MARC-OLIVIER DUCEPPE
PROTÉOME ET BILAN PHOTOSYNTHÉTIQUE DE LA
POMME DE TERRE (SOLANUM TUBEROSUM L.) EN
RÉPONSE AU DORYPHORE DE LA POMME DE TERRE
(LEPTINOTARSA DECEMLINEATA SAY)
Thèse présentée
à la Faculté des études supérieures de l‟Université Laval dans le cadre du programme de doctorat en biologie végétale
pour l‟obtention du grade de Phisolophiae Doctor (Ph.D.)
DÉPARTEMENT DE PHYTOLOGIE
FACULTÉ DES SCIENCES DE L‟AGRICULTURE ET DE L‟ALIMENTATION UNIVERSITÉ LAVAL
QUÉBEC
2011
Avant-propos
Cette thèse de doctorat est présentée sous la forme d‟articles scientifiques et comprend 4 chapitres. Une introduction générale s'attarde, d'abord, à l‟importance économique de la pomme de terre, notre modèle d‟étude, et à l‟impact négatif des insectes défoliateurs sur le rendement de cette production agricole. Le Chapitre 1, intitulé « Revue de la littérature », dresse un survol des notions importantes et de l‟état des connaissances actuelles à propos des réponses biochimiques et physiologiques des plantes aux insectes défoliateurs; les hypothèses et objectifs de recherche du projet sont également décrits à la fin du chapitre.
Les Chapitres 2 à 4 présentent l‟ensemble des travaux effectués dans le cadre de la thèse. Ils sont rédigés en anglais, sous la forme d‟articles scientifiques standards. Chaque article est précédé d‟une description des objectifs poursuivis et d‟une synthèse des principaux résultats obtenus en cours de route. D'une manière plus précise, le Chapitre 2 porte sur l‟analyse protéomique des réponses de la pomme de terre au doryphore de la pomme de terre. Il intègre un article scientifique intitulé « Wounding, beetle leaf
feeding and aphid phloem feeding differentially alter the potato leaf proteome ». J'ai réalisé
l'essentiel des travaux décrits et rédigé le manuscrit présenté1. Le Prof. Conrad Cloutier et son équipe ont contribué aux aspects entomologiques de ce travail. Le Prof. Dominique Michaud a dirigé mes recherches et participé activement à la rédaction du manuscrit.
Le Chapitre 3 est une analyse protéomique des sécrétions orales du doryphore de la pomme de terre. Il intègre un article scientifique intitulé « Host plant proteins in the larval regurgitant of Colorado
potato beetle, Leptinotarsa decemlineata Say ». J'ai réalisé l'essentiel des travaux décrits et rédigé le
manuscrit présenté. Le Prof. Cloutier et son équipe ont contribué aux aspects entomologiques de ce travail. Le Prof. Michaud a dirigé les recherches et participé activement à la rédaction du manuscrit.
1 Une partie des travaux présentés dans ce chapitre ont été effectués dans le cadre de mes travaux de maîtrise,
Le Chapitre 4 porte sur l‟effet d'une défoliation par le doryphore de la pomme de terre sur les capacités photosynthétiques de sa plante-hôte, la pomme de terre. Il intègre un article scientifique intitulé « Insect regurgitant- and plant leaf-derived factors impact photosynthesis in potato plants interacting with the Colorado potato beetle ». J'ai réalisé l'essentiel des travaux décrits et rédigé le manuscrit présenté. Le Prof. Cloutier et son équipe ont contribué aux aspects entomologiques de ce travail. Le Prof. Yves Desjardins m‟a apporté une aide technique et logistique précieuse pour les mesures de photosynthèse et m'a secondé pour l'interprétation des données. Comme pour les articles précédents, le Prof. Michaud a dirigé mes recherches et participé activement à la rédaction du manuscrit.
Pour compléter la thèse, une Discussion et une Conclusion générales sont finalement présentées, avec pour but de mettre en perspective les principaux résultats obtenus et de proposer des travaux futurs. Une annexe résumant quelques notions de base sur les mesures de photosynthèse dans un contexte d‟étude des stress chez les plantes est également incluse à la fin de la thèse.
J‟aimerais remercier en tout premier lieu mon directeur de thèse, le Dr Dominique Michaud, pour m‟avoir permis de réaliser mon doctorat au sein de son laboratoire. Merci Dominique pour ton soutien financier, professionnel et moral. Mon passage dans ton laboratoire fut une expérience plaisante et très enrichissante. Tu as su me guider durant ce long parcours, tout particulièrement durant la rédaction de ce manuscrit. Ton esprit de synthèse inouï et tes conseils judicieux ont eu raison de la complexité de ce projet.
J‟adresse également mes remerciements les plus sincères à mon codirecteur de thèse, le Dr Conrad Cloutier, avec qui j‟ai aussi eu l‟honneur de travailler durant ma maîtrise. Merci Conrad pour tes précieux conseils. Ta rigueur scientifique est sans reproche et elle a certes contribué à mener ce projet à bon terme. Je tiens également à remercier le Dr Yves Desjardins. Yves, ton expertise et tes conseils éclairés concernant l‟aspect physiologique de ce projet m‟ont été d‟une grande valeur.
Merci à tous ceux qui ont contribué de près ou de loin à l‟avancement de mon doctorat. Je pense en particulier au personnel du CRH, au personnel de l‟INAF, à toute l‟équipe du laboratoire du Dr Michaud et plus particulièrement M. Simon Boudreault, pour son aide inestimable avec les insectes utilisés durant ce projet.
Je ne peux passer sous silence le Réseau Biocontrôle, qui, en plus de m‟avoir fourni un soutien financier et technique, m‟a permis de rencontrer des gens magnifiques que je n‟oublierai jamais. Je remercie aussi le Conseil de recherches en sciences naturelles et en génie du Canada (CRSNG), pour son soutien financier essentiel.
En terminant, merci à tous ceux qui me sont si chers, vous m‟avez donné l‟énergie nécessaire aux moments opportuns.
Les plantes ont développé, au cours du temps, des mécanismes de protection leur permettant de survivre et de se développer en dépit des nombreux stress biotiques auxquels elles sont soumises. Cette thèse doctorale, en trois volets, visait à caractériser les réponses biochimiques et physiologiques de la pomme de terre (Solanum tuberosum) attaquée par un insecte coléoptère, le doryphore de la pomme de terre (Leptinotarsa decemlineata). Le premier volet du projet visait à caractériser l'impact de l'insecte sur le protéome foliaire de la plante, en utilisant comme modèle des plantes traitées avec des larves de doryphore, des plantes soumises à des blessures mécaniques et des plantes infestées par un insecte suceur, le puceron de la pomme de terre (Macrosiphum euphorbiae). Le second volet visait à caractériser le protéome des sécrétions orales du doryphore, avec pour objectif de cerner l'incidence relative des protéines de l'insecte et de la plante hôte aux sites de blessure générés par l'herbivore. Le troisième et dernier volet visait, enfin, à déterminer l'impact des altérations du protéome foliaire de la plante sur ses capacités photosynthétiques, et à mettre en évidence l'impact possible des composantes moléculaires de la plante sur les réponses observées. En résumé, nos résultats ont démontré que plusieurs protéines des métabolismes primaire et secondaire, incluant des protéines associées à la photosynthèse, sont régulées dans les feuilles en réponse au doryphore de la pomme de terre. L'impact négatif de l'herbivore sur plusieurs protéines photosynthétiques, notamment celles du photosystème I, a toutefois des répercussions négligeables sur la capture de lumière par la plante. La seconde phase du processus photosynthétique, en revanche, est affectée de manière notable par le doryphore, vraisemblablement par l'action de molécules provenant aussi bien de l'insecte que de la plante elle-même. Ces résultats suggèrent, dans l'ensemble, un impact spécifique, mais limité du doryphore de la pomme de terre sur le protéome primaire et les fonctions photosynthétiques de sa plante hôte. Ils suggèrent aussi la mise en place de mécanismes compensatoires in planta et la grande plasticité du métabolisme primaire de la plante en réponse à l'herbivore.
Higher plants have developed, over time, a variety of protection mechanisms allowing them to survive and cope with a variety of biotic stress cues in their surrounding environment. The main goal of this three-part doctoral thesis was to characterize the biochemical and the physiological responses of potato (Solanum tuberosum) to defoliation by the coleopteran insect herbivore Colorado potato beetle (Leptinotarsa decemlineata). The first objective of the project was to characterize the impact of the insect on the host plant's leaf proteome, using as a model plants treated with potato beetle larvae, mechanically wounded plants and plants infested with a sucking/piercing insect, the potato aphid (Macrosiphum euphorbiae). The second objective was to gain some insight about the proteome of potato beetle oral secretions, with the aim of assessing the relative incidence of insect and host plant proteins at wound sites generated during insect feeding. The third objective, finally, was to determine the impact of leaf proteome alterations on photosynthetic capacities of the host plant, and to determine the possible impact of the plant's own molecular constituents on the responses observed. In brief, our results showed that several proteins involved in the primary and the secondary metabolisms, including photosynthesis-related proteins, were regulated in leaves in response to potato beetle feeding. However, the negative impact of the insect on several photosynthetic proteins, notably photosystem I proteins, only had negligible effects on the light capture process by the plant. The second phase of photosynthesis, on the other hand, was significantly affected by the insect, presumably via molecular effectors from both the insect and the host plant itself. These findings suggest, overall, a specific, but somewhat limited impact of Colorado potato beetle larvae on the leaf proteome and photosynthetic capacities of the potato host. They also suggest the possible induction of compensatory mechanisms in planta and the high plasticity of primary metabolism functions in the plant upon herbivore feeding.
