Impact of physicochemical properties of filtration
membranes on peptide migration and selectivity during
electrodialysis with filtration membranes: Development
of predictive statistical models and understanding of
mechanisms involved
Thèse
Sabita Kadel
Doctorat en sciences des aliments
Philosophiæ doctor (Ph. D.)
Québec, Canada
Impact of physicochemical properties of filtration
membranes on peptide migration and selectivity during
electrodialysis with filtration membranes:
Development of predictive statistical models and understanding
of mechanisms involved
Thèse
Sabita Kadel
Sous la direction de :
Laurent Bazinet, directeur de recherche
Carole Lainé, codirectrice de recherche
ii
Résumé
Au cours du procédé d'électrodialyse avec membrane de filtration (EDMF), les peptides chargés migrent sélectivement à travers des membranes de filtration (MFs) dans les compartiments respectifs de récupération des peptides anioniques (A-RC) ou cationiques
(C+
RC). Par conséquent, le type d'interaction entre les peptides et l'interface de la MF, en
raison de ses propriétés physicochimiques, doit avoir un impact significatif sur la performance globale de l’EDMF (migration et sélectivité des peptides). Donc, l'objectif principal de cette thèse de doctorat était d'étudier les propriétés physicochimiques principales des MFs qui contribuent aux interactions interfaciale peptide-membrane facilitant ou entravant la migration globale et la séparation sélective des peptides pendant l’EDMF, et de comprendre les mécanismes impliqués dans ces interactions. Ainsi, dans cette étude, 16 MFs, caractérisées en termes de propriétés physicochimiques (potentiel zêta, conductivité, nature hydrophile/hydrophobe de la surface et des pores, épaisseur, rugosité, porosité et pourcentage de distribution des macropores dans la couche filtrante), ont été testées lors de l'EDMF pour séparer simultanément les peptides anioniques et cationiques d'un hydrolysat de protéines de lactosérum complexe et bien caractérisé.
Dans la première étude, 6 MFs, différentes en termes de matériau, ont été testées incluant une membrane d’ultrafiltration (polyéthersulfone (PES)) comme contrôle et cinq membranes de microfiltration (fluorure de polyvinylidène (PVDF) et chlorure de polyvinyle (PVC-silice, fonctionnalisée (sulfopropyle ou amine quaternaire) ou non)). Les analyses de redondance (RDA) et de régression multivariées ont démontré qu’au moins deux des quatre propriétés suivantes des MF avaient un impact significatif sur la migration de tout peptide chargé ; le potentiel zêta, l’hydrophilie de surface/des pores, la porosité et la rugosité. De plus, l'effet important de la taille des pores sur la sélectivité des peptides a également été rapportée dans cette étude. Enfin, des modèles statistiques prédictifs qui relient la migration des peptides avec les propriétés de MF significatives ont été proposés.
Dans la deuxième étude, réalisée sur des membranes de PES avec une large gamme de seuils de coupure (MWCO) de (5 à 300 kDa), une relation linéaire a été observée entre le MWCO et la migration globale des peptides (MGP) pour les deux compartiments de récupération.
iii
Cependant, la migration sélective des peptides vers A-RC ou C+RC s'est révélée être influencée
par le MWCO des MFs ainsi que par les propriétés physicochimiques (charge et poids moléculaire (PM)) des peptides ; la migration d'un peptide ayant un faible PM et une faible charge (positive ou négative) était favorisée lorsqu’une MF ayant un petit MWCO était utilisée, tandis que l’inverse se produisait pour un peptide ayant un PM élevé et une charge élevée.
