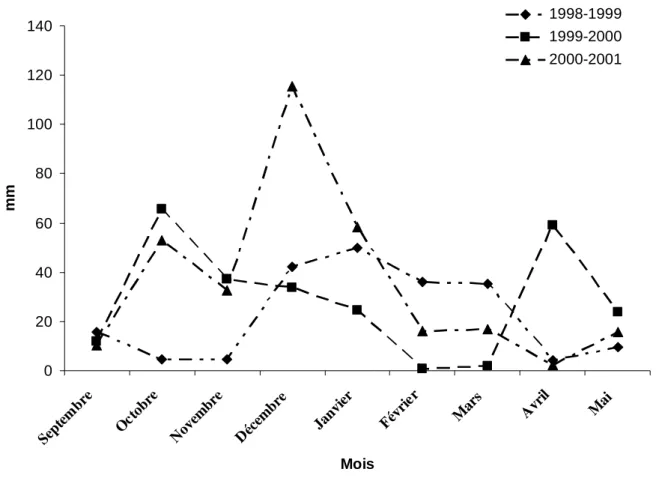
Deuxième Partie : MATÉRIEL ET METHODES I/ Conditions pluviométriques des années 1999 et 2000...63
II/ Animaux de l’étude...63
III/ Protocole expérimental...66
1/ Détermination de l’âge...66
2/ Note de l’état orporel...66
3/ Prélèvement des ovaires ...67
4/ Etude histologique des coupes d’ovaires...67
a. Préparation des coupes histologiques pour microscopie photonique...67
b. Méthode d’évaluation de la population folliculaire...68
5/ Prélèvements sanguins...68
6/ Les méthodes de dosage des Protéines totales, albumine, urée, β-OH et A.S.A.T...69
7/ Ponction des follicules et collecte des ovocytes...69
b. Collecte des ovocytes………..69
c. Composition du milieu de collecte……….69
8/ Evaluation morphologique de la qualité des ovocytes ...69
9/ Maturation des ovocytes In Vitro (M.I.V.)...70
a. Composition du milieu de maturation...70
b. Mise en culture...70
IV/ Analyses statistiques...70
Troisième Partie : RESULTATS Chapitre I : Population folliculaire et rendement en ovocytes...73
I/ Population folliculaire ...73
II/ Rendement en ovocytes et leur taux de maturation...74
Chapitre II : Exploration histologique de l’ovaire...77
I/ Structure de l’ovaire...77
II/ Evaluation de la population folliculaire...79
Chapitre III : Effet des paramètres génétiques et non génétiques sur la population folliculaire et le rendement en ovocytes...81
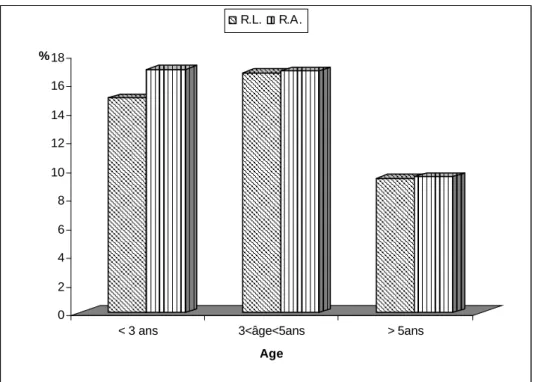
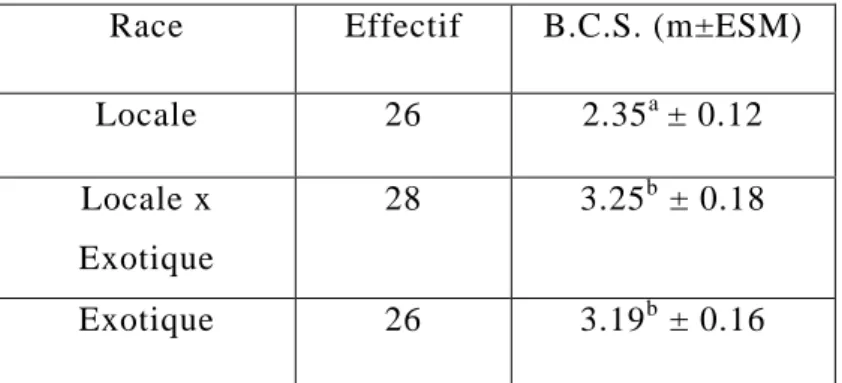
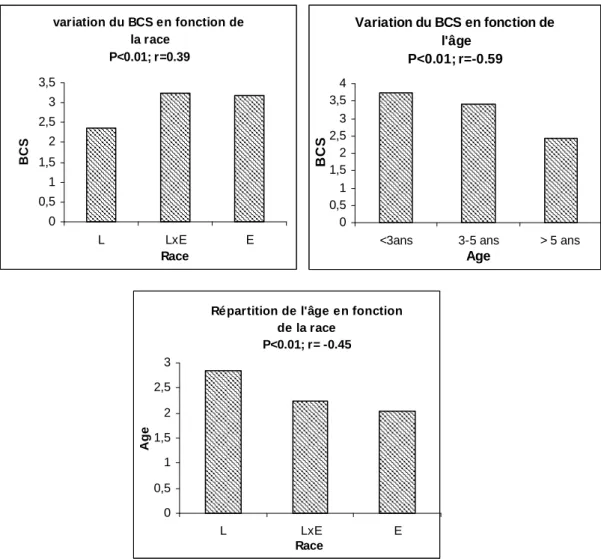
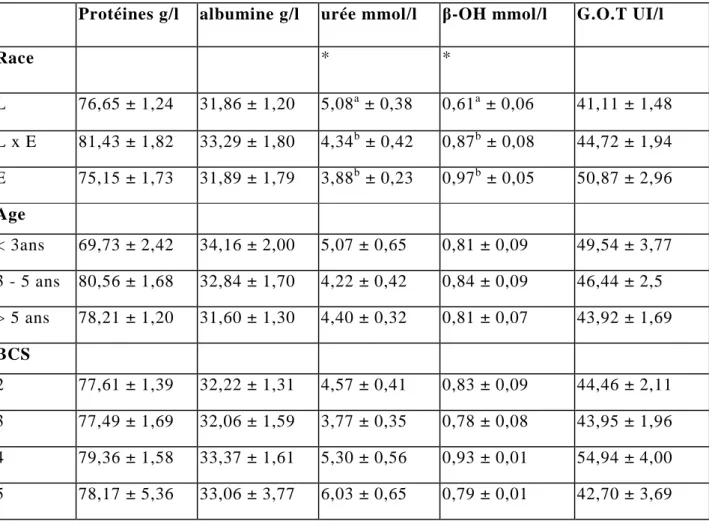
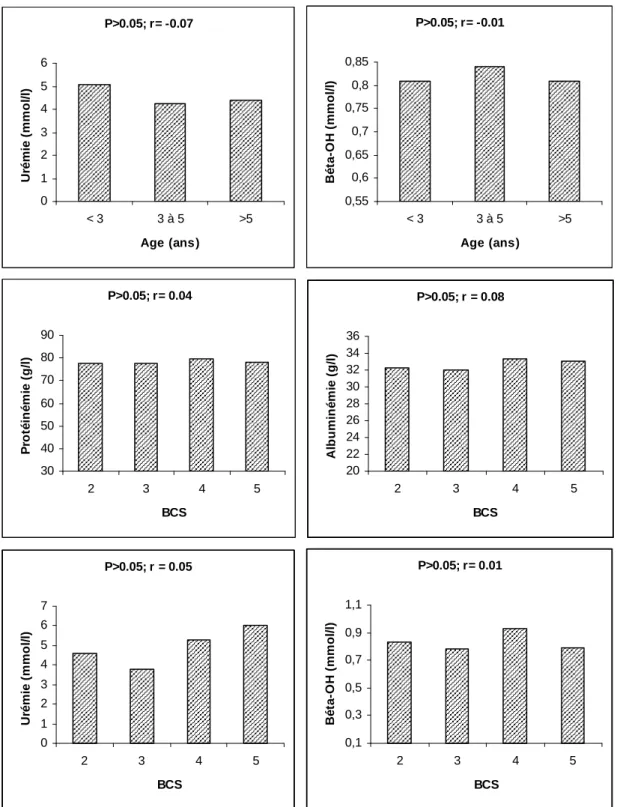
I/ Résultats descriptifs...81 1/ Race, âge et B.C.S...………81 a/ Race...81 b/ Age...81 c/ Etat corporel (B.C.S.) ...81 2/ Paramètres sanguins...83
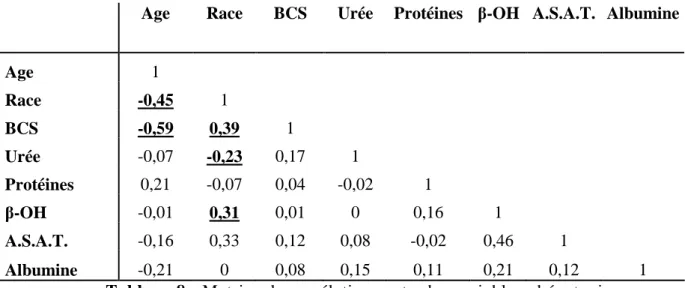
3/ Corrélations entre les différents paramètres ...85
a/ B.C.S –Age...85
b/ B.C.S. – Race...85
c/ Race – Age...86
d/ Race – Urémie...87
g/ L’albumine...90
h/L’A.S.A.T...90
II/ Résultats analytiques...92
1/ Nombre de follicules...92
1.1/ Effet de la race ...93
1.2/ Effet de l’âge ...93
1.3/ Effet du B.C.S. ...93
1.4/ Effet des paramètres biochimiques...96
a/ Taux d’albumine sérique ...96
b/ La protéinémie...96
c/ L’urémie...96
d/ Taux sérique de β-OH...97
2/ Rendement en ovocytes...98
2.1/ Effet de la race ...99
2.2/ Effet de l’âge ...99
2.3/ Effet B.C.S...99
2.4/ Effet des paramètres biochimiques...102
a/ Taux d’albumine sérique ...102
b/ La protéinémie ...102
c/ L’urémie ...102
d/ Taux sérique de β-OH ...103
3/Qualité des ovocytes...104
3.1/ Effet de la race, de l’âge et de l’état corporel...104
3.2/ Effet des paramètres biochimiques ...107
a/ Taux d’albumine sérique ...107
b/ La protéinémie ...108
c/ L’urémie ...108
d/ Taux sérique de β-OH ...109
Quatrième Partie : DISCUSSION ET CONCLUSION GÉNÉRALE...112
ANNEXES...148 TRAVAUX SCIENTIFIQUES…...……….162
INTRODUCTION
GENERALE
Le secteur de l’élevage constitue l’une des composantes majeures de l’économie agricole du Maroc. Il participe à 30% de la valeur ajoutée agricole, emploie pratiquement 20% de la population rurale active et fournit les matières premières (lait, viande, peaux, laine) à plusieurs secteurs agro-industriels.
Parmi les espèces domestiques exploitées à l’échelle nationale, figurent les bovins qui participent à plus de 50% dans l’approvisionnement du pays en viande rouge et à presque 90% dans son ravitaillement en lait. En 1999, les productions des bovins ont atteint 130 000 tonnes de viande et 1.130 milliards de litres de lait (M.A.D.R.P.M., 2000).
Cependant, la productivité reste basse malgré la diversité de ses races, du essentiellement à des problèmes zootechniques, sanitaires et de reproduction qui demeure une reproduction traditionnelle mal planifiée (M.A.D.R.P.M., 1998).