Avant-propos... ii
Remerciements ... iv
Résumé ... vii
Abstract ... viii
Table des matières... ix
Liste des figures ... xiii
Liste des tableaux ... xv
Liste des abréviations ... xvii
Préface... 1
Chapitre 1 Revue de la littérature ... 3
1.1 La réponse biochimique des plantes aux insectes défoliateurs ... 5
1.1.1 Une description du stress « insecte défoliateur » ... 5
1.1.1.1 La composante mécanique ... 7
1.1.1.2 La composante biochimique ... 11
1.1.2 En résumé... 22
1.2 La réponse physiologique des plantes aux insectes défoliateurs ... 23
1.2.1 Les effets indirects des insectes herbivores sur la photosynthèse... 23
1.2.1.1 L‟impact du mode d‟alimentation ... 24
1.2.1.2 L‟impact de la réponse de la plante ... 25
1.2.2 Les techniques d‟imagerie à haute résolution ... 28
1.2.3 En résumé... 30
1.3 Description du projet de recherche ... 31
Chapitre 2 Protéome foliaire de la pomme de terre en réponse au doryphore de la pomme de terre ... 34
2.1 Résumé ... 35
Wounding, beetle leaf feeding and aphid phloem feeding differentially alter the potato leaf proteome ... 37
2.3 Introduction ... 39
2.4 Results ... 42
2.4.1 Differential gene-inducing effects among treatments ... 42
2.4.2 Control, wounded and insect-treated leaves exhibit distinct proteome patterns ... 43
2.4.3 Potato beetles and aphids differentially impact photosynthesis-related proteins in leaves... 47
2.5 Discussion ... 53
2.6 Materials and methods ... 57
2.6.1 Plants ... 57
2.6.2 Stress treatments ... 58
2.6.3 Northern blotting ... 58
2.6.4 Real-time RT-PCR ... 59
2.6.5 Sample preparation for 2-DE ... 61
2.6.6 2-DE ... 61
2.6.7 Gel image analysis ... 62
2.6.8 MS analyses ... 62
2.6.9 Protein identification ... 63
2.7 Acknowledgments... 64
Chapitre 3 Caractérisation protéomique des sécrétions orales du doryphore de la pomme de terre ... 67
3.1 Résumé ... 68
Host plant proteins in the larval regurgitant of Colorado potato beetle, Leptinotarsa decemlineata Say ... 71
3.2 Abstract ... 72
3.3 Introduction ... 73
3.4 Results and discussion ... 75
3.4.1 Salivary glands of Colorado potato beetle larvae ... 75
3.4.2 Insect and plant proteins in the regurgitant of potato beetle larvae ... 76
3.4.3 Insect digestive proteases and plant defense proteins in the potato beetle regurgitant 80 3.5 Materials and methods ... 82
3.5.2 Sample collection ... 82
3.5.3 Sample preparation for 2-DE ... 83
3.5.4 2-DE ... 83
3.5.5 Gel image analysis ... 84
3.5.6 MS analyses and protein identification ... 84
3.6 Acknowledgments... 85
Chapitre 4 Efficacité photosynthétique de la pomme de terre en réponse au doryphore de la pomme de terre ... 89
4.1 Résumé ... 90
Insect regurgitant- and plant leaf-derived factors impact photosynthesis in potato plants interacting with the Colorado potato beetle ... 95
4.2 Abstract ... 96
4.3 Introduction ... 97
4.4 Results ... 99
4.4.1 Photosynthesis-related proteins are differentially regulated in wounded and insect-treated leaves ... 99
4.4.2 Wounding and potato beetle herbivory differentially impact photosynthesis in potato leaves... 105
4.5 Discussion ... 108
4.6 Materials and methods ... 112
4.6.1 Plants ... 112
4.6.2 Stress treatments ... 112
4.6.3 Leaf total proteins ... 113
4.6.4 Immunodetections ... 113
4.6.5 Photosynthetic parameters ... 114
4.7 Acknowledgments... 115
Discussion générale et conclusion ... 122
Références ... 130
Annexe I Notions de base sur les mesures de photosynthèse ... 142
La photosynthèse ... 143
Les courbes de réponse au CO2 ... 151
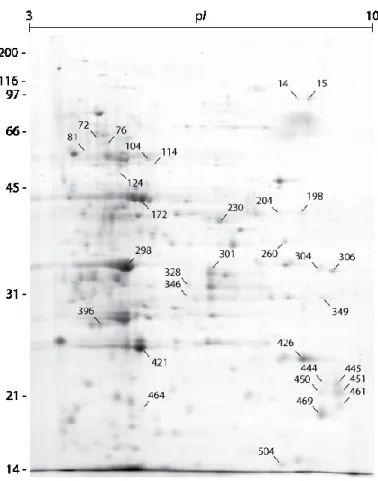
Figure 1.1 Modélisation schématique de l‟impact d‟un insecte phytophage de type défoliateur. . 6 Figure 1.2 Classes d‟éliciteurs identifiés dans les sécrétions orales des insectes herbivores. ... 17 Figure 1.3 Effet spatial de la défoliation sur l‟efficacité photosynthétique. ... 30 Figure 2.1 Northern blot analysis for the induction of Pin-II and protein P4 mRNA transcripts by
wounding (W), potato aphid piercing-sucking (A) and Colorado potato beetle leaf chewing (CPB) in potato leaves. ... 43
Figure 2.2 Gel image for the proteome of potato leaves subjected to potato aphid feeding, as
visualized after Coomassie blue staining following 2-DE. ... 44
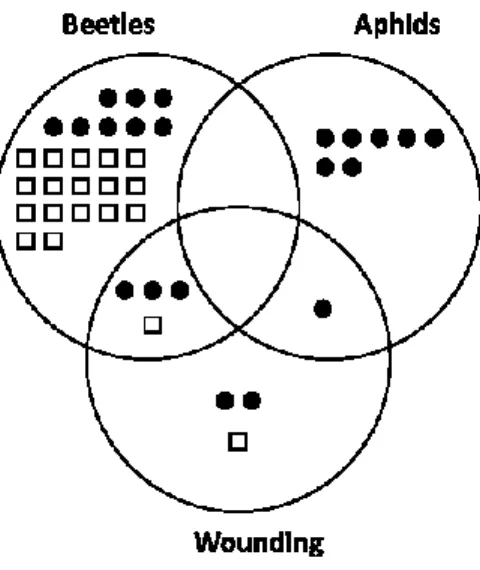
Figure 2.3 Venn diagram illustrating the relative number of proteins specifically or co-regulated
in potato leaves following mechanical wounding, Colorado potato beetle leaf chewing or potato aphid feeding. ... 46
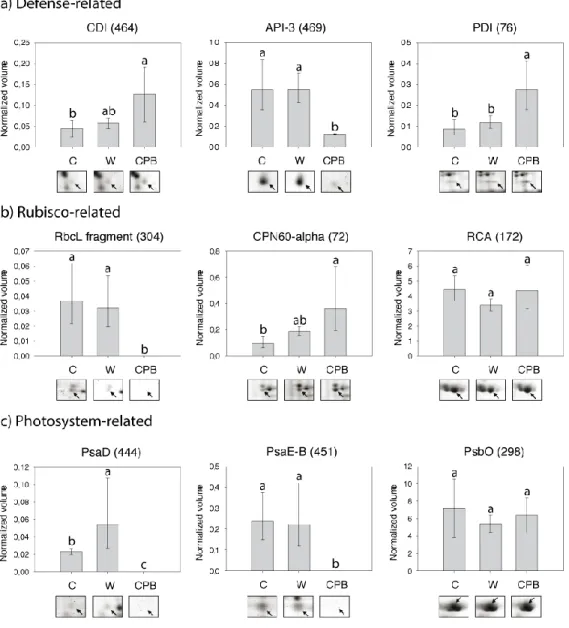
Figure 2.4 Selected defense- (A) and photosynthesis- (B, C) related proteins identified as
modulated in potato leaves subjected to wounding (W) or Colorado potato beetle (CPB) feeding... 49
Figure 2.5 Abundance of mRNA transcripts for photosynthesis-related proteins in potato leaves
subjected to wounding (W) or Colorado potato beetle (CPB) feeding. ... 52
Figure 3.1 Optical microscope images for the maxillary glands of Colorado potato beetle 4th -instar larvae. ... 76
Figure 3.2 Representative 2-D gels for potato leaf proteome and the proteome of larval Colorado
potato beetle regurgitant and maxillary glands. ... 78
Figure 4.1 Immunodetection patterns of photosynthesis-related proteins in control, healthy
potato plants. ... 101
Figure 4.2 Effects of stress treatment and leaf age on steady-state levels of AtpB and PsbA in
potato leaves... 104
Figure 4.3 Effects of stress treatment on Vcmax and TPU photosynthetic parameters in potato
Figure A.1 Niveaux d‟excitation de la chlorophylle en fonction de la longueur d‟onde des
photons incidents et voies de dissipation de l‟énergie. ... 143
Figure A.2 Paramètres de fluorescence, tels que mesurés par le LI-6400. ... 146 Figure A.3 Paramètres dérivés d‟une courbe de réponse lumineuse. ... 147 Figure A.4 Comportement de la courbe de réponse lumineuse en fonction de l‟état
physiologique de la plante. ... 149
Figure A.5 Vue schématique du cycle de Calvin. ... 153 Figure A.6 Paramètres dérivés des courbes ACi. ... 154
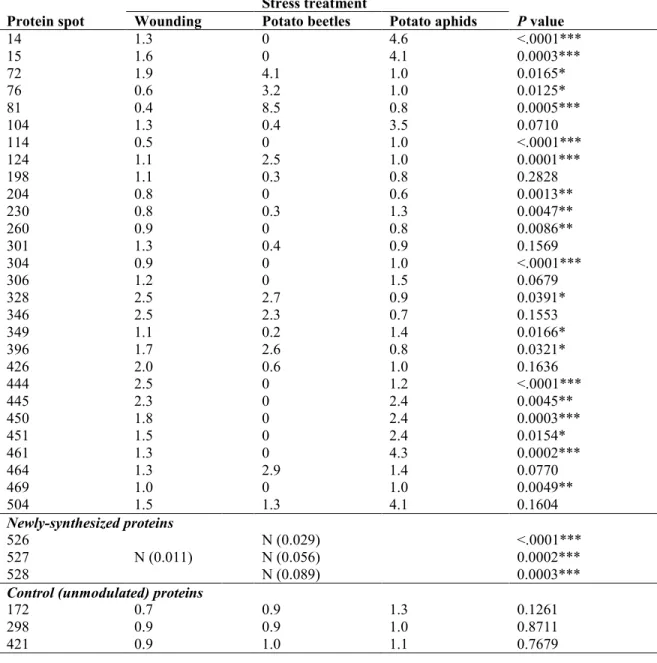
Table 2.1 Relative levels of potato leaf protein spots exhibiting a more than two-fold decrease or
increase for at least one treatment, compared to healthy control leaves (see Figure 2.2 for
protein numbering)... 45
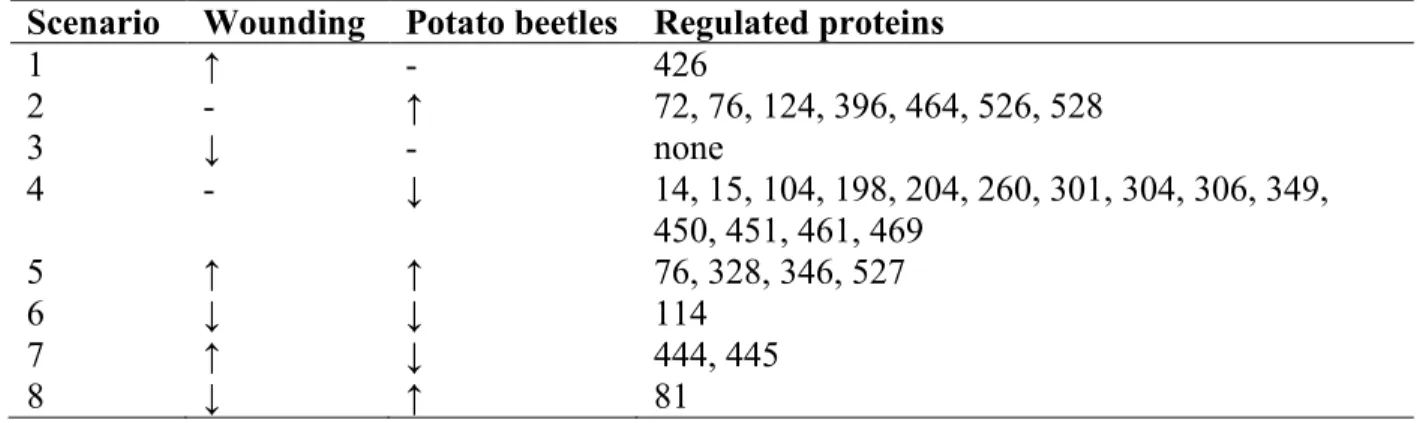
