Rôle de DEPTOR dans la régulation du bilan d'énergie
Texte intégral
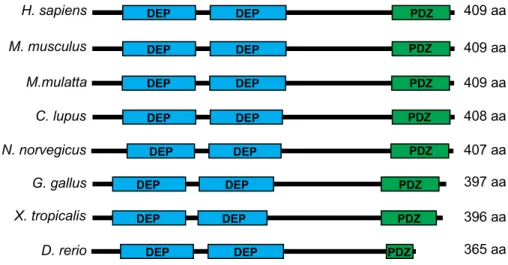
Figure

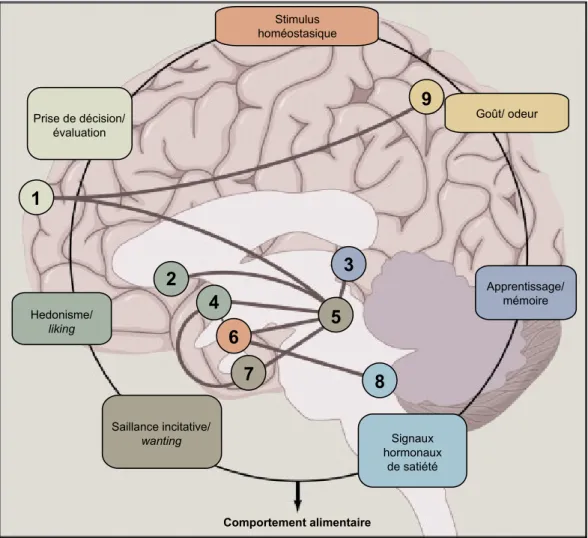

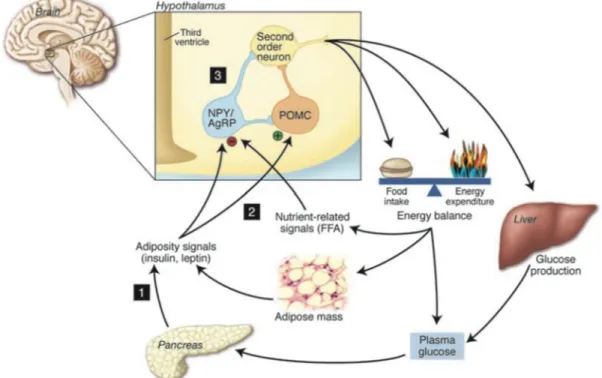



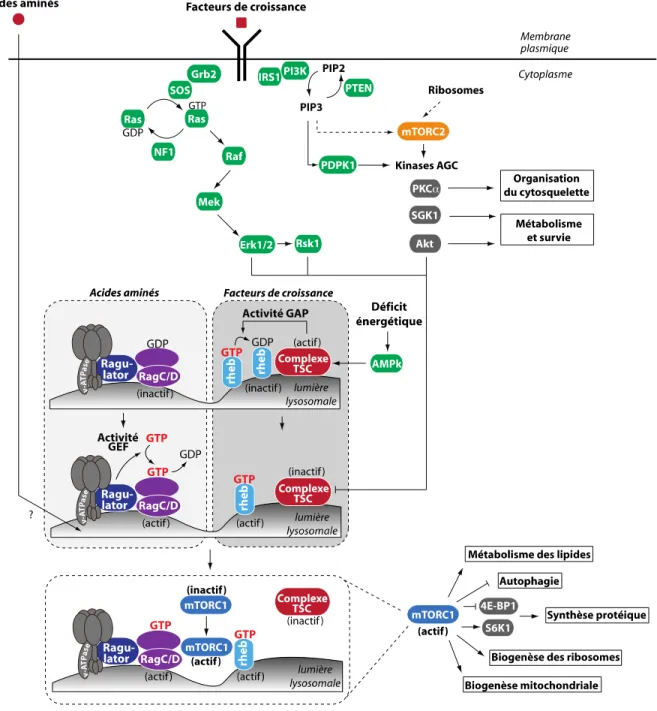
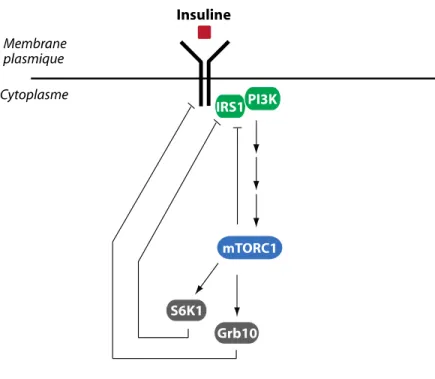
Documents relatifs
En effet, l’entrée de calcium par l’intermédiaire des récepteurs NMDA joue un rôle essentiel dans certains processus de plasticité synaptique comme la LTP (Regehr and Tank
Cette observation suggère que la protéine C/EBP pour rait être le médiateur de certains effets transcriptionnels de l' AMPc, d'autant plus que les deux protéines du
5 Le peuple décide en même temps si, en cas de vote affirmatif, la révision totale doit être faite p ar le G rand Conseil ou p ar une constituante.. Art. Le G rand Conseil
i) L‘approfondissement des connaissances quant à la composition chimique des huiles essentielles des espèces sélectionnées qui à notre connaissance ont été peu
Le Récepteur qui capte les changements (Stimuli) auxquels il faut apporter une réponse, et qui envoie l’information au niveau du centre de régulation, via la
Dans la revue officielle du Governatorato de Rome, Capitolium, on parle à l’époque de la campagne autour de Rome comme d'un territoire « atteint » par le paludisme, à assainir :
Mais aussi parce que, compte tenu du fondement marxien de cette théorie, la problé- matique du passage entre le micro et le macroéconomique n’est pas très naturelle pour la théorie
En mobilisant quantité d’études et de rapports produits sur la profession médicale ainsi que des matériaux de première main (articles de presse, littérature