L’emploi de nouvelles techniques de reproduction : l’Insémination Artificielle (I.A) (Marquant-LeGuienne, 1986 ; Zakaria, 2001), le Transfert Embryonnaire (T.E.) (El Aidi
et al., 1995) et la Production d’Embryons in Vitro (P.I.V.) (Thibier, 1990; Marquant-Le
Guienne, 1991 ) contribue à l’intensification de l’amélioration génétique du cheptel bovin, l’I.A. par la voie du male, le T.E et la P.I.V. par la voie de la femelle.
Au Maroc, l’insémination artificielle et le transfert embryonnaire ont constitué un apport appréciable dans l’amélioration de la structure génétique du cheptel bovin, à travers l’importation de semences des races performantes (Frisonnes, Holstein,….) et leur croisement avec le type local (Benlekhal, 1996). Les expériences menées en T.E. ont été localisées mais très concluantes (El Aidi et al., 1995).
Cependant, la P.I.V. d’embryons bovins qui constitue la troisième génération des biotechnologies de la reproduction, s’est affirmée et a progressé rapidement ces dernières années chez les mammifères comme l’atteste le nombre considérable de publications sur ce sujet n’a jamais été tentée à notre connaissance au Maroc.
En effet, chez les bovins, la P.I.V. d’embryons est très utilisée dans le monde et permet la production d’embryons à partir d’ovocytes issus d’ovaires prélevés aux abattoirs dans le but de promouvoir la multiplication de cette espèce (Dauzet et Marquant-LeGuienne, 1994).
Chez les bovins, le premier veau obtenu par Fécondation In Vitro (F.I.V.) d’un ovocyte, maturé in vivo est né en 1981 (Brackett et al., 1982). Depuis cette date, les processus de maturation, de fécondation et de culture in vitro ont été considérablement améliorés (Brachett et al., 1982 ; Marquant-Le Guienne, 1991 ; Brachett et Zuelke, 1993 ;
nouvelle biotechnologie dans les programmes d’élevage et également le développement de laboratoires spécialisés dans la production en masse d’embryons produits totalement in vitro. L’intérêt économique d’une telle source d’embryons est très positif (Dauzet et Marquant-Le Guienne, 1994).
D’autre part, la fécondation in vitro est une technique qui permet l’étude des mécanismes biologiques se produisant in vivo (Drion et al., 1996).
De même, le développement embryonnaire in vitro permet de contrôler et de suivre la croissance des œufs jusqu’au stade compatible avec la transplantation utérine de l’embryon ou sa congélation.
La production d’embryons bovins in vitro comprend différentes étapes : la maturation des ovocytes, la capacitation des spermatozoïdes, la fécondation et enfin la culture des embryons. La possibilité d’intervenir à chacune de ces étapes offre de nombreux intérêts à la P.I.V.
Cette dernière est une chaîne d’opérations dans laquelle l’ovocyte ou gamète femelle intervient en amont et détermine la réussite de la technique.
La maturation des ovocytes in vitro est l’étape initiale importante qui détermine la qualité de cette cellule pour l’obtention d’embryons transférables après fécondation in vitro. Elle comprend une maturation nucléaire et une maturation cytoplasmique, nécessaires pour le déroulement normal de la fécondation.
En effet, elles permettent à l’ovocyte d’être reconnu et pénétré par le spermatozoïde, d’assurer la formation simultanée des pronuclei et leur développement ultérieur jusqu’au stade morula ou blastocyste (Dieleman et al., 1983 ; Mermillod, 2001).
En outre, la maturation in vitro joue un rôle important pour le clonage. Elle permet l’obtention d’une source d’ovocytes « receveurs » (en métaphase II) à coût réduit et en grand nombre (Heyman et al., 1990).