Dans la troisième étude, l'effet de la combinaison du matériau de la membrane (polyacrylonitrile (PAN), PES et PVDF) /MWCO (30 et 50 kDa) sur la migration et la sélectivité des peptides, a tout d’abord été étudié. Les effets simples du matériau membranaire et du MWCO sur la MGP vers C+
RC, de même que l'effet combiné des
matériaux membranaires/MWCO sur la MGP vers A
-RC et la migration sélective des peptides
vers les deux compartiments de récupération ont été observés. Deuxièmement, une RDA réalisée sur l’ensemble des données obtenues pour les MFs sélectives testées dans cette recherche doctorale, a démontré l'impact significatif du potentiel zêta, de la conductivité, de la rugosité et du pourcentage de distribution des macropores dans la couche filtrante des MFs sur la MGP. Concernant la migration sélective des peptides, en plus des propriétés des MFs susmentionnées, l'impact significatif de l'angle de contact a été démontré pour au moins la migration d’un peptide anionique et/ou cationique vers leurs compartiments de récupération respectifs. Ces propriétés significatives ont favorisé différentes interactions telles qu’électrostatique, exclusion de taille et hydrophile/hydrophobe entre l’interface de la MF et le peptide, ce qui a eu pour effet de, soit faciliter, soit inhiber la migration de ce peptide. Enfin, des modèles statistiques prédictifs globaux ont été développés pour la MGP et pour la migration de chaque peptide individuel vers A-RC et/ou C+RC en fonction des propriétés
importantes de la MF utilisée. Ces modèles permettent ainsi l'estimation du comportement de migration de ces peptides lorsque les MFs, sur une large gamme de propriétés physicochimiques, sont utilisées en EDMF.
Les résultats obtenus dans cette thèse ont démontré, pour la première fois, la corrélation significative entre les propriétés physicochimiques des MFs, et la migration et la sélectivité des peptides pendant l'EDMF. Cependant, les modèles prédictifs développés dans cette étude
iv
peuvent être utilisés pour la gamme de peptides et les propriétés physicochimiques des MFs testées. Par contre, les mécanismes et explications proposés dans cette étude, concernant les interactions MF/peptide, peuvent être généralisés afin de comprendre tous les types d'interactions peptide/membrane. Comme perspectives à ce travail, l’étude de différentes sources d'hydrolysats, d’autres MFs et d’un hydrolysat produit par d’autres enzymes permettra la validation de ces modèles statistiques et leur généralisation.
v
Abstract
During electrodialysis with filtration membranes (EDFM), charged peptides selectively migrate through filtration membranes (FMs) to their respective anionic (A-RC) or cationic
(C+RC) peptide recovery compartments. Consequently, the type of interactions occurring
between FM and peptide at the interface, due to their physicochemical properties, must have significant impact on overall EDFM performances (peptide migration and selectivity). Therefore, the main objective of this doctoral thesis was to investigate the major FM properties that contribute to peptide-membrane interactions at the interface, which either facilitates or hinders global migration and selective separation of peptides during EDFM, and to understand the mechanisms involved behind those interactions. Thus, in this study, 16 FMs, characterized in terms of their physicochemical properties (zeta potential, conductivity, hydrophilic/hydrophobic nature of the surface and pores, thickness, roughness, porosity and percentage of macropores distribution in filtrating layer) were tested during EDFM to simultaneously separate anionic and cationic peptides from a well-characterized complex whey protein hydrolysate.
In the first study, 6 FMs were tested, differing in terms of membrane materials, including one ultrafiltration (polyethersulfone (PES)) as a control and 5 microfiltration ( one polyvinylidene fluoride (PVDF) and four polyvinyl chloride (PVC)-silica: two functionalized (sulfonyl or amino) or two non-functionalized). Redundancy analysis (RDA) and multivariate regression analysis demonstrated that at least two FM properties among zeta potential, pore/surface hydrophilicity, porosity and roughness significantly impacted the migration of any charged peptide. In addition, the important effect of pore size on peptide selectivity was also reported. Finally, predictive statistical models that link each peptide migration with significant FM properties were proposed.