Table 2.2 Possible expression scenarios (excluding no response to either treatment) for stress-regulated proteins in potato leaves subjected to mechanical wounding or potato beetle chewing. ... 47
Table 2.3 Identity of stress-regulated proteins in potato leaves and their expression trend for three different stress treatments1. ... 50
Table 2.4 Oligonucleotide primer sequences for real-time RT-PCR. ... 60
Table 3.1 Insect and plant protein spots identified by MS (see Figure 3.2 for spot naming). ... 79
Table 4.1 Photosynthesis-related proteins monitored by quantitative immunodetection1. ... 100
Table 4.2 Effects of leaf age/position and treatment on photosynthesis-related proteins in potato leaves... 103
Table 4.3 Variation in the number of AtpB and PsbO bands detected on immunoblots. ... 104
Table 4.4 Effects of time and treatment on photosynthetic parameters derived from light response curves. ... 106
Table 4.5 Effects of time and treatment on fluorescence parameters inferred from data collected with a LI-6400 system equipped with a fluorescence chamber. ... 106
Table 4.6 Effects of time and treatment on photosynthetic parameters derived from ACi curves. ... 107
Supplementary Table 2.1 MALDI-TOF MS identification of potato leaf proteins regulated by wounding, potato beetle feeding or aphid phloem feeding. ... 65
Supplementary Table 2.2 LC-MS/MS identification of potato leaf proteins regulated by wounding, potato beetle feeding or aphid phloem feeding. ... 66
Supplementary Table 3.1 SELDI-TOF MS identification of selected proteins from potato leaves, and from larval Colorado beetle salivary glands and regurgitant. ... 86
Supplementary Table 3.2 LC-MS/MS identification of selected proteins from potato beetle
salivary glands, potato beetle regurgitant and potato leaves. ... 88
Supplementary Table 4.1 “Treatment x time interaction” mean values (±SD) of potato
photosynthetic parameters derived from light response curves (n=3). ... 116
Supplementary Table 4.2 “Treatment effect” mean values (±SD) of potato photosynthetic
parameters derived from light response curves (n=9)... 117
Supplementary Table 4.3 “Time effect” mean values (±SD) of potato photosynthetic parameters
derived from light response curves (n=15). ... 117
Supplementary Table 4.4 “Treatment x time interaction” mean values (±SD) of potato
photosynthetic parameters derived from fluorescence measurements (n=3). ... 118
Supplementary Table 4.5 “Treatment effect” mean values (±SD) of photosynthetic parameters
derived from fluorescence measurements (n=9). ... 119
Supplementary Table 4.6 “Time effect” mean values (±SD) of photosynthetic parameters
derived from fluorescence measurements (n=15). ... 119
Supplementary Table 4.7 “Treatment x time interaction” mean values (±SD) of potato
photosynthetic parameters derived from CO2 response curves (n=3). ... 120
Supplementary Table 4.8 “Treatment effect” mean values (±SD) of potato photosynthetic
parameters derived from the CO2 response curves (n=9). ... 121
Supplementary Table 4.9 “Time effect” mean values (±SD) of potato photosynthetic parameters
derived from CO2 response curves (n=15)... 121
Tableau A.1 Paramètres photosynthétiques mesurés pour déterminer l‟état physiologique d‟une
plante soumise à différents traitements. ... 145
Tableau A.2 Impact de la variation de la pente et du taux d‟assimilation maximal (Amax) de la
courbe de réponse lumineuse sur les différents paramètres de fluorescence mesurés et dérivés. ... 151
°C degré Celsius
1,3-PG 1,3-biphosphoglycérate
1-D unidimensionnel
2-D bidimensionnel
2-DE électrophorèse bidimensionnelle sur gel (two-dimensional gel electrophoresis) 2-HOT acide2-hydroxyoctadécatriénoïque
3-PG 3-phosphoglycérate
32P phosphore 32 (radio-isotope)
A taux d‟assimilation du CO2 (µmol CO2 m-2 s-1)
ADN acide désoxyribonucléique ADP adénosine diphosphate
AJ acide jasmonique
Amax taux maximal de photosynthèse (µmol CO2 m-2 s-1)
Aobs taux de photosynthèse mesuré (µmol CO2 m-2 s-1)
ARN acide ribonucléique AS acide salicylique ATP adénosine triphosphate
ATPase adénosine triphosphate synthétase (ou ATP synthase) AtpB sous-unité β du complexe ATPase
BM blessure mécanique
bp paire de bases (base pair) Calc. calculé
Cc pression partielle de CO2 à l‟intérieur des chloroplastes (Pa)
cDNA ADN complémentaire
CHAPS 3-[(3-Cholamidopropyl)dimethylammonio]-1-propanesulfonate Ci concentration intercellulaire de CO2 (µmol CO2 mol air-1)
CO2 dioxyde de carbone
cv. cultivar
Cys cystéine
Da Dalton
DNase désoxyribonucléase
EDTA acide éthylènediaminetétracétique (ethylenediaminetetraacetic acid) EST étiquette de séquence transcrite (expressed sequence tag)
ETR taux de transport des électrons du PSII (Electron transport rate) Exp. valeur expérimentale
FAC conjugué d‟acide gras (fatty acid – amino acid conjugate)
FAO Organisation des Nations Unies pour l‟Agriculture et l‟Alimentation (Food and
Agriculture Organization of the United Nations) Fo fluorescence minimale
Fv/Fm efficacité photochimique maximale
Fv‟/Fm‟ efficacité photochimique du centre réactionnel oxydé du PSII, pour une feuille adaptée à la
lumière
g force gravitationnelle
G3P 3-phosphoglycéraldéhyde (glyceraldehyde 3-phosphate)
gm conductance du mésophylle (µmol m-2 s-1 Pa-1)
GOX glucose oxydase
H2O2 peroxyde d‟hydrogène
H3PO4 acide phosphorique
HCl chlorure d‟hydrogène
IEF focalisation isoélectrique (isoelectric focusing) IPG gradient de pH stable (immobilized pH gradient)
J taux de transport d‟électrons du PSII servant à la réduction du NADP+ (µmol e- m-2 s-1)
L. Linné, Carl von
L:D ratio diurne : nocturne de la photopériode (light:dark) LC chromatographie en phase liquide (liquid chromatography)
m/z rapport masse / charge
MALDI matrix-assisted laser desorption/ionisation
MM masse moléculaire
MOWSE MOlecular Weight SEarch
MS spectrométrie de masse (mass spectrometry) MS/MS spectrométrie de masse en tandem
mRNA acide ribonucléique messager
NADPH forme réduite du nicotinamide adénine dinucléotide phosphate NADP+ forme oxydée du nicotinamide adénine dinucléotide phosphate NCBI National Center for Biotechnology Information
NH4HCO3 bicarbonate d‟ammonium
NPQ dissipation non photochimique de la fluorescence (non photochemical quenching)
P Probabilité (P value)
P-R relié à la pathogenèse (Pathogenesis-related)