Le taux de réussite de la production d’embryons in vitro rapporté dans la littérature varie de 19% à 38% selon la méthodologie et les conditions de travail (Carolan et al., 1996 ; Beker et al., 2002 ; Chian et al., 2002 ; Avery et al., 2003), la proportion d’ovocytes qui arrivent au stade blastocyste est de 12%, 13%, 23% et 16% quand ils sont collectés respectivement au 2ème, 3ème, 5ème et 7ème jour de l’émergence de la vague folliculaire (jour :0) .
Des facteurs intrinsèques et extrinsèques à la donneuse d’ovocytes tels que les aspects biochimiques, hormonaux et nutritionnels (Kendrick et al., 1999) ont été rapportés comme étant à l’origine des fluctuations des résultats dans un même laboratoire.
Bien que les pourcentages de maturation obtenus in vitro soient encourageants dans plusieurs laboratoires, cette technique n’a jamais été appliquée sur les ovocytes de la race locale Marocaine. En l’occurrence, la race Oulmès-Zaer qui a fait l’objet de plusieurs opérations de production et de transfert d’embryons qui ont eu lieu dans la région d’Oulmès.
Le taux de réussite de récoltes et transferts d’embryons chez cette race était de 40% (El Aidi et al., 1995).
Cependant, le transfert d’une technologie doit débuter par une phase d’adaptation aux conditions spécifiques du nouveau milieu. En effet, le nombre d’embryons qui résulte de ces opérations varie entre pays, régions, et laboratoires.
Les objectifs de ce travail se résument à:
1/ La maîtrise de la technique : collecte des ovocytes et leur maturation in vitro.
2/ L’exploration Histologique de la fonction ovarienne par l’étude de la population folliculaire et leur disposition dans le cortex ovarien chez la race locale et ses produits de croisement avec des races exotiques.
3/ Détermination de l’impact du niveau alimentaire, de l’âge et la race sur la population folliculaire et le rendement en ovocytes.
PREMIERE PARTIE
SYNTHESE BIBLIOGRAPHIQUE
CHAPITRE I : Caractéristiques de reproduction chez la vache I/ Développement embryonnaire des ovaires.
L’origine embryologique des ovaires est mixte. Les cellules germinales souches d’abord extra embryonnaire colonisent, après migration au travers de l’embryon, une zone dense de tissu mésenchymateux supra-mésonéphrotique recouverte d’épithélium, les corps de Wolff (Drion et al., 1996).
Les ovaires résultent de la différenciation spontanée des corps de wolff qui recouvrent le mésonéphros de l’embryon sexuellement indifférencié. Chez les mammifères, cette différenciation s’opère à partir de la septième semaine de développement embryonnaire, période à laquelle est accomplie la migration des cellules germinales primordiales dans l’épithélium cœlomique. Les corps de Wolff prolifèrent et se condensent pour former une crête longitudinale bilatérale, appelée crête génitale (ou bourrelet germinal), située en région lombaire (Magre et Vigier, 2001).
Initialement en position postérieure au niveau de la paroi de la vésicule vitelline du fœtus (Snow et Monk, 1983), les cellules germinales souches migrent activement le long du mésentère dorsal de l’intestin postérieur pour atteindre la crête génitale (Figure 1). La fibronectine, présente tout au long de la voie empruntée (Hoffman et al., 1974), constitue l’inducteur de cette migration. Les réarrangements des tissus en formation semblent constituer aussi un facteur mécanique de cette migration en conjonction avec des substances chémotactiques d’origine gonadique (Snow et Monk, 1983).
Au cours de cette migration, et quelques fois après (Derivaux et Ectors, 1989), se produit une multiplication mitotique des cellules germinales souches.
La survie et la prolifération de ces cellules semblent être en rapport avec une collaboration entre cellules somatiques et facteurs de croissance.
En effet, deux éléments sont actuellement identifiés (Drion et al., 1996): - Les genes dominants « white spotting (W) » et « Steel gène (S1) »,
- des facteurs de croissance tel le LIF « Leukemia Inhibitor Factor » et le bFGF « basic Fibroblast Growth Factor ».
Le gène W code pour un récepteur cellulaire de type tyrosine kinase. Le gène S1 code pour un facteur de croissance peptidique, le « stem » ou « mast cell growth factor ». Il est le ligand de ce récepteur. Le produit du gène S1, en interagissant avec le récepteur du gène W, stimulerait les cellules somatiques pour produire des facteurs de croissance nécessaires à la multiplication des cellules germinales.
1/ Constitution et régulation du stock de follicules primordiaux
Chez les mammifères, la phase de multiplication des ovogonies par mitoses successives est en général terminée avant ou peu après la naissance. Les singes Lémurs de Madagascar constituent l’exception à cette règle. Ils présentent une multiplication des ovogonies qui persiste à l’âge adulte (Drion et al., 1996).
Chez les bovins, la multiplication des ovogonies s’étend du 45ème au 150ème jour de la vie intra- utérine et engendrent des ovaires à jusqu'à 2 millions d’ovogonies durant la vie fœtale. Si tôt la phase mitotique terminée, ces dernières entament le processus de méiose qui s’interrompt en prophase I (stade dictyetène) et deviennent ainsi les ovocytes I.
La phase de multiplication des ovogonies permet la constitution d’un stock folliculaire dont l’importance dépend de l’espèce :
160 000 chez la brebis, 235 000 chez la vache, 20 000 chez la rate,
et 1000 000 chez la femme. Le stock dépend aussi de la race, de l’âge, du niveau hormonal et du statut de reproduction des individus (Driancourt et al., 1991).
Le nombre de follicules primordiaux reste apparemment stable jusqu'à la 4ème année de vie chez la vache, puis décline pour s’annuler vers la 20ème année (Drion et al., 1996). Seuls les ovocytes I qui s’entourent de quelques cellules folliculaires et d’une lame basale (future membrane de slavjanski) persistent pour former les follicules primordiaux. Ainsi, bien que le nombre d’ovocytes I « culmine » durant la vie utérine, il n’en reste qu’un petit nombre à la naissance (Drion et al., 1996) .
2/ La folliculogenèse
La folliculogenèse est l’ensemble des phénomènes qui caractérisent l’apparition, la croissance, et la maturation des follicules. C’est encore la succession des différentes étapes du développement du follicule depuis le moment où il sort de la réserve jusqu'à l’ovulation, où au cas le plus fréquent jusqu'à l’atrésie (Driancourt et al., 2001a). En effet, plus de 99% des follicules sont voués à l’atrésie, ils dégénèrent sans avoir pu évoluer jusqu’à terme (Drion et al., 1996) .
Comme chez la plupart des espèces de mammifères, la folliculogenèse chez les bovins commence pendant le développement fœtal (Maneesh et al., 2000). Les follicules antraux sont observés dans les ovaires des fœtus vers la fin de la gestation.
La genèse des follicules dans l’ovaire des mammifères résulte d’une interaction complexe entre prolifération, différenciation et atrésie à l’intérieur des vagues de follicules en voie de maturation (Greenwald et Roy, 1994).
Les premiers événements détectables dans la transformation des follicules primordiaux en follicules initiés à la croissance sont :
- l’incorporation de la thymidine tritiée par les cellules folliculeuses
- l’augmentation de la taille de l’ovocyte associée à une synthèse d’ARN et la formation de la zone pellucide qui est responsable de la spécificité d’espèce pour la reconnaissance par le spermatozoïde (Scaramuzzi et al., 1993 ; Drion et al., 1996).