In the second study, which was carried out on PES membranes with a wide range of molecular weight cut-offs (MWCOs) (5 kDa to 300 kDa), a linear relation was noticed between MWCO and global peptide migration (GPM) to both recovery compartments. However, the selective peptide migration to A-RC or C+RC was found to be influenced by the
vi
MWCO of FMs as well as physicochemical properties (charge and molecular weight (MW)) of peptides. For instance, the migration of a peptide having low MW and low charge (positive or negative) was favored when a FM with small MWCO was used, while the opposite was observed for a peptide having high MW and high charge.
In the third study, the effect of combination of membrane material (PAN, PES and PVDF)/MWCO (30 and 50 kDa) on peptide migration and selectivity was first studied. The simple effect of membrane material and MWCO on GPM to C+RC was observed, while the
combined effect of membrane materials/MWCO on GPM to A-RC and selective peptide
migration to both recovery compartments was observed. Secondly, a RDA was performed on the data obtained for all the selective FMs tested in this doctoral research, which demonstrated the significant impact of zeta potential, conductivity, roughness and percentage of macropores distribution in the filtrating layer of FMs on GPM. Concerning selective peptide migration, in addition to the aforementioned FM properties, the significant impact of contact angle was noticed for at least one anionic and/or cationic peptide migration to their respective recovery compartments. These significant FM properties were found to trigger different interactions such as electrostatic, size exclusion and hydrophilic/hydrophobic between FM and peptide at the interface resulting in either facilitation or inhibition of peptide migration. Finally, global predictive statistical models were developed for GPM and each individual peptide migration to A-RC and/or C+RC based on these significant FM properties,
which allow the estimation of their migration behavior when FMs having a wide range of physicochemical properties are used during EDFM.
The results obtained in this Ph.D. thesis demonstrated, for the first time, the significant correlation between physicochemical properties of FMs, and peptide migration and selectivity during EDFM. The predictive models developed in this study can be used for the range of peptides and FMs tested. Moreover, the types of interactions occurring between FMs and peptide at the interface, and mechanisms and explanations proposed in this study can be applied to understand all types of peptide/membrane interactions. Validation of such models
vii
by using different sources of hydrolysates or different FMs or a hydrolysate produced by other enzymes will be the main perspectives of this research work.
viii
Table of contents
Résumé ... ii
Abstract ... v
Table of contents ... viii
List of tables ... xii
List of figures ... xv
List of abbreviations ... xviii
Acknowledgements ... xxiii
Forewords ... xxv
Introduction ... 1
1. Chapter 1: Literature review ... 5
1.1 Bioactive peptides ... 6
1.1.1 Generalities ... 6
1.1.2 Bioactive peptides from whey proteins ... 7
1.2 Bioactive peptides separation and purification methods ... 16
1.2.1 Chromatographic techniques ... 17
1.2.2 Isoelectric focusing ... 20
1.2.3 Membrane-based methods ... 21
1.3 Parameters affecting EDFM process ... 38
1.3.1 Physicochemical characteristics of feed solution ... 39
1.3.2 Operating parameters of EDFM ... 41