PAGE électrophorèse sur gel de polyacrylamide (polyacrylamide gel electrophoresis)
PCR réaction en chaîne par polymérase (polymerase chain reaction)
pH potentiel hydrogène Pi phosphore inorganique
pI point isoélectrique
PsaC sous-unité VII du photosystème I PsaD sous-unité II du photosystème I PsaE sous-unité IV du photosystème I PsbA sous-unité D1 du photosystème II
PsbO sous-unité de 33 kDa du « complexe de scission de l‟eau » du photosystème II
PSI photosystème I
PSII photosystème II
pv. pathovar
qN dissipation non photochimique de la fluorescence
qP dissipation photochimique de la fluorescence
RbcL grande sous-unité de la rubisco RbcS petite sous-unité de la rubisco
Rd taux de respiration diurne (µmol CO2 m-2 s-1)
RT transcriptase inverse (reverse transcriptase) rubisco ribulose-1,5-bisphosphate carboxylase/oxygénase RuBP ribulose-1,5-bisphosphate
RuBP_regen phase de la photosynthèse où du RuBP est régénéré
SD écart-type (standard deviation)
SDS dodécylsulfate de sodium (sodium dodecyl sulfate)
SE erreur-type (standard error)
SELDI surface-enhanced laser desorption/ionisation
SGN Solanaceae Genomic Network
TFA acide trifluoroacétique (trifuoroacetic acid) TOF temps de vol (time-of-flight)
TPU utilisation des trioses phosphates (triphosphate use) (µmol m-2 s-1)
V volt
v/v volume / volume
Vcmax vélocité maximale de la carboxylation permise par la rubisco (µmol CO2 m-2 s-1)
Vh volt heure
W blessures mécaniques (wounding)
W+P blessures mécaniques additionnées d‟extrait végétal (wounding+plant extract) W+R blessures mécaniques additionnées de régurgitant (wounding+regurgitant) w/v poids/volume (weight/volume)
φPSII efficacité photochimique du PSII
↓ diminution
↑ augmentation
La pomme de terre, Solanum tuberosum L., fait partie de la famille des Solanacées, qui inclut aussi le tabac, la tomate et l‟aubergine. Elle est cultivée à travers le monde pour la valeur nutritive de son tubercule, qui est riche en amidon, en vitamine C et en potassium (Gagnon et al. 2007).
En 2006, au Québec, la production de la pomme de terre s‟étendait sur plus de 19 000 hectares (ha). Sa culture a engendré des revenus de 122 M$, ce qui en faisait la première culture maraîchère d‟importance dans la province. À l‟échelle canadienne, c‟est plus de 160 000 ha de pomme de terre qui ont été cultivés (Gagnon et al. 2007), pour une production de près de 5 M de tonnes métriques (tm) et un revenu total de 900 M$.
La culture de la pomme de terre est aussi très importante mondialement. En 2006, l‟Organisation des Nations Unies pour l‟Agriculture et l‟Alimentation (FAO) rapportait une production mondiale de près de 314 M tm (Gagnon et al. 2007). Les principaux pays producteurs de cette culture étaient, en ordre décroissant, la Chine (73,5 M tm, pour 23% de la production mondiale), la Fédération de Russie (38,6 M tm), l‟Inde (23,9 M tm), les États-Unis (19,7 M tm), l‟Ukraine (19,5 M tm) et l‟Allemagne (10 M tm). La pomme de terre est si importante mondialement que l‟année 2008 fut proclamée Année internationale de la pomme de terre par les Nations Unies.
Comme toute plante agricole, les rendements en pommes de terre sont affectés par plusieurs insectes et maladies. Le doryphore de la pomme de terre, Leptinotarsa decemlineata Say, est de loin l‟insecte qui cause le plus de pertes économiques pour cette culture. Il est présent partout où on la cultive en Amérique du Nord et en Europe de l‟Est. Sans contrôle phytosanitaire, le doryphore peut consommer la totalité des surfaces photosynthétiques des plantes, avec des conséquences majeures sur les rendements en tubercules (Noronha et al. 2008).
L‟application de pesticides de synthèse représente le principal moyen de lutte contre le doryphore en Amérique du Nord. Avec le temps, les populations visées ont développé une résistance à la plupart des pesticides homologués. Plusieurs recherches impliquant des instances gouvernementales, publiques et privées sont en cours dans le but de développer de nouvelles stratégies de lutte contre cet insecte. La recherche fondamentale et la recherche appliquée sont essentielles dans le développement de moyens de lutte intégrée efficaces. La recherche fondamentale, en particulier, permet d‟identifier, à plus ou moins long terme, des éléments clés pouvant mener à de nouvelles stratégies de lutte efficaces contre les ravageurs des cultures.
Au cours de ce projet de recherche, nous avons étudié certains aspects moléculaires, biochimiques et physiologiques de l‟interaction « pomme de terre-doryphore », en nous attardant principalement au côté « végétal » de l‟interaction. Notre décision de travailler avec la pomme de terre comme modèle d‟étude, outre son importance économique, repose sur le fait qu‟elle fait partie des Solanacées, une des familles taxonomiques les plus étudiées dans le domaine des stress biotiques et abiotiques chez les plantes. Les récents travaux réalisés par le Dr Ian Baldwin et ses collaborateurs, au Max Planck Institute for Chemical Ecology en Allemagne, représentent un exemple éloquent de l‟importance des Solanacées dans l‟avancement des nos connaissances sur les interactions plantes-insectes. Ce groupe de recherche a notamment publié une série d‟articles scientifiques sur la réponse du tabac sauvage (Nicotiania attenuata) à son herbivore spécialiste, le lépidoptère Manduca sexta (Halitschke et al. 2001; Hermsmeier et al. 2001; Schittko et al. 2001; Winz and Baldwin 2001; Halitschke et al. 2003; Hui et al. 2003; Giri et al. 2006; Gaquerel et al. 2009), qui a changé notre vision de la réponse des plantes aux stress biotiques. Les recherches de ce groupe innovaient du fait qu‟elles étudiaient les réponses de la plante hôte à l‟aide d‟une approche globale basée sur l‟utilisation de plusieurs méthodes d‟analyses complémentaires, une approche que nous avons préconisée durant le présent projet de recherche. Les travaux du Dr Baldwin ont entre autres montré que les deux partis impliqués dans l‟interaction, la plante et l‟insecte, avaient, au cours d‟une longue période de coévolution, acquis de nombreuses armes défensives et offensives afin d‟augmenter leur valeur adaptative (fitness) par une gamme de mécanismes de tolérance, de résistance, de compensation et d‟évasion.