Chez les mammifères, il existe deux phases dans la croissance des follicules dont les mécanismes de régulation sont différents (Driancourt et al., 2001b) :
- La phase de croissance folliculaire basale (du stade primordial au stade pré-antral) essentiellement dépendante de facteurs de croissance (GDF-9, Steel, IGF-1) ou endocriniens (insuline) (Canty et al., 2003);
- La phase de croissance terminale c’est à dire du stade cavitaire qui varie en général chez les mammifères entre 2 et 5mm, en particulier chez la vache : 2 à 3mm à l’ovulation (Austin et al., 2002). Elle correspond à la durée de la phase folliculaire du cycle : 4 jours chez la ratte, 2 à 3 jours chez la brebis, 5 jours chez la truie et la vache, 14-17 jours chez les primates.
3/ Aspects morphologiques de la croissance folliculaire
La phase de croissance folliculaire est définie comme l’intervalle entre le moment où le follicule quitte la réserve et le moment où il atteint l’ovulation. Elle est contemporaine de la croissance de l’ovocyte que le follicule contient (Driancourt et al., 2001a).
Les premières nomenclatures désignaient les follicules selon le stade de développement : follicules primordiaux, follicules en croissance et follicules vésiculaires (Hulshof et al., 1994).
Une nomenclature propre aux follicules en croissance est actuellement en vigueur (Figures 2, 3) :
- les follicules primordiaux, les follicules primaires et les follicules secondaires correspondent aux follicules préantraux ;
- le terme de follicule de De Graaf ne s’applique qu’au follicule mûr.
3.1/ Les follicules pré-antraux
Quand 2 à 3 couches de cellules folliculeuses sont formées, les cellules thécales se différencient à partir du stroma et s’orientent en couches concentriques autour de la membrane basale qui se transforme en membrane de Slavjanski faite de fibres de collagène type IV, fibronectine, laminine et protéohéparane sulfate.
Ces follicules sont alors formés de deux ou plusieurs couches de cellules de la granulosa. Leur diamètre est compris entre 0.05 et 0.1 mm.
Un follicule de 0,1 mm de diamètre comprend 10 000 cellules de granulosa environ chez la brebis (Scaramuzzi et al., 1993). A ce stade, le développement du follicule se caractérise par une augmentation de la taille de l’ovocyte associée à une prolifération active des cellules folliculaires (Mariana et al., 1991). Dès lors, le follicule préantral, possédant des récepteurs à la LH dans la thèque interne et à FSH dans la granulosa, est potentiellement capable de répondre à une stimulation gonadotrope (Driancourt et al., 1991). Chaque follicule comporte une vascularisation thécale, permettant le transport de régulateur endocrine varié tel les IGF (Insuline-like Growth Factor). Des récepteurs à IGF1 sont présents sur la thèque et la granulosa du follicule (Gougeon, 1996). Enfin, des expériences de culture in vitro ont montré qu’il existerait des régulations entre follicules (régulations paracrines) (Driancourt, 1997).
3.2/ Les follicules à antrum
Des espaces apparaissent entre les cellules de la granulosa des follicules de grande taille. L’antrum apparaît d’abord, sous forme d’antrum diffus, comblant les espaces entre les cellules et fusionnant ensuite en une seule cavité (Mariana et Machado, 1976). Les thèques sont alors différenciées autour du follicule : thèque interne, d’aspect glandulaire riche en cellules et capillaires et thèque externe riche en fibres de collagène (Driancourt et al., 2001a) .
Pendant la formation de l’antrum, l’ovocyte atteint sa taille presque définitive (80%). À partir de ce moment, le follicule s’accroît car la prolifération cellulaire continue mais surtout, parce que l’antrum s’agrandit (Mariana et al., 1991). Au terme de sa
4/ Entrée en méiose
La date d’entrée en méiose des ovogonies se produit avant la naissance. Elle débute spontanément, ou plus vraisemblablement, sous l’influence d’un facteur d’origine mésonéphrotique, le MIS (Meiosis inducing substance) (Westergaard et al., 1985 ; Dominko et First, 1997).
La substance induisant la méiose (MIS) est produite par les cellules du bord interne de l’ovaire, dérivées du rete ovarii. C’est à cet endroit qu’a lieu la première rencontre entre les cellules germinales et les cellules somatiques.
La compétence de l’ovocyte à terminer spontanément sa méiose (maturation nucléaire) hors du follicule est acquise lorsqu’il est issu d’un follicule de diamètre supérieur à 2 mm. Ensuite, l’acquisition de la compétence au développement (maturation cytoplasmique) augmente progressivement avec la taille du follicule dont l’ovocyte est issu (Cognié et Baril, 2002).
In vivo, la compétence au développement (reprise de la méiose) des ovocytes est influencée par leur environnement folliculaire (Mermillod et al., 1999 a), lui-même régulé pendant la phase folliculaire du cycle sexuel par le niveau de pulsatilité de LH (Oussaid et al., 1999).
II / Puberté
C’est le moment où l’individu acquiert la fonction sexuelle c’est à dire devient apte à produire un gamète fécondant (chez le mâle) ou fécondable (chez la femelle); c’est un préalable nécessaire à la mise à la reproduction. L’apparition de la puberté est repérée concrètement dès que les premiers signes de l’activité sexuelle sont visibles : première chaleur chez la femelle. Le déterminisme de l’apparition de la puberté provient de la mise en place et du fonctionnement du système hormonal relatif à la reproduction, impliquant l’hypothalamus, l’hypophyse et les gonades. Ce système contrôle l’apparition du comportement sexuel, la mise en place de l’évolution des caractères sexuels primaires et secondaires conduisant à l’existence du dimorphisme sexuel (Bonnes et al., 1988).
L’âge de la puberté varie en fonction de deux principaux facteurs : le niveau alimentaire et les facteurs génétiques. Il est de 6 mois à 1 an chez l’espèce bovine (Mc Donald, 1980; Diskin et al., 2003).
Chez les genisses de race locale Marocaine, l’âge à la puberté indiqué par la première ovulation varie de 13.6 mois (Al Mandri, 1986) à 21.5mois (Hosaini-Hilali, 1986). Indiqué par le premier œstrus, cet âge varie de 13.4 mois (Al Mandri, 1986) à 23.6mois
(Hosaini-Hilali, 1986). Il est de 17.8mois lorsqu’il est indiqué par le premier œstrus ovulatoire (Dewulf et Lahlou-Kassi, 1983). Quand plus de 50% des génisses sont cyclées, l’âge à la puberté est de 16.5 mois (Asri, 1984).
III/ Cycle sexuel
Le cycle sexuel peut être défini comme étant l’ensemble des modifications anatomiques (au niveau de l’ovaire), hormonales (Figure 4) et comportementales qui se succèdent entre deux œstrus (INRAP, 1988).
La physiologie du cycle sexuel est complexe et fait intervenir le niveau central : hypothalamus et hypophyse et l’appareil génital par les ovaires et l’utérus. L’ovaire règle à la fois sa propre production hormonale et la production ou le fonctionnement du tractus génital. L’activité sur le tractus genital se fait à la fois directement et via l’axe hypothalamo-hypophysaire (Figure 5).
1. Composante anatomique
Une fois formés, les ovaires sont assimilés à des glandes, situés dans la cavité abdominale. Chez les mammifères, c’est un organe pair, appendu à la région lombaire et pourvu d’une double fonction : gamètogène, assurant l’ovogenèse et endocrine, par la sécrétion des hormones œstrogènes et progestatives (Barone, 1987).
Chez les bovins, ces gonades sont oviformes, légèrement aplaties, de couleur blanc-rosé, leur grand axe est d’environ 1.5cm et elles présentent des bosselures plus ou moins apparentes à leur surface qui correspondent à des follicules à des stades avancés de leur évolution (Derivaux et Ectors, 1989).