1.3.3 Filtration membranes and their physicochemical properties ... 41
1.4 Redundancy analysis and multivariate regression models... 59
2. Chapter 2: Problematic, hypothesis and objectives ... 63
2.1 Problematic ... 64 2.2 Research questions ... 64 2.3 Hypothesis ... 65 2.4 Objectives ... 65 2.4.1 Main objective ... 65 2.4.2 Specific objectives ... 65
3. Chapter 3: Use of redundancy analysis and multivariate regression models to select the significant membrane properties affecting peptide migration during electrodialysis with filtration membranes ... 67
ix
3.1 Contextual transition ... 68
3.2 Résumé ... 69
3.3 Abstract ... 70
3.4 Introduction ... 71
3.5 Materials and methods ... 72
3.5.1 Chemicals ... 72
3.5.2 Membrane materials ... 72
3.5.3 Raw materials ... 74
3.5.4 Electrodialysis cell and EDFM configuration ... 75
3.5.5 Protocol ... 76
3.5.6 Analyses ... 77
3.6 Results ... 79
3.6.1 Total cationic and anionic peptides migration ... 79
3.6.2 Peptide identification in recovery compartments ... 81
3.6.3 Multivariate analyses: RDA and regression models ... 86
3.7 Discussion ... 92
3.7.1 Conductivity, surface charge and electrostatic interactions ... 92
3.7.2 Porosity and size exclusion ... 94
3.7.3 Roughness (Ra and Rz) ... 95
3.7.4 Pore hydrophilicity and surface contact angle ... 96
3.7.5 Thickness ... 97
3.8 Conclusion ... 97
3.9 Acknowledgements ... 98
3.10 Achievement of objectives and advancement of knowledge ... 99
4. Chapter 4: How Molecular Weight Cut-Offs and Physicochemical Properties of Polyether Sulfone Membranes Affect Peptide Migration and Selectivity during Electrodialysis with Filtration Membranes ... 105
4.1 Contextual transition ... 106
4.2 Résumé ... 107
4.3 Abstract ... 108
4.4 Introduction ... 109
4.5 Materials and methods ... 111
4.5.1 Chemicals ... 111
x
4.5.3 Whey protein hydrolysate ... 112
4.5.4 Electrodialysis cell and EDFM configuration ... 113
4.5.5 Protocol ... 114
4.5.6 Analyses ... 114
4.6 Results and discussion ... 119
4.6.1 Physicochemical properties of FM ... 119
4.6.2 Global rate of peptide migration to recovery compartments ... 121
4.6.3 Individual peptide migration to recovery compartments: Peptides selectivity ………...125
4.7 Conclusion ... 131
4.8 Acknowledgements ... 132
4.9 Achievement of objectives and advancement of knowledge ... 133
5. Chapter 5: How physicochemical properties of filtration membranes impact peptide migration and selectivity during electrodialysis with filtration membranes: Development of predictive statistical models and understanding of mechanisms involved ... 134
5.1 Contextual transition ... 135
5.2 Résumé ... 136
5.4 Abstract ... 138
5.5 Introduction ... 140
5.6 Materials and methods ... 142
5.6.1 Membranes ... 142
5.6.2 Whey protein hydrolysate ... 142
5.6.3 EDFM configuration ... 143
5.6.4 Protocols ... 144
5.6.5 Analyses ... 145
5.7 Results ... 148
5.7.1 Impact of MWCO/membrane material combination on peptide migration and selectivity during EDFM ... 148
5.7.2 Redundancy analysis and predictive statistical models ... 157
5.8 Discussion ... 167
5.8.1 Impact of physicochemical properties of FMs on peptides migration ... 168
5.8.2 Interactions between FMs and peptides due to their respective physicochemical properties ... 176
xi
5.10 Acknowledgement ... 186
5.11 Achievement of objectives and advancement of knowledge ... 187
General discussion, conclusion and perspectives ... 194
General discussion ... 195
Back to the hypothesis ... 198
Conclusion and perspectives ... 199
xii
List of tables