Les plantes vasculaires, qui sont sessiles, possèdent un système de défense sophistiqué pour contrer l‟effet néfaste des différents stress auxquels elles sont soumises. Leur arsenal comprend, en bref, des moyens de lutte physiques et biochimiques qui peuvent être constitutifs ou induits (Kessler and Baldwin 2002).
Les défenses induites impliquent une importante dépense énergétique qui n‟est pas sans répercussions sur le métabolisme primaire de la plante stressée (Schwachtje and Baldwin 2008). Celle-ci doit les déployer judicieusement, de manière à maximiser ses chances de survie sans trop compromettre sa croissance, sa reproduction et ses réserves énergétiques. Dans le but de maximiser leur valeur adaptative, les végétaux ont développé avec le temps des mécanismes de reconnaissance des stress, qui permettent d‟exprimer les moyens de défense nécessaires aux moments opportuns.
Ces mécanismes d‟induction sont de complexité variable. Parfois, la reconnaissance n‟implique qu‟un seul gène, comme dans le cas des réactions d‟incompatibilité avec certaines bactéries ou certains champignons phytopathogènes (Cui et al. 2005). Dans d‟autres cas, comme pour les insectes herbivores défoliateurs, la reconnaissance résulte de « l‟interprétation » d‟un grand nombre de stimuli. Une compréhension approfondie de ces mécanismes de reconnaissance facilitera certainement, avec le temps, le développement de moyens de lutte efficaces contre les ravageurs des cultures, basés sur une « optimisation raisonnée » des défenses intrinsèques de la plante.
Les paragraphes qui suivent décrivent plus en détail les interactions plantes-insectes à l‟échelle biochimique, en portant une attention particulière sur la réponse de la plante aux insectes défoliateurs comme le doryphore de la pomme de terre, un insecte d‟importance agronomique majeure. Le portrait type d‟un insecte défoliateur sera aussi tracé, en décrivant de quelle manière il est perçu comme un stress par sa plante hôte.
1.1 La réponse biochimique des plantes aux insectes défoliateurs
L‟interaction entre un insecte phytophage et sa plante hôte est un processus très complexe. En présence d‟un insecte spécialiste, l‟interaction est d‟autant plus complexe qu‟elle résulte de milliers, voire de millions d‟années de coévolution. Cette grande complexité rend évidemment impossible la prise en compte de tous les éléments impliqués dans l‟interaction et il devient alors important de développer des modèles d‟étude. Parfois très simples, parfois plus étoffés, ces modèles permettent de décomposer un système complexe en sous-systèmes de complexité moindre. Tout compte fait, l‟élucidation des sous-systèmes nous aide à mieux comprendre le processus global, un peu comme le fait de placer les morceaux d‟un casse-tête nous permet de visualiser l‟image finale.
1.1.1 Une description du stress « insecte défoliateur »
Lorsqu‟un insecte herbivore comme le doryphore de la pomme de terre s‟alimente, d‟importantes blessures sont infligées à sa plante hôte. Durant la prise alimentaire, de petits morceaux de tissus végétaux sont prélevés à répétition par les mandibules de l‟insecte, qui agissent ni plus ni moins comme des couteaux. La chair de la plante est alors déchirée et broyée pour être ensuite ingurgitée par l‟insecte, causant ainsi une réduction des surfaces photosynthétiques. Les tissus à proximité du site d‟alimentation sont également recouverts de régurgitant, un mélange de salive et de reflux intestinaux de l‟insecte, qui contient du matériel végétal partiellement digéré et des microorganismes. Les fèces de l‟insecte peuvent aussi entrer en contact avec les cellules végétales fraîchement endommagées. Enfin, certains insectes de plus grande taille peuvent endommager l‟épiderme des feuilles durant leur déplacement, dû à leur poids parfois important qui doit être supporté par des pattes menues, créant ainsi de multiples perforations (Hall et al. 2004).
Ces éléments qui caractérisent le stress « insecte défoliateur » peuvent être regroupés en deux catégories principales : 1- les éléments de nature mécanique et 2- les éléments de nature biochimique. Ils représentent les diverses composantes de l‟interaction qui permettront à la plante de décoder le stress « insecte défoliateur » et d‟ajuster le déploiement de ses défenses en fonction de son agresseur. Théoriquement, plus la reconnaissance sera efficace et rapide, plus la plante augmentera son aptitude à survivre et à se reproduire.
La Figure 1.1 décortique en sous-systèmes l‟impact d‟un stress causé par un insecte phytophage défoliateur « modèle ». Les composantes principales, mécanique et biochimique, intègrent plusieurs sous-composantes qui jouent un rôle potentiellement important dans l‟activation des mécanismes de défense de la plante hôte.
1.1.1.1 La composante mécanique
Des insectes phytophages défoliateurs comme le doryphore de la pomme de terre causent des bris mécaniques majeurs aux tissus végétaux lorsqu‟ils s‟alimentent. La plante hôte, de son côté, est capable de « reconnaître » ce type de stress.
Les insectes herbivores représentent un stress pour les plantes parce qu‟ils perturbent le fonctionnement normal de leur métabolisme. Les insectes défoliateurs, en particulier, affectent
directement leur plante hôte par le retrait de surfaces photosynthétiques. En tant qu‟organismes
phototautotrophes, les plantes sont entièrement dépendantes de l‟énergie lumineuse pour compléter leur cycle vital. Conséquemment, le retrait, le bris ou l‟isolation de tout tissu impliqué dans la production, le transport ou l‟entreposage des réserves énergétiques se traduit par une baisse de la valeur adaptative de la plante attaquée. L‟introduction de composés phytotoxiques à l‟intérieur des cellules lors de l‟alimentation de l‟insecte, comme des oxydases durant l‟alimentation des pucerons (Madhusudhan and Miles 1998), constitue une autre forme d‟effet direct des insectes herbivores sur la plante hôte.
De surcroît, les cellules endommagées des tissus touchés représentent une porte d‟entrée pour les bactéries et les champignons phytopathogènes. Elles constituent aussi une porte de sortie pour l‟eau. Par exemple, lorsque les tissus intervasculaires des feuilles de soya (Glycine max L.) sont blessés mécaniquement, leur taux de transpiration augmente jusqu‟à 150 % pendant six jours suivant la blessure (Aldea et al. 2005).
La plante blessée, dans ces conditions, a tout avantage à corriger la situation le plus rapidement possible. Pour y parvenir, elle doit détourner une partie plus ou moins importante de son métabolisme primaire (croissance, reproduction et mises en réserve) vers son métabolisme de défense, dit secondaire. Les blessures mécaniques occasionnées par les insectes herbivores ont
donc aussi un effet indirect sur la plante hôte, en provoquant un déséquilibre métabolique dans les cellules non blessées.
1.1.1.1.1 Les signaux chimiques
Les nombreux travaux de recherche effectués sur le sujet au cours des deux dernières décennies ont permis d‟identifier plusieurs signaux impliqués dans la réponse des plantes aux blessures et aux insectes herbivores. Bien que le « signal primaire » à la base de l‟ensemble des réponses n‟ait pas encore été identifié, des hypothèses ont été avancées suggérant que la propagation du signal de stress après blessure puisse être de nature hydraulique (Stratmann and Ryan 1997), électrique (Rhoades et al. 1996) ou chimique (Ryan 2000).
Plusieurs molécules bioactives (ou signaux chimiques) ont été identifiées dans les tissus avoisinant les sites de blessures (réponse locale) et dans les tissus distaux (réponse systémique). Par exemple, des blessures mécaniques induisent une accumulation transitoire locale et systémique d‟espèces réactives d‟oxygène (ROS, reactive oxygen species) dans les feuilles (van Breusegem et al. 2001), de même que la production d‟oligogalacturonides (Bergey et al. 1999) et d‟acide jasmonique (Farmer and Ryan 1990).
La systémine, première hormone peptidique identifiée chez les plantes (Pearce et al. 1991), est également retrouvée dans tous les tissus des Solanacées en réponse à des blessures et aux insectes herbivores (Ryan 2000). Ce polypeptide de 18 acides aminés possèderait les caractéristiques nécessaires pour agir comme médiateur du signal de stress des régions atteintes aux régions distales, et serait impliqué dans la réponse systémique. D‟autres phytohormones agissant à titre de médiateur chimique, comme l‟éthylène (O'Donnell et al. 1996) et l‟acide abscissique (Pena-Cortes et al. 1989), seraient aussi impliquées dans l‟induction de la réponse aux blessures mécaniques.