1.1/ Vagues folliculaires
La régulation de la croissance folliculaire est complexe chez la vache.
En effet, à partir de la puberté, la croissance folliculaire est permanente, des vagues de croissance fertile et abortive (atrétique) se succèdent.
En effet, à partir du pool de follicules ovariens, 15 à 30 follicules entament leur croissance basale chaque jour et quittent donc la réserve. Au bout de 130 jours, certains atteignent le stade de follicules tertiaires (début de la croissance terminale) et vont rentrer dans le courant des vagues folliculaires.
Chaque vague consiste chez la vache en l’émergence, tous les 7 à 9 jours environ, de plusieurs follicules tertiaires antraux (5mm de diamètre au moins), parmi lesquels apparaît le follicule dominant (Ginther et al., 1989 ; Medan et al., 2005). Le follicule ovulatoire étant issu de la dernière vague (Hunter et al., 1988 ; Drion et al., 1996). Dans ce sens, de nombreuses études échographiques confirment la théorie des vagues selon laquelle le développement folliculaire évolue sous la forme de croissances et de régressions successives de plusieurs follicules (Sirois et Fortune, 1988 ; Knopf et al., 1989 ; Taylor et Rajamahendran, 1991).
Trois phénomènes se succédent : recrutement, sélection et dominance (Drion et al., 1996).
- Le recrutement est l’entrée en croissance terminale d’une cohorte de follicules gonadodépendants (diamètre : 2mm). Ces follicules ont dépassé le stade où habituellement la plupart des follicules deviennent atrétiques. Le recrutement, est un phénomène aléatoire, provoqué par l’augmentation transitoire du taux circulant de FSH (Follicle stimulating Hormone).
- La sélection est l’émergence du (ou des) follicule(s) ovulatoire(s) parmi les follicules recrutés. Cette sélection est secondaire à la réduction de FSH qui a initié le recrutement.
En effet, le développement du groupe de follicules recrutés s’accompagne d’une augmentation de la production d’œstradiol, mais également d’inhibine qui a un rétrocontrôle négatif sur l’hypophyse et diminue la production de FSH. A l’exception du (ou des) follicule(s) sélectionné(s) capable(s) de se développer en présence de faible taux de FSH, les autres follicules rentrent en atrésie.
- La dominance fait suite à la sélection. Elle est exercée par le plus gros follicule présent sur l’un ou l’autre ovaire. Le follicule dominant est le seul qui soit capable de provoquer la régression de follicules en croissance, ou d’inhiber la croissance d’autres follicules, et d’ovuler dans un environnement hormonal approprié. La dominance correspond au blocage du recrutement et à la croissance rapide du volume du (ou des) follicule(s) ovulatoire(s).
Bien que la FSH diminue, le follicule dominant persiste car il a acquis un mécanisme d’auto stimulation interne (l’œstradiol qu’il produit amplifie sa synthèse d’IGF-l (Insuline-like growth factor l) qui stimule sa synthèse d’œstradiol). La suite de l’évolution du follicule dominant dépend de l’évolution de la progestéronémie.
Si la progestéronémie diminue, c’est à dire s’il y a lutéolyse, alors que le follicule dominant est en phase de croissance, il ovule. Si, à l’inverse, la progestéronémie se maintient à un niveau élevé après que le follicule dominant ait atteint sa taille maximale, il commence sa régression, et une autre vague de croissance apparaît (Drion et al., 1996).
Ce schéma de croissance folliculaire est également décrit en période pré- pubertaire (Driancourt et al., 1991), en post partum et durant les 70 premiers jours de la gestation. En effet, malgré la présence d’un corps jaune, l’émergence de vagues de croissance folliculaire continue, mais sans phénomènes de sélection ni de dominance (Savio et al., 1990).
1.2/ Croissance et maturation de l’ovocyte
L’ovocyte est une cellule hautement spécialisée qui doit subir une longue différenciation le préparant à former avec le gamète mâle la première cellule souche d’un nouvel être. En l’occurrence, il doit subir une longue évolution cytologique et moléculaire durant toute la croissance folliculaire (Bevers et al., 1997).
1.2.1/ Croissance de l’ovocyte
Dès la naissance, le cortex ovarien contient un stock de follicules primordiaux hébergeant des ovocytes bloqués en fin de prophase I méiotique (stade vésicule
Stimulés par des mécanismes mal élucidés, quelques follicules primordiaux entrent dans une phase de croissance qui se traduit par la multiplication des cellules folliculaires mais surtout de la croissance de l’ovocyte (Gosden et al., 1997).
La croissance de l’ovocyte continue et lorsque l’antrum se forme, l’ovocyte atteint environ 80% de sa taille définitive (Mermillod et al., 1999a). Sa croissance continue de manière ralentie, tandis que le volume du follicule croît régulièrement jusqu’à la décharge gonadotrope ovulante. L’ovocyte reste bloqué en fin de prophase I méiotique pendant toute la croissance folliculaire. Il acquiert d’abord la compétence à reprendre sa méiose puis la compétence à assurer la fécondation et le développement (Eppig et al., 1996).
Durant cette croissance de l’ovocyte, une différenciation du cytoplasme a lieu parallèlement. En effet, les mitochondries s’accumulent dans le cytoplasme, au voisinage du réticulum endoplasmique rugueux, et migrent dans le cortex durant la phase finale de la maturation (Velilla et al., 2000). L’appareil de Golgi passe également par une phase d’activité intense, participant à la formation de la zone pellucide et des granules corticaux (Velilla et al., 2000). Les granules corticaux sont de petites vésicules (200-600 nm), en nombre de 5000 dans un ovocyte de souris en fin de croissance. Ils se positionnent contre la membrane plasmique dans la phase finale de la maturation de l’ovocyte et jouent un rôle capital dans la fécondation en empêchant la polyspermie (Guraya, 1983 ; Yoshida et al., 1990 ; Velilla et al., 2000).
Pendant la croissance, l’ovocyte accumule de grandes quantités d’ARN, augmentant d’environ 300 fois son contenu en ARN total. Les transcriptions, d’abord très intenses, diminuent lorsque l’ovocyte a atteint environ 80% de sa taille définitive. Elles deviennent indétectables dès la reprise de méiose ; les 2/3 environ sont des ARN ribosomaux, reflet de l’intense activité nucléaire observée pendant la période de croissance de l’ovocyte jusqu’à l’apparition de l’antrum (Mermillod, 2001).
La capacité de synthèse protéique de l’ovocyte de mammifères est d’abord élevée puis diminue au cours de la croissance. L’accumulation de certaines de ces protéines, très spécifiques de l’ovocyte, a été bien suivie. C’est le cas de trois glycoprotéines composant la zone pellucide (ZP1, ZP2 et ZP3), produites tout au long de la phase de croissance de l’ovocyte et représentant jusqu’à 10% des synthèses protéiques totales de l’ovocyte (Harrenstien et al., 2004).
La maturation ovocytaire recouvre l’ensemble des changements nucléaires et cytoplasmiques qui interviennent dans l’ovocyte I, à l’intérieur du follicule mûr suite à la décharge gonadotrope ovulante (décharge de LH) (Mermillod, 2001).