• Chapter 1Table 1.1: Peptides derived from the tryptic hydrolysis of β-lg (Adapted and modified from [45,46]) ... 11 Table 1.2 :Bioactive peptides obtained by the enzymatic hydrolysis of bovine β-lg (Adapted and updated from [47,48]) ... 12 Table 1.3: Peptides derived from the enzymatic hydrolysis of α-la (Adapted and updated from [46]) ... 15 Table 1.4: Bioactive peptides obtained by enzymatic hydrolysis of bovine α-la (Adapted and updated from [47,48]) ... 16
• Chapter 3
Table 3.1: Physicochemical properties of filtration membranes measured prior EDFM ... 73 Table 3.2: Characterization of peptides obtained after tryptic hydrolysis of Prolacta ... 75 Table 3.3: Identification of peptides recovered from cationic recovery compartment after 120 min of EDFM for PES, PVDF, CF55, S11, S11+ and S11- membranes ... 82 Table 3.4: Identification of peptides recovered from anionic recovery compartment after 120 min of EDFM for PES, PVDF, CF55, S11, S11+ and S11- membranes ... 84 Table 3.5: Significant estimated coefficient for individual peptide migration into C+RC ... 89
Table 3.6: Significant estimated coefficients for individual peptides migration into A
-RC .. 92
Table 3.7: Spearman coefficients of physicochemical characteristics of membranes for each peptide migration to cationic recovery compartment ... 103 Table 3.8: Spearman coefficients of physicochemical characteristics of membranes for each peptide migration to anionic recovery compartment ... 104
• Chapter 4
Table 4.1: Characteristics of major peptides obtained after tryptic hydrolysis of a whey protein isolate ... 112 Table 4.2: Physicochemical properties of PES membranes measured prior EDFM ... 120 Table 4.3: Relative abundance of identified peptides recovered from anionic recovery compartment after 120 min of EDFM for PES membranes with MWCO of 5, 10, 20, 50, 100 and 300 kDa. ... 128
xiii
Table 4.4: Relative abundance of identified peptides recovered from cationic recovery compartment after 120 min of EDFM for PES membranes with MWCO of 5, 10, 20, 50, 100 and 300 kDa. ... 131
• Chapter 5
Table 5.1. Characteristics of major peptides obtained after tryptic hydrolysis of a whey protein isolate [173]. ... 142 Table 5.2: Physicochemical properties of filtration membranes measured prior EDFM ... 150 Table 5.3: Relative abundance of identified/not identified peptides recovered from anionic recovery compartment after 120 min of EDFM using PAN, PES and PVDF membranes with MWCO of 30 and 50 kDa... 155 Table 5.4: Relative abundance of identified/not identified peptides recovered from cationic recovery compartment after 120 min of EDFM using PAN, PES and PVDF membranes with MWCO of 30 and 50 kDa... 156 Table 5.5: Significant estimated coefficients for individual peptide migration to A-RC ... 164
Table 5.6: Significant estimated coefficients for individual peptide migration to C+RC .... 167
Table 5.7: Best FM/FMs amongst the tested ones in this study to separate major bioactive peptides from whey protein hydrolysate obtained by tryptic hydrolysis (FM/FMs in the table is selected based on the highest RA of a specific peptide obtained when respective FM was used). ... 186 Table 5.8: Physicochemical properties of FMs that were taken from our previous studies (Chapter 3 and Chapter 4) as well as the new data on porosity and percentage of macropores distribution in filtrating layer for RDA and multivariate regression analysis. ... 188 Table 5.9: Global rate of peptide migration to recovery compartments through FMs that were taken from our previous studies (Chapter 3 and 4) for RDA and multivariate regression analysis. ... 