La majorité des réponses induites par les blessures surviennent rapidement. À l‟intérieur de quelques minutes ou quelques heures après le stress, il y a génération/relâchement, perception, translocation et « interprétation » de signaux spécifiques qui activeront les gènes de défense reliés aux blessures mécaniques (Leon et al. 2001). Par exemple, chez Arabidopsis, certains gènes de la famille TOUCH voient leur expression centupler en moins de dix minutes en réponse au toucher (Braam and Davis 1990). Les gènes TOUCH, ainsi que certains gènes encodant des protéines reliées à la pathogenèse (protéines P-R) et des kinases, sont aussi surexprimés dans les feuilles d‟Arabidopsis 15 minutes après une blessure physique (Reymond et al. 2000). Les voies de signalisation cellulaire impliquant la phosphorylation des protéines et le complexe calcium-calmoduline sont également sollicitées en réponse aux blessures mécaniques.
1.1.1.1.2 L’effet spatio-temporel
L‟effet spatio-temporel (ou effet espace x temps) détermine par ailleurs l‟intensité (ou la sévérité) des blessures mécaniques. Cette composante est importante dans la reconnaissance et l‟induction des mécanismes de défense chez les plantes. Il a été démontré à maintes reprises que la réponse des plantes aux blessures et aux insectes herbivores diffère tant sur le plan qualitatif que sur le plan quantitatif. L‟intensité des blessures serait d‟ailleurs un élément important dans la reconnaissance de l‟agresseur.
Les plantes attaquées par des insectes herbivores produisent des composés volatils qui attirent leurs ennemis naturels, une relation tritrophique qui témoigne de la coévolution des espèces dans l‟environnement (Turlings et al. 1990). Or, lorsque des plantes sont sujettes à des blessures simples, elles produisent une gamme de composés volatils qui diffèrent sur le plan qualitatif ou quantitatif, en comparaison à des plantes attaquées par des insectes herbivores. En revanche, des travaux réalisés par Mithofer et collaborateurs (2005) ont montré qu‟un robot permettant d‟infliger des blessures mécaniques complexes reproduisant le motif alimentaire des herbivores
(dans l‟espace et dans le temps) induisait une signature qualitative de composés volatils similaire à celle induite par les herbivores.
Pour des raisons pratiques, l‟effet spatio-temporel n‟est souvent pas complètement pris en compte dans les dispositifs expérimentaux où l‟on utilise un traitement « blessures » à titre de référence pour « mimer » l‟impact d‟un insecte défoliateur. Il est d‟ailleurs en général impossible de recréer un traitement de référence parfait. Afin d‟illustrer ces propos, prenons comme exemple une expérience où nous voudrions vérifier l‟effet des sécrétions orales d‟un insecte phytophage sur les réponses de sa plante hôte. Pour réaliser cette expérience, nous aurions besoin d‟un minimum de trois traitements : un traitement témoin sans stress, un traitement avec l‟insecte phytophage étudié et, finalement, un traitement « stress » de référence. Ce dernier traitement devrait idéalement engendrer le même type de blessures dans le temps et dans l‟espace que l‟insecte phytophage, mais sans qu‟il y ait déposition de sécrétions orales. La meilleure façon de réaliser ce traitement serait d‟utiliser des insectes dont on aurait éliminé l‟aptitude à excréter des fluides. Dans les faits, cette procédure est très difficile à réaliser et est aussi tributaire de l‟anatomie et de la morphologie de l‟insecte à l‟étude.
Un groupe de recherche de l‟Université de la Pennsylvanie a toutefois réalisé cette tâche avec succès. Musser et collaborateurs (2002) ont réussi à bloquer, par cautérisation, la sécrétion de salive labiale chez la larve du lépidoptère Manduca sexta. Les glandes labiales, qui sont les principales glandes salivaires chez M. sexta (Peiffer and Felton 2005), contiennent une grande quantité de glucose oxydase (GOX), une enzyme qui joue un rôle majeur dans la modulation de la réponse de défense du tabac (Nicotiana tabacum). Lorsque des plants de tabac sont blessés mécaniquement, leur taux de nicotine augmente en réponse au stress, par rapport à des plants non blessés (témoin). À l‟inverse, lorsque des plants de tabac sont attaqués par des larves de M. sexta, le taux de nicotine reste similaire à celui des plantes témoins. Ces résultats à eux seuls indiquent que les sécrétions orales de l‟insecte sont probablement impliquées dans la réponse différentielle observée entre les deux traitements de stress. Par contre, nous ne pouvons conclure sur l‟influence de l‟effet spatio-temporel dans une telle expérience. Rien n‟indique qu‟à lui seul, le
motif d‟application différentiel des blessures mécaniques occasionnées par les deux traitements n‟ait pu expliquer la différence d‟accumulation de nicotine observée.
En complément à ces observations, les auteurs ont montré que des plants de tabac attaqués par des larves incapables de sécréter de la salive labiale (conséquence de la cautérisation), mais dont le comportement alimentaire est inchangé, accumulent des taux de nicotine similaires aux plants blessés mécaniquement. En ajoutant ce dernier traitement à leur expérience, Musser et son équipe ont montré formellement que la salive labiale de l‟insecte prévenait l‟augmentation du taux de nicotine dans la plante, favorisant ainsi les taux de croissance et de survie de l‟insecte. Le tabac répondait de manière similaire à l‟attaque par d‟autres larves de lépidoptères, comme
Heliothis virescens (Delphia et al. 2006). Sur le plan biologique, il apparaissait clair, par ces
travaux, que la composante mécanique ne pouvait à elle seule expliquer la totalité des réponses observées chez une plante soumise à la défoliation par un insecte herbivore.
1.1.1.2 La composante biochimique
Au cours d‟une interaction plante-insecte, une multitude de signaux moléculaires sont échangés entre les deux partis. Ces molécules servent à communiquer des informations essentielles sur le partenaire d‟interaction. Par exemple, pour l‟insecte, ces signaux lui permettent de juger si les tissus de la plante représentent une source alimentaire adéquate ou non. Pour la plante, ils lui permettent d‟identifier son agresseur et de déployer des défenses adaptées afin d‟augmenter ses chances de survie sans trop affecter sa croissance et son développement (Kessler and Baldwin 2002).
Les molécules qui induisent l‟activation des mécanismes de défense chez les plantes sont appelées éliciteurs. Dans certains cas, les éliciteurs produits lors de l‟interaction induisent des mécanismes de défense chez la plante qui favorisent plutôt la survie de l‟insecte que celle de la
plante – on parle alors d‟effecteurs (Felton and Tumlinson 2008). Ces médiateurs chimiques sont transmis à la plante par la salive, le régurgitant ou les fèces de l‟insecte. À titre de référence, la salive est produite par les glandes salivaires alors que le régurgitant provient du tube digestif et est libéré par la cavité buccale. Le terme « sécrétions orales » est souvent utilisé, aussi, pour décrire le régurgitant. D‟autres glandes pourraient aussi être à l‟origine de signaux biochimiques reconnus par la plante hôte (Felton 2008).
Les éliciteurs « transmis » à la plante durant l‟alimentation de l‟insecte herbivore peuvent être de nature animale; c‟est le cas notamment de la β-glucosidase, retrouvée dans les sécrétions orales des larves du lépidoptère Pieris brassicae (Mattiacci et al. 1995). D‟autres éliciteurs sont d‟origine végétale, comme l‟inceptine (Schmelz et al. 2006), un peptide qui provient de la dégradation de l‟ATPase chloroplastique de la plante hôte. Cette protéine est partiellement hydrolysée dans le tube digestif de l‟insecte et un fragment stable, l‟inceptine, est reconnu par la plante pendant la régurgitation. Le mécanisme d‟action de l‟inceptine serait basé sur la reconnaissance d‟un fragment peptidique qui n‟est normalement pas produit dans les cellules végétales. Il est probable aussi que des molécules, qui sont normalement séquestrées à l'intérieur d'un compartiment cellulaire, soient libérées lorsque les feuilles sont broyées. Maintenant libres dans le régurgitant, elles pourraient être reconnues par la plante comme un indice d'herbivorie (Heil 2009). Il existe aussi des éliciteurs hybrides, qui résultent de la fusion de molécules en provenance de la plante et de l‟insecte. La volicitine fut le premier éliciteur hybride identifié, chez les larves de Spodoptera exigua (Alborn et al. 1997). Cette molécule est produite dans le régurgitant de l‟insecte, où un groupement hydroxyle et une glutamine sont ajoutés à l‟acide linolénique dérivé des tissus de la plante hôte (Paré and Tumlinson 1998).
Sur le plan expérimental, le rôle « éliciteur » des sécrétions orales est démontré lorsque l‟application du régurgitant d‟un insecte sur des blessures mécaniques artificielles restitue, en totalité ou en partie, la réponse de la plante hôte répondant à l‟herbivore. De façon similaire, un éliciteur spécifique est identifié lorsqu‟un composé du régurgitant reproduit, en totalité ou en partie, la réponse observée suite à l‟application de régurgitant. La réponse de la plante est
caractérisée en mesurant, par exemple, la production de composés volatils (quantité et qualité). L‟activation des sentiers métaboliques de défense comme ceux de l‟acide jasmonique, de l‟acide salicylique et de l‟éthylène; la production et l‟accumulation de molécules associées aux stress comme le peroxyde d‟hydrogène et les inhibiteurs de protéases; ou l‟activité d‟enzymes comme les peroxydases, sont souvent utilisées, aussi, comme indicateurs pour évaluer l‟effet inducteur des éliciteurs.