On distingue deux grands événements :
- La reprise de la méiose depuis le stade vésicule gérminative (VG) jusqu’en métaphase II ;
- la maturation cytoplasmique qui est l’acquisition de l’aptitude à une fécondation normale et au démarrage du développement.
a. Aspects cytologiques de la maturation des ovocytes a.1/ Rupture de la vésicule germinale
La rupture de la vésicule germinale se produit dans les heures qui suivent la décharge gonadotrope ovulante (Mermillod, 2001). Elle commence par un plissement de l’enveloppe nucléaire, en rapport avec le début de condensation de chromosomes, attachés à l’enveloppe par une extrémité. Les pores nucléaires disparaissent puis l’enveloppe se fragmente avant de disparaître elle même rapidement.
a.2/ Condensation des chromosomes
La condensation des chromosomes commence aux premiers signes d’ondulation de l’enveloppe nucléaire à laquelle ils sont attachés, se poursuit après la rupture de la vésicule germinative (Mermillod et Marchal, 1999b).
a.3/ Formation du fuseau chromosomique
Dans l’ovocyte, les centrioles disparaissent avant la croissance, dès le stade pachytène .Il ne subsiste que du matériel péricentriolaire filamenteux dans lequel s’établissent des MTOC (microtubules organizing center) formant les centrosomes de l’ovocyte. Les MTOC émettent des microtubules qui rejoignent les kinétochores des chromosomes puis migrent pour former les deux pôles d’un fuseau aplati, en forme de tonneau (Mermillod, 2001).
a.4/ Passage et arrêt en métaphase II
Chez la souris, la prométaphase (condensation des chromosomes et arrangement en plaque métaphasique) dure environ 6 heures et la métaphase I dure 4 heures. Cette durée importante est due sans doute au temps nécessaire à la synthèse et à l’assemblage du
temps qu’une boursouflure de la membrane qui deviendra le premier globule polaire. Ces deux phases sont rapides, les chromosomes homologues se séparent et migrent aux pôles du fuseau alors que le corps intermédiaire, une région riche en filaments et tubules participe à l’étranglement et finalement à l’expulsion du 1er globule polaire (Mermillod, 2001).
a.5/ Maturation cytoplasmique
La migration des granules corticaux (vésicules à contenu dense formées à partir de l’appareil de golgi dans les ovocytes des follicules pré ovulatoires) est le trait le plus marquant de la maturation cytoplasmique. Ces granules ont une localisation cytoplasmique sous corticale diffuse dans l’ovocyte immature et dès la reprise de méiose, migrent vers la zone corticale en s’associant au cytosquelette.
La migration des granules s’accompagne d’un rassemblement des autres organites (mitochondries et appareil de Golgie) dans la région périnucléaire, ces déplacements étant sous la dépendance des microtubules (Velilla et al., 2000).
La membrane des granules corticaux s’attache à la membrane plasmique par des ponts de calpactine (protéine impliquée dans l’exocytose) et la libération de leur contenu dans l’espace péri-vitellin au moment de la fécondation, ceci évite la polyspermie (Yoshida et
al., 1993).
La maturation cytoplasmique se traduit donc par l’acquisition d’un facteur de décondensation de la tête du spermatozoïde après fécondation (Marquant-Leguienne, 1986).
b. Aspects moléculaires de la maturation de l’ovocyte
Parmi les aspects moléculaires qu’on observe au cours de la maturation ovocytaire, il y a l’expansion du cumulus (Humblot et al., 2005), sur laquelle on peut se baser pour juger de la maturation des ovocytes.
En effet, comme en attestent les expériences de maturation ovocytaire in vitro, la présence de cellules de cumulus est importante pour la qualité de la maturation cytoplasmique (Salustri et al., 1996). Des ovocytes débarrassés de leurs cellules de cumulus et placés en culture sont capables d’accomplir leur méiose jusqu’en métaphase II. Cependant, leur fécondation est anormale et ils ne peuvent se développer.
Cette expansion est sous la dépendance de la FSH du fluide folliculaire dont l’action serait modulée par la LH.
In vitro, la FSH, provoque l’expansion du cumulus et pas la LH (Mermillod et Marchal, 1999b). In vivo, l’action de la FSH présente dans le fluide folliculaire avant le pic de LH pourrait être bloquée par un inhibiteur. La LH lèverait cette inhibition lors du pic ovulatoire (Cecconi et colonna, 1996).
L’expansion du cumulus pendant la maturation de l’ovocyte, résulte de l’accroissement des espaces intercellulaires suite à la production par les cellules d’une matrice visco-élastique abondante, composée d’acide hyaluronique (Mermillod et Marchal, 1999b). Cette matrice permet à l’ensemble cumulus-ovocyte de conserver son intégrité même après la dissociation des complexes de jonction. La synthèse d’acide hyaluronique par les cellules du cumulus est sous la dépendance de la FSH. Certains facteurs de croissance (EGF, IGF-I) agissent aussi sur l’expansion du cumulus.
En effet, des facteurs originaires de l’ovocyte interviennent dans ce phénomène, c’est notamment le cas du « cumulus expansion enabling factor » (CEEF) ou facteurs d’expansion du cumulus, molécule proche du TGF-β, produit du gène GDF-9 de l’ovocyte.
Le produit de ce gène GDF-9, capable de réguler plusieurs aspects du métabolisme des cellules somatiques du follicules (prolifération, synthèse des stéroïdes, d’acide hyaluronique et de prostaglandines) est une illustration du dialogue existant entre l’ovocyte et son environnement somatique (Elvin et al., 2000).
De leur part, Mermillod et Marchal, 1999b, ont rapporté que le rôle des cellules du cumulus peut s’exercer par l’apport de métabolites vers l’ovocyte (tel que le gluthation) et également par la transmission de signaux vers l’ovocyte. Ainsi, la stimulation de la maturation de l’ovocyte bovin par l’hormone de croissance passe par le cumulus (Izadyar et al., 1998).
D’un point de vu mécanique, l’expansion du cumulus est également importante puisqu’elle facilitera le détachement du complexe cumulus oophorus-ovocyte et sa capture par le pavillon lors de l’ovulation.
1.3/ Ovulation
Arrivé au terme de sa croissance le follicule forme à la surface de l’ovaire une saillie conique et libère l’ovocyte, en réponse à une forte élévation des gonadotropines ou décharge ovulante. L’ovulation se produit 11 à 12 heures après la décharge chez la lapine, 29 à 31 heures chez la vache et 35 à 36 heures chez la femme.
L’ovocyte libéré est accompagné de la corona radiata, d’une partie du cumulus oophorus et du liquide folliculaire.
Pendant le processus de l’ovulation, plusieurs changements structuraux et métaboliques se produisent et entraînant une désorganisation du follicule et de là sa rupture (Driancourt et al., 1991). C’est ainsi que :
• La thèque externe devient œdémateuse par diffusion du plasma sanguin.
• Les faisceaux de fibres de collagène de la thèque externe et de l’albuginée se dissocient.
• Les cellules de la granulosa se détachent de la lame basale, cessent de se diviser et perdent les jonctions perméables qui les unissent.
• Les cellules du cumulus subissent les mêmes transformations que les cellules de la granulosa mais comme elles secrètent abondamment de l’acide hyaluronique, leur dissociation est totale. L’ovocyte pilote cette sécrétion : le cumulus sans ovocyte ne produit plus d’acide hyaluronique.
• Les cellules péri ovocytaires, dont les prolongements traversent la zone pellucide, demeurent plus ou moins longtemps attachées à l’ovocyte, formant la corona radiata. Chez les ruminants cette corona disparaît rapidement, elle ne joue aucun rôle dans la fécondation.
• Peu avant la rupture du follicule, la lame basale séparant la granulosa de la thèque interne disparaît par endroits et des vaisseaux sanguins néoformés pénètrent dans la granulosa entraînant des cellules de cette thèque, préparant ainsi la formation du corps jaune (Driancourt et al., 2001b).
• Au niveau de l’apex de la saillie conique les changements sont différents : - les cellules de l’épithélium ovarien s’étirent et s’aplatissent
accompagnant l’extériorisation du follicule à la surface de l’ovaire.