188 Table 5.10: Relative abundance of peptides migrated to anionic recovery compartment through FMs that were taken from our previous studies (Chapter 3 and Chapter 4) for RDA and multivariate regression analysis. ... 189 Table 5.11: Relative abundance of peptides migrated to cationic recovery compartment through FMs that were taken from our previous studies (Chapter 3 and Chapter 4) for RDA and multivariate regression analysis. ... 189 Table 5.12: Two-way ANOVA results for global peptide migration to recovery compartments affected by MWCO and membrane material combination. ... 189 Table 5.13: Two-way ANOVA results for individual peptide migration to A-RC affected by
MWCO and membrane material combination. ... 190 Table 5.14: Two-way ANOVA results for individual peptide migration to C+RC affected by
xiv
Table 5.15: Significant FM properties that affect global peptide migration to recovery compartments, along with their associated t-value. The higher the t-value (in absolute value), the greater importance is the related variable. Negative t-value associated with a FM property would mean its negative correlation with global peptide migration. (Ra is excluded from the table since it does not represent the actual surface topography) ... 191 Table 5.16: Significant FM properties that affect individual peptide migration to anionic recovery compartment, along with their associated t-value. The higher the t-value (in absolute value), the greater importance is the related variable. Negative t-value associated with a FM property would mean its negative correlation with global peptide migration. (Ra is excluded from the table since it does not represent the actual surface topography) ... 191 Table 5.17: Significant FM properties that affect individual peptide migration to cationic recovery compartment, along with their associated t-value. The higher the t-value (in absolute value), the greater importance is the related variable. Negative t-value associated with a FM property would mean its negative correlation with global peptide migration. (Ra is excluded from the table since it does not represent the actual surface topography) ... 192
xv
List of figures
• Chapter 1Figure 1.1: Primary structure of β-lg A (Adapted and modified [43]) ... 9
Figure 1.2: Primary structure of α-la A (Adapted from [49]) ... 14
Figure 1.3: Pressure-driven membranes according to the size of the particles retained. (Adapted from [61,62] and modified) ... 22
Figure 1.4: Schematic representation of concentration polarization and fouling during pressure driven membrane separation processes (Adapted and modified from [47]). ... 25
Figure 1.5: Configuration and principle of EMF for selective transport of positively charged peptides (cationic peptides) when an electric field is applied across the flat-sheet membranes. AEM: Anion-exchange membrane, CEM: Cation-exchange membrane and FM: Filtration membrane (Adapted and modified from [72]) ... 27
Figure 1.6: Principle of electrophoretic membrane contractor. CEM: Cation-exchange membrane, AEM: Anion-exchange membrane, FM: Filtration membrane. (Adapted and modified from [14]) ... 29
Figure 1.7: Schematic diagram illustrating the principle of electrodialysis (CEM: Cation-exchange membrane, AEM: Anion-Cation-exchange membrane. Adapted and modified from [78]) ... 31
Figure 1.8: Formation of concentration gradients during electrodialysis. AEM: Anion-exchange membrane, CEM: Cation-Anion-exchange membrane, δ1: Diluate boundary layer, δ2: Concentrate boundary layer (Adapted and modified from [80]) ... 33