1.1.1.2.1 La salive
La salive des insectes herbivores est habituellement difficile à isoler et à récolter en quantité suffisante pour être étudiée avec les techniques analytiques conventionnelles. Pour cette raison, très peu d‟informations sont disponibles sur la composition chimique de la salive des insectes herbivores et sur son rôle dans l‟activation des réponses défensives des plantes.
Les lépidoptères possèdent deux paires de glandes salivaires : les glandes labiales (ou glandes à soie) et les glandes mandibulaires (Felton 2008). Les glandes labiales, généralement longues et tubulaires, convergent pour former un canal commun. Le contenu de ces glandes est sécrété par un organe spécialisé qui forme une petite protubérance, la filière (Felton and Eichenseer 1999). Les glandes labiales représentent les principales glandes salivaires de l‟insecte durant les stades larvaires. En stimulant la salivation à l‟aide d‟une solution de saccharose, on peut récolter la salive en quantité suffisante pour en déterminer la composition (Peiffer and Felton 2005). Jusqu‟à présent, seules des protéines ont été identifiées dans la salive labiale (Liu et al. 2004). Par contre, tout porte à croire que plusieurs autres groupes de composés soient aussi présents.
Par ailleurs, les glandes mandibulaires des lépidoptères sont généralement plus discrètes que les glandes labiales (Felton and Eichenseer 1999), quoique leur taille demeure variable entre les espèces (Parthasarathy and Gopinathan 2005). Ces glandes sont reconnues pour être riches en
lipides (Wroniszewska 1966; Eichenseer et al. 2002; Howard and Baker 2004) et leur contenu est déchargé par l‟entremise de pores situés sur les mandibules (Felton and Eichenseer 1999). Les coléoptères de la famille des Chrysomelidae, contrairement aux lépidoptères, ne possèdent pas de glandes labiales, mais plutôt une paire de glandes maxillaires (Nault 1997). Ces glandes, généralement longues et tubulaires, seraient déchargées à l‟extérieur par une ouverture dans l‟épiderme de la région cardo-stripitale des maxilla (Srivastava 1959). À notre connaissance, aucune étude n‟a été menée pour déterminer la composition spécifique du contenu des glandes maxillaires chez les Chrysomelidae.
La seule molécule clairement identifiée dans la salive des insectes herbivores comme ayant la capacité d‟induire une réponse défensive chez la plante hôte est la GOX. Cette enzyme fut d‟abord isolée dans les glandes labiales de larves d‟Helicoverpa zea (Eichenseer et al. 1999) et, par la suite, chez plusieurs autres lépidoptères (Merkx-Jacques and Bede 2004). Comme discuté précédemment, la GOX présente dans la salive des larves d‟H. zea provoque une inhibition de la surproduction de nicotine induite par les blessures (Musser et al. 2002) et permet d‟augmenter leurs chances de survie lorsqu‟elles s‟alimentent sur des plants de tabac. Pour cette raison, la GOX est considérée comme étant un effecteur qui, produit par les glandes salivaires de l‟insecte, cause une suppression des mécanismes de défense de la plante normalement induits en réponse aux insectes herbivores. Cette notion demeure toutefois relative : cette même molécule, lorsqu‟appliquée sur des blessures mécaniques chez le soya, agit comme un éliciteur des défenses contre des agents pathogènes et réduit l‟incidence des symptômes pathologiques causés par Pseudomonas syringae pv. glycinea (Felton and Eichenseer 1999).
Sur le plan biochimique, la GOX catalyse la conversion du D-glucose en acide D-gluconique et
en peroxyde d‟hydrogène (H2O2). Cette production d‟H2O2 serait à l‟origine de la suppression
des défenses la plante contre les herbivores. Cette forme réactive de l‟oxygène induit effectivement chez les plantes le sentier métabolique de l‟acide salicylique (AS), associé à la défense contre les agents pathogènes, un antagoniste bien caractérisé du sentier métabolique de l‟acide jasmonique (AJ), généralement associé à la protection contre les insectes herbivores
(Fidantsef et al. 1999; Stout et al. 1999; Felton and Korth 2000). C‟est donc par un mécanisme d‟interférence métabolique que la salive d‟H. zea confèrerait un avantage à l‟insecte et, par conséquent, une résistance accrue de la plante aux agents pathogènes (Fidantsef et al. 1999; Musser et al. 2005).
Ce type de stratégie offensive semble répandu dans la nature. Par exemple, la bactérie phytopathogène Pseudomonas syringae produit une toxine, la coronatine, qui mime l‟action de l‟AJ (Zhao et al. 2003). En induisant l‟activation de sentiers métaboliques antagonistes chez son hôte, la bactérie peut par la suite compléter son cycle vital et proliférer. Fait à noter, des plants d‟Arabidopsis infectés par P. syringae montrent une résistance induite systémique contre
Trichoplusia ni, un insecte herbivore défoliateur (Cui et al. 2005).
1.1.1.2.2 Le régurgitant
Plusieurs études ont montré que les sécrétions orales des insectes herbivores induisent l‟activation des mécanismes de défense chez la plante hôte. Néanmoins, seules quelques unes ont ciblé le ou les éliciteurs impliqués.
Les premiers travaux ayant mis en évidence le rôle des sécrétions orales des insectes herbivores sur l‟activation des mécanismes de défense de la plante hôte remontent au début des années 1990. Turlings et collaborateurs (1990) ont montré, par exemple, que l‟application de régurgitant du lépidoptère Spodoptera exigua sur des feuilles de maïs blessées restaurait la réponse observée lorsque les plantes étaient soumises à l‟insecte. Les plants de maïs émettent de grandes quantités de composés volatils en réponse aux larves de S. exigua. En revanche, des blessures mécaniques n‟induisent pas une telle production de composés volatils, à moins d‟y ajouter du régurgitant de
Korth et Dixon (1997) ont montré en outre, de manière éloquente, l‟importance du régurgitant dans la réponse aux insectes herbivores défoliateurs. L‟accumulation des transcrits d‟ARN messager du gène pinII, codant pour un inhibiteur de protéases, est plus rapide chez des plants de pomme de terre attaqués par des larves de Manduca sexta que chez des plants blessés mécaniquement. En ajoutant du régurgitant de l‟insecte sur les blessures, la cinétique d‟accumulation des transcrits de pinII était à l‟inverse similaire à celle observée chez les feuilles attaquées par M. sexta. Ces deux exemples classiques montrent que les sécrétions orales des insectes herbivores de type broyeur sont impliquées dans l‟activation des défenses indirectes et directes chez les végétaux.
Les résultats de plusieurs autres études ont aussi suggéré que le régurgitant de l‟insecte a un effet sur la réponse de la plante hôte, mais aucune preuve directe n‟appuie en général ces conclusions. C‟est le cas, par exemple, de mes travaux de maîtrise (Duceppe 2004). Comme nous avions uniquement comparé les réponses de la pomme de terre au doryphore et aux blessures mécaniques, nous ne pouvions affirmer avec confiance que les différentes réponses observées entre ces deux traitements étaient liées à la présence de sécrétions orales chez l‟insecte. Comme le projet visait à étudier la réponse de la plante à des stress de différentes natures, plutôt que l‟effet des sécrétions orales, nous n‟avions alors pas inclus de traitement avec le régurgitant de l‟insecte.
Le régurgitant contient de la salive, des tissus végétaux plus ou moins digérés, des sucs gastriques, des microbes provenant de l‟insecte ou de la plante hôte, et toutes sortes de particules potentiellement ingérées par l‟insecte. Toutes les molécules qui composent ces différentes fractions sont donc sujettes à entrer en contact avec les cellules de la plante hôte et bon nombre de celles-ci représentent des éliciteurs potentiels. Plusieurs éliciteurs ont été identifiés dans le régurgitant des insectes herbivores. Ils peuvent être regroupés en cinq familles, selon leur nature chimique (Felton and Tumlinson 2008): 1) les protéines, 2) les conjugués d‟acides gras, 3) les acides gras, 4) les peptides et 5) les oxilipines (Figure 1.2). Tous les éliciteurs identifiés jusqu'à maintenant ont été identifiés à partir du régurgitant de larves de lépidoptères (chenilles),
exception faite des éliciteurs de type « acides gras » retrouvés chez certains orthoptères (sauterelles).
Figure 1.2 Classes d‟éliciteurs identifiés dans les sécrétions orales des insectes herbivores.
Adapté de Felton et Tumlinson (2008). La structure tridimensionnelle de la β-glucosidase est tirée de Pozzo et al. (2010).