- les cellules sous-jacentes de la granulosa, des thèques et de l’albuginée se dissocient puis disparaissent par apoptose et nécrose.
- La rupture folliculaire s’achève par la désintégration complète de l’apex. La pression intrafolliculaire diminue mais persiste entre la décharge de LH et l’ovulation semble faciliter la rupture. Par contre, l’expulsion de l’ovocyte et des cellules de la corona radiata résulte bien d’une contraction du follicule.
1.4/ Corps jaune
Après la rupture du follicule, les cellules de la granulosa, nouvellement vascularisées, s’hypertrophient et prolifèrent in situ pour former le corps jaune, alors que les cellules
de la thèque s’incorporent dans le tissu interstitiel de l’ovaire et participent à la sécrétion des androgènes ovariens (Hunter, 1980).
Chez la plupart des mammifères, le corps jaune formé après l’ovulation a une durée de vie limitée entre 4 et 21 jours si la femelle n’a pas été fécondée. Sa régression ou lutéolyse permet l’apparition d’un nouveau cycle ovulatoire. La gestation induit un blocage de la lutéolyse et la persistance du corps jaune cyclique en corps jaune gestatif dont la sécrétion de progestérone est indispensable à l’établissement de gestation (Leymarie et martal, 2001).
D’autre part, cette lutéinisation qui coïncide avec une augmentation très importante de la sécrétion de progestérone est accompagnée dans toutes les espèces, sauf les primates, de la disparition des sécrétions d’androgènes et d’œstrogène (Niswender et al., 2000).
2/ Composante hormonale
Le cycle d’activité de l’ovaire est en étroite relation avec les variations des profils hormonaux qui se produisent au niveau de l’ovaire d’une part et au niveau du système nerveux central et de l’axe hypotalamo-hypophysaire d’autre part.
Les sécrétions hormonales de l’hypothalamus, de l’hypophyse et de l’ovaire contrôlent la succession des événements du cycle (INRAP, 1988).
2.1/ Hormones hypothalamiques
GnRH ou Gonadotropine Releasing Hormone ou gonadolibérine est l’hormone de décharge ou encore l’hormone de libération (libérins) d’autres hormones (Gruyter, 1988).
Cette hormone est également nommée FSH-RH (Folliculo-Stimuline-Releasing Hormone) ou LH-RH (Luteinizing Hormone-Releasing Hormone) (Hafez, 1993).
C’est une hormone protidique responsable de la synthèse et de libération de deux hormones hypophysaires, les gonadotrophines FSH et LH (Bonnes et al, 1988). En effet, de nombreuses situations expérimentales visant à supprimer ou à limiter la sécrétion de la GnRH ont permis de montrer son importance sur la synthèse et la libération de FSH et LH (Filicori et al., 1994). La GnRH joue manifestement un rôle pivot dans l’initiation, la régulation et la suppression de la fonction reproductrice.
Elle a une sécrétion pulsatile, chaque pulse est formée de la somme de petites quantités de GnRH, libérées chacune par un neurone (Caraty et al., 2001). Le pulse peut être défini comme un épisode bref de libération hormonale dans le sang (Pelletier, 1983), il
est caractérisé par une montée rapide des concentrations sanguines suivie d’une diminution de type exponentiel liée à la demi-vie de l’hormone.
2.2/ Gonadotrophines hypophysaires (FSH et LH)
Au début de l’œstrus, se produit une décharge de gonadotropines qui entraîne l’ovulation, marquant la fin de la phase folliculaire et le début de la phase lutéale (Driancourt et Levasseur, 2001c ; Medan et al., 2005).
Les gonadotrophines jouent un rôle central dans la régulation de la fonction de la reproduction tant chez le mâle que chez la femelle ; elles sont en effet les intermédiaires essentiels du système nerveux central sur les activités endocrines et gamétogéniques des gonades (Medan et al., 2005).
La FSH et LH appartiennent à la famille des hormones glycoprotéiques à action directe et unique sur les gonades chez le mâle et la femelle ; ce sont des hormones gonadotropes ou gonadotrophines ou gonadostimulines : FSH (Follicule Stimulating Hormone) ou follitropine ou hormone folliculo-stimulante et LH (Luteinising Hormone) ou lutropine ou hormone lutéinisante (Bonnes et al., 1988).
La LH et la FSH confèrent à l’hypophyse une fonction de relaie amplificateur dans le contrôle de la fonction de reproduction :
- par le système nerveux central, principalement sous l’impulsion de la GnRH (Moenter et al., 1991 ; Bartolome et al., 2005).
- par des hormones périphériques et notamment les stéroïdes sexuels et l’inhibine, via la circulation générale.
- par divers facteurs produits localement par les cellules folliculaires comme l’inhibine, l’activine et IGF (facteurs de croissance) ainsi que leurs protéines de liaison telle que : la follistatine.
D’autre part, la FSH accompagne la croissance du follicule secondaire en follicule dominant dans les ovaires des mammifères et contrôle le développement des follicules, elle est l’hormone de la phase folliculaire précoce (Erikson et Danforth, 1995).
Chez les bovins, il ressort que la FSH, joue un rôle important dans l’initiation du développement folliculaire (Tanaka et al., 2001 ; Stenbak et al., 2001).
Elle stimule l’activation de l’aromatase et accélère la production des œstrogènes (Bao et
al., 1997).
En plus, la prolifération des cellules de la granulosa du follicule est induite par une action autocrine de l’œstrogène, et stimulée par la FSH (Peters et McNatty, 1980). La
sécrétion de FSH se produit par pics, mais d’une façon moins marquée, et est régulée par la sécrétion d’œstradiol et d’inhibine par les follicules.
Les principales fonctions de la LH sont la stimulation de la croissance folliculaire (Bartolome et al., 2005), la maturation finale du follicule dominant par la stimulation de la production d’œstradiol (Stock et Fortune, 1993), l’induction de l’ovulation et la stimulation de la sécrétion de progestérone par le corps jaune.
En effet, le pic de LH induit par l’effet conjugué d’une hypersensibilité
hypophysaire et d’une sécrétion de GnRH hypothalamique permet la reprise de la méiose par l’ovocyte, la rupture folliculaire et la lutéinisation des cellules de la granulosa (Bartolome et al., 2005).
2.3/ Hormones ovariennes
Ce sont la testostérone, les œstrogènes, la progestérone. De nature lipidique, elles sont fabriquées à partir du cholestérol et sont sécrétées principalement par les gonades mais aussi par le placenta et les glandes surrénales (Bonnes et al., 1988).
Etymologiquement, œstrogène signifie «qui engendre l’œstrus ». Sécrétés essentiellement par les follicules de l’ovaire, les œstrogènes ont pour rôle primordial de provoquer l’œstrus ou chaleurs.
L’augmentation du nombre des jeunes follicules antraux coïncide avec l’accumulation d’œstradiol dans l’antrum. L’œstradiol, stimule la prolifération des cellules de la granulosa et la formation de l’antrum (Peters et McNatty, 1980). L’effet lutéolytique de l’oestradiol a été rapporté par Colazo et al.,(2005) :Une perfusion d’oestradiol induit l’atrésie folliculaire suite à la baisse du taux circulant de FSH. A partir du moment où la concentration en oestradiol décline, un redressement du taux de FSH a lieu et une nouvelle vague folliculaire émerge 24 heures après.
La progestérone signifie « qui permet la gestation ». Sécrétée essentiellement par le corps jaune de l’ovaire, la progestérone est d’abord l’hormone responsable du maintien de la gestation (Graham et Clarke, 1997).
La progestérone, exerce un rétrocontrôle négatif sur la production de GnRH, FSH et LH.
2.4/ Autres hormones
a/ Les Prostaglandines
Ensemble de molécules de nature lipidique, synthétisées par de nombreuses cellules sécrétrices, les prostaglandines sont présentes dans presque tous les tissus de l’organisme des mammifères, elles exercent des rôles multiples, en général par action
Les prostaglandines permettant l’éclatement du follicule au moment de l’ovulation. Elles déclenchent la régression du corps jaune ou lutéolyse. Elles entretiennent les contractions du myomètre au moment de la mise bas. Les prostaglandines sont essentiellement d’origine utérine.
b/ Ocytocine
L’ocytocine, appelé aussi oxytocine, est un nanopeptide formé au niveau des noyaux supra-optiques et paraventriculaires de l’hypothalamus, et transporté puis stocké par la posthypophyse. Ses principales cibles sont l’utérus et les glandes mammaires. Elle intervient chez la femelle au moment de la mise bas et de l'éjection du lait.
2.5/ Régulations hormonales
a. Contrôle de la sécrétion de la GnRH
Chez les mammifères, l’initiateur et le régulateur fondamental de la fonction reproductrice est la GnRH (gonadotrophin releasing hormone ou gonadolibérine), qui est synthétisée et libérée par les neurones de l’hypothalamus. La GnRH se lie aux récepteurs spécifiques situés sur les cellules gonadotropes de l’antéhypophyse ce qui provoque la synthèse et la libération des gonadotropines, l’hormone folliculo-stimulante (FSH) et l’hormone lutéinisante (LH).
La FSH, à son tour agit spécifiquement sur les petits follicules ovariens pour stimuler leur croissance, tandis que la LH agit sur le follicule dominant mûr pour provoquer la maturation finale et l’ovulation qui en résulte (Drion et al., 1996).
La GnRH est secrétée par l’hypothalamus sous forme de décharges ou de manière épisodique, chaque décharge de GnRH provoquant la décharge de LH par l’antéhypophyse. La caractéristique fondamentale de la sécrétion des hormones hypothalamo-hypophysaires (GnRH, FSH, LH) est la pulsatilité; en d’autres termes, leur sécrétion n’est pas continue mais passe par un maximum très bref, pulsation ou « pulse », à la suite duquel la concentration plasmatique décroît progressivement jusqu'à une valeur minimale où elle stagne jusqu'à la prochaine élévation brève.
L’amplitude des pulsations est constante alors que leur fréquence varie, son augmentation provoquant alors un accroissement de la concentration de l’hormone dans le sang (Marie, 1996).
La sécrétion de GnRH est régulée par des facteurs internes et externes :
-Facteurs internes : Les principaux facteurs internes qui régulent la sécrétion de
progestérone agit sur les neurones de la GnRH pour diminuer la sécrétion de GnRH en abaissant la fréquence des décharges de GnRH, tandis que les effets de l’oestradiol sur la femelle dépendent de la présence ou de l’absence de concentrations de progestérone durant la phase lutéinique. Lors de la phase lutéinique où les concentrations de progestérone sont élevées, l’oestradiol agit en synergie avec la progestérone pour diminuer la sécrétion de GnRH par l’hypothalamus, c'est-à-dire qu’il y a une rétroaction négative sur la GnRH (Archbald et al., 1993). Lors de la phase folliculaire, en l’absence de progestérone et en présence de fortes concentrations de GnRH, l’oestradiol sécrété par le follicule pré-ovulatoire a une rétroaction positive sur la GnRH, ce qui provoque la prolongation d’une sécrétion élevée responsable des pics pré-ovulatoires de LH et de FSH (Drion et al., 1996).
-Facteurs externes : les principaux facteurs externes qui affectent la sécrétion de
GnRH sont : le statut nutritionnel de la vache (Armstrong et al., 2002) et le stimulus d’allaitement chez la vache allaitante. Le mécanisme selon lequel l’alimentation affecte la sécrétion de GnRH n’est pas éclairci mais le programme d’alimentation et l’état corporel réel de l’animal sont tous deux importants.
b. Contrôle de la sécrétion de LH et de FSH.
L’action de la GnRH sur l’antéhypophyse peut également être influencée par des hormones spécifiques produites par le follicule. La plus intéressante de toutes est l’inhibine. Cette hormone supprime sélectivement la libération de FSH par l’antéhypophyse sans affecter la sécrétion de LH (Mermillod et Marchal, 1999b).
L’activine stimule aussi la synthèse de FSH et l’équilibre entre ces deux facteurs peut déterminer le niveau de sécrétion de FSH.
La libération différentielle de LH et de FSH par la même cellule gonadotrope requiert des mécanismes de contrôle intracellulaires différents.
Les gonadotropines synthétisées sont stockées dans des granules sécrétoires à l’intérieur du cytoplasme, et sont secrétées par action différentielle par exocytose (Mermillod et Marchal, 1999b).
Il apparaît que le stockage de LH se prolonge durant le cycle oestral, mais le stockage de FSH est de courte durée.
Durant le cycle œstral de la brebis, jusqu’à 50% de FSH est libérée chaque jour, tandis que seulement 1 à 5% de LH est libérée. Par contre jusqu’à 70% de la LH totale est libérée durant la montée préovulatoire (Figure 3). Il est à présent évident que la LH est
étroitement régulée par la GnRH (Gazal et al., 1998), mais aussi par un contrôle local via les interactions de l’inhibine et de l’activine (Mermillod et Marchal, 1999b)
Chez la vache, les facteurs qui diminuent la fréquence des décharges de LH et empêchent par conséquent l’ovulation sont : le déficit énergétique en post partum ou pendant les périodes de sécheresse (Beam et Butler, 1999), une mauvaise nutrition, mauvais état corporel, perte supérieure à 10% du poids corporel après le vêlage et une durée d’allaitement supérieure à 6 semaines après le vêlage.
3/ Composante comportementale 3.1/ Saisonnalité
La plupart des mammifères présentent des cycles annuels de reproduction caractérisés par la succession d’une période d’activité sexuelle plus ou moins longue appelée saison sexuelle et d’une période de repos, résultant de la mise en silence de la fonction gonadotrope, appelée saison d’anœstrus ou anœstrus saisonnier (Martinet et Mondain-Monval, 1991).
Cependant, une vache post-pubérale non gestante bien nourrie sous des conditions environnementales ordinaires de domestication est vraiment un animal polyœstrien non saisonnier.
Bien que la vache n’ait pas une reproduction saisonnière, plusieurs études indiquent que la vache atteint une fertilité élevée au printemps et basse durant l’hiver et l’été (Mc Donald, 1980 ; DeRensis et Scaramuzzi, 2003).
En effet, les températures saisonnières élevées provoquent des changements endocriniens et une altération de la fonction endocrine : diminution de la sécrétion d’œstradiol, augmentation de la progesteronémie et perturbation des mécanismes de maturation ovocytaire et d’ovulation (Hochi et al., 1993 ; Berthelot et Bergonier, 1995 ; Rocha et al., 1998; DeRensis et Scaramuzzi, 2003).
D’autre part, l’équipe de Miettinen et al., (1991) a constaté que l’effet de la saison sur la fonction ovarienne passe par le pâturage et qu’une balance énergétique négative induite par un pâturage inadéquat qui affecte négativement les performances de reproduction. Dans le même sens et dans les conditions de reproduction in vitro, l’effet dépressif de l’été sur la qualité des ovocytes de la race Holstein est évident, il a été montré, que la proportion d’ovocytes et d’embryons arrivés au stade blastocyste au bout de huit jours étaient plus élevées en hiver qu’en été (AL -Katanani et al., 2002).
Le stress thermique altère le développement folliculaire en réduisant la production d’hormones stéroïdiennes (Wolfensen et al., 1997 ; Wilson et al., 1998). Ces