1. Les protéines : Le premier éliciteur identifié dans les sécrétions orales d‟un insecte
(Mattiacci et al. 1995). Il fallut ainsi cinq ans pour identifier le premier éliciteur du régurgitant d‟un insecte herbivore après avoir montré sa capacité à reproduire, du moins en partie, la réponse de la plante aux insectes herbivores lorsqu‟appliqué sur des blessures mécaniques. Certains composés volatils impliqués dans l‟attraction des parasitoïdes d‟insectes herbivores (défense indirecte) sont entreposés sous forme de β-glucosides chez le chou (Brassica oleracea); la β-glucosidase permettrait la libération de ces composés volatils. Une application ectopique de l‟enzyme sur des feuilles non blessées ou des blessures mécaniques seules n‟engendrent pas la libération des composés volatils attirant les prédateurs de l‟herbivore (Boland et al. 1992), alors que l‟application de β-glucosidase sur des blessures le fait.
La glucose oxydase est la seule autre protéine identifiée jusqu'à présent qui pourrait aussi faire partie de cette catégorie, bien qu‟elle soit plutôt considérée comme un effecteur (Felton and Tumlinson 2008). Le régurgitant du doryphore de la pomme de terre (Leptinotarsa decemlineata) contiendrait aussi des protéines qui pourraient agir à titre d‟effecteurs, mais leur identité reste inconnue (Lawrence et al. 2007).
2. Les conjugués d’acides gras (fatty acid-amino acid conjugates, ou FAC) : La volicitine
fut le premier éliciteur de cette famille à être identifié chez les larves du lépidoptère
Spodoptera exigua (Alborn et al. 1997), mais les FAC semblent être répandus chez les
insectes de cet ordre (Pohnert et al. 1999; Mori et al. 2001; Alborn et al. 2003). Chez tous les lépidoptères étudiés jusqu‟à maintenant, les FAC sont produits dans le jabot œsophagien et la partie antérieure du système digestif par l‟action d‟enzymes membranaires (Tumlinson and Lait 2005). Étrangement, les FAC sont rapidement hydrolysés dans le tube digestif de l‟insecte après leur synthèse. Ils pourraient servir de signaux transmetteurs transitoires ou de surfactants pour l‟insecte, mais leur rôle exact reste toujours à déterminer.
Les FAC retrouvés dans le régurgitant varient en fonction des aliments consommés par les larves. C‟est d‟ailleurs une stratégie qu‟a adoptée Heliothis subflexa, un lépidoptère qui s‟alimente sur les fruits de Physalis angulata. Ces derniers étant dépourvus d‟acide
linolénique, l‟insecte augmente ses chances de survie en ne produisant pas de volicitine (De Moraes and Mescher 2004). Chez la plante hôte, les FAC induisent la production systémique de composés volatils qui attirent les prédateurs et les parasitoïdes de lépidoptères. Il est intéressant de constater que les groupes fonctionnels ajoutés par l‟insecte à cet éliciteur hybride (un groupement hydroxyle et une glutamine ou un glutamate) semblent très importants dans la reconnaissance des FAC par la plante hôte (Truitt et al. 2004).
3. Les acides gras : Ces éliciteurs de composés volatils ont d‟abord été identifiés dans les
sécrétions orales des sauterelles Schistocerca americana (Alborn et al. 2007). Fréquents dans le sous-ordre des Cælifera chez les Orthoptera, ils ont été nommés « cæliférines ». Les cæliférines sont composées d‟une chaîne d‟acides gras saturée ou mono-insaturée de 15 à 20 carbones, au sein de laquelle le carbone α contient un groupement hydroxyle sulfaté, et le carbone ω un second groupement hydroxyle sulfaté ou une fonction carboxyle conjuguée à la glutamine par un lien amide. Cette structure moléculaire rappelle celle des FAC, mais la structure de ces composés ne change pas en fonction de la diète de l‟insecte.
4. Les peptides : Une nouvelle classe d‟éliciteurs, les éliciteurs peptidiques, fut identifiée
dans les sécrétions orales des insectes herbivores par Schmelz et collaborateurs (2006), qui cherchaient à identifier les composantes du régurgitant des larves du lépidoptère
Spodoptora frugiperda responsables de l‟émission d‟éthylène chez le niébé (Vigna unguiculata). Bien que les sécrétions orales de S. frugiperda contiennent des FAC,
ceux-ci n‟avaient pas la capaceux-cité d‟induire le relâchement caractéristique d‟éthylène observé suite à l‟application du régurgitant de l‟insecte. Ces résultats étaient surprenants, surtout que les FAC furent d‟abord identifiés chez le genre Spodoptera (Alborn et al. 1997). En décomposant le régurgitant de l‟insecte en multiples fractions, les auteurs ont identifié l‟agent responsable de l‟induction, un peptide qu‟ils ont nommé « inceptine ». L‟inceptine provient d‟une protéolyse partielle de l‟ATPase γ chloroplastique, une protéine conservée essentielle aux plantes, dans le système digestif de l‟insecte (Schmelz
et al. 2007). Ce fragment protéolytique n‟est habituellement pas produit dans la plante hôte, ou du moins n‟est pas exposé à son récepteur potentiel. Chez le niébé, la détection de S. frugiperda impliquerait ainsi la reconnaissance de fragments protéolytiques hétérologues, par un processus de reconnaissance du « non-soi ».
Les sécrétions orales de plusieurs lépidoptères possèdent la capacité d‟induire un flux d‟ions transmembranaire en conditions artificielles (Maischak et al. 2007), une activité cellulaire associée à la formation de pores. Or, des peptides présents dans le régurgitant de S. littoralis induisent une telle activité membranaire (Luhring et al. 2007). De surcroît, ces peptides seraient impliqués dans la diffusion de la volicitine dans les cellules de la plante hôte. Cette mobilité serait toutefois restreinte à de très courtes distances, puisque la volicitine n‟est pas retrouvée dans les feuilles non attaquées par les insectes herbivores (Truitt and Pare 2004). Ces peptides ne semblent donc pas être impliqués dans le processus d‟élicitation; leur rôle physiologique, ainsi que leur origine, restent toujours à déterminer.
5. Les oxilipines : Cette famille d‟éliciteurs n‟a été découverte que très récemment.
Gaquerel et collaborateurs (2009) ont décrit brièvement le rôle de l‟acide 2-hydroxyoctadécatriénoique (2-HOT) dans la perception des larves de Manduca sexta chez le tabac sauvage (N. attenuata), mais les données qui supportent leurs propos n‟ont pas encore été publiées. Cette oxilipine serait produite dans le régurgitant de l‟insecte à partir de l‟acide linolénique par l‟action d‟une α-dioxygénase, tous deux en provenance de la plante. Cette enzyme d‟origine végétale particulièrement stable est résistante à la protéolyse et active dans les conditions retrouvées dans le tube digestif de M. sexta. Comme les FAC et l‟inceptine, l‟éliciteur 2-HOT représenterait un éliciteur produit dans le régurgitant des insectes herbivores, mais dérivé au départ de la plante.
En somme, les éliciteurs sont des molécules que la plante hôte a « appris » à reconnaître et à utiliser au cours de sa coévolution avec l‟agresseur. Les éliciteurs sont parfois produits par l‟insecte directement, tandis que d‟autres dérivent de la plante, qui reconnaît alors ses propres molécules, souvent modifiées par l‟insecte.
Le concept de coévolution implique que les deux parties impliquées dans l‟interaction soient en constante bataille pour contrecarrer ou surclasser les moyens offensifs et défensifs de l‟adversaire, une sorte de course à l‟armement (Ryan and Byrne 1988). Dans cette optique, les sécrétions orales des insectes herbivores ont acquis, au fil du temps, de nouvelles fonctions qui ne sont pas directement reliées à la digestion proprement dite. En ce qui concerne la plante hôte, elle a « appris » à reconnaître certaines molécules présentes dans les sécrétions orales de ses ravageurs afin de bien cerner la nature du stress exercé et d‟adapter ses mécanismes de défense en conséquence. De leur côté, certains insectes herbivores trompent leur hôte en leur présentant des éliciteurs, les « effecteurs », qui induisent les « mauvais » mécanismes de défense, avec comme résultat une augmentation de leur valeur adaptative au détriment de la plante hôte.
Même si de plus en plus de recherches démontrent l‟importance du rôle des sécrétions orales dans les interactions plante-insecte, l‟implication des différents composés potentiellement actifs reste à déterminer dans bien des cas (Halitschke et al. 2003). Aussi, des travaux récents suggèrent que le régurgitant et la salive peuvent contribuer conjointement à la réponse observée chez la plante en conditions naturelles (Delphia et al. 2006). Par exemple, des plants de tabac (Nicotiana tabacum) blessés mécaniquement et traités avec du régurgitant de larves d‟Heliothis
virescens, un lépidoptère, relâchent plus de nicotine volatile que des plants traités avec de la
salive de l‟insecte ou des plants attaqués par l‟insecte lui-même. De plus, un mélange de salive et de régurgitant induit une libération plus faible de nicotine volatile que le régurgitant seul. Cet exemple met en évidence la complexité des systèmes biologiques en milieux naturels et démontre du même coup l‟importance d‟évaluer le rôle de l‟ensemble des fluides sécrétés lors des interactions plantes-insectes (Felton and Tumlinson 2008).