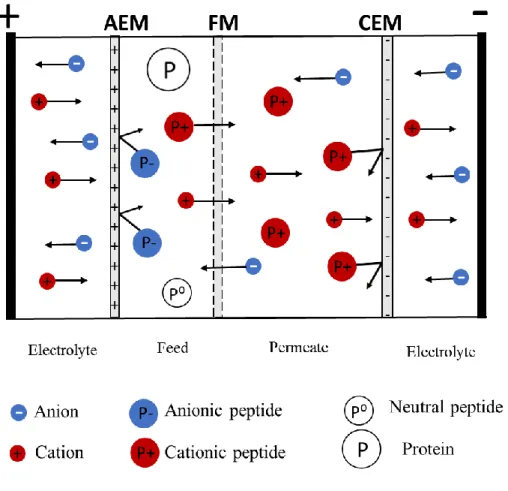
Figure 1.9: Electrodialysis with filtration membrane configuration for the separation of (a) anionic peptides and (b) cationic peptides. AEM: Anion-exchange membrane, CEM: Cation-exchange membrane, FM: Filtration membrane, P-: Anionic peptide, P+: Cationic peptide, P0: Neutral peptide (Adapted and modified from [86]) ... 35
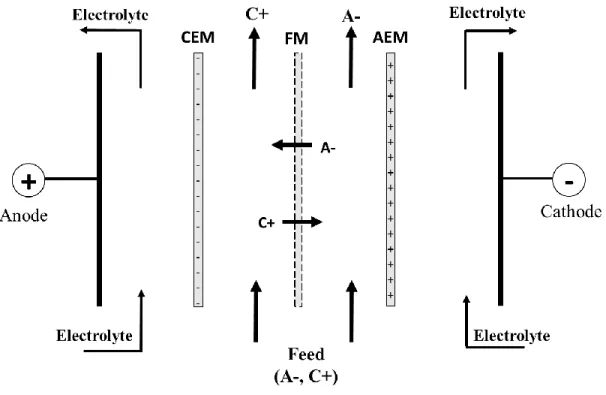
Figure 1.10: Configuration of the EDFM using two filtration membranes for the simultaneous separation of cationic and anionic peptides. FM: Filtration membrane. AEM: Anion-exchange membrane, and CEM: Cation-Anion-exchange membrane. (Adapted and modified from [88]) ... 36
Figure 1.11: Molecular structure of PES ... 44
Figure 1.12: Molecular structure of PVDF ... 45
Figure 1.13: Molecular structure of PAN ... 46
Figure 1.14: Schematic representation of zeta potential of filtration membranes (Adapted and modified from [116]) ... 48
Figure 1.15: Working principle of Stylus profilometer (Adapted and modified [125]) ... 51
xvi
Figure 1.17: An illustration of membrane tortuosity (adapted and modified from [59]) ... 56 Figure 1.18: Schematic representation of RDA correlation biplot ... 62
• Chapter 3
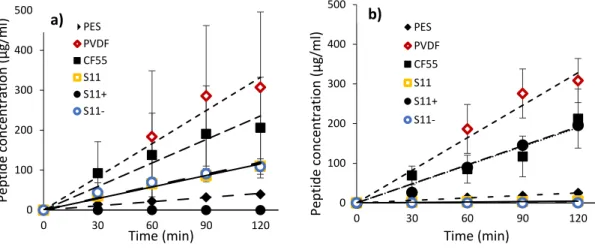
Figure 3.1: EDFM cell configuration for the simultaneous separation of anionic and cationic whey peptides. FM1 and FM2 are filtration membranes with same MWCO. P+: cationic peptides, P-: anionic peptides respectively. C+: cations, A-: anions, AEM: Anion-exchange membrane and CEM: Cation-exchange membrane ... 76 Figure 3.2: Evolution of peptides concentration in (a) cationic and (b) anionic compartment as a function of time during EDFM. ... 80 Figure 3.3: Biplot diagram of redundancy analysis of peptides recovered from C+RC according
to the membrane properties with (a) and without collinearity (b) ... 88 Figure 3.4: Biplot diagram of redundancy analysis of peptides recovered from A-RC according
to the membrane properties with (a) and without collinearity (b,c) ... 91 Figure 3.5: UV profile of Prolacta hydrolysates after digestion by trypsin ... 100 Figure 3.6: UV profiles of peptides recovered from cationic recovery compartment after 120 min of EDFM for PES, PVDF, CF55, S11, S11+ and S11- membranes ... 101 Figure 3.7: UV profiles of peptides recovered from anionic recovery compartment after 120 min of EDFM for PES, PVDF, CF55, S11, S11- and S11+ membranes ... 102
• Chapter 4
Figure 4.1: EDFM cell configuration for the simultaneous separation of anionic and cationic whey protein hydrolysate. FM1 and FM2 are filtration membranes with same MWCO, AEM: anion-exchange membrane and CEM: cation-exchange membrane, P+: cationic peptide, P-: anionic peptide, P0: neutral peptide (adapted from [171])... 113 Figure 4.2: Distribution of percentage macropores as a function of MWCO ... 121 Figure 4.3: Evolution of peptide concentrations in (a) anionic and (b) cationic peptide recovery compartments as a function of time during EDFM. ... 122 Figure 4.4: Rates of global peptide migration (g/m2.h) to a) anionic and b) cationic recovery
compartment as a function of MWCO. ... 125 • Chapter 5
Figure 5.1: EDFM cell configuration for the simultaneous separation of anionic and cationic whey protein hydrolysate. FM1 and FM2 are filtration membranes with the same MWCO, AEM: anion-exchange membrane and CEM: cation-exchange membrane, P+: cationic peptide, P−: anionic peptide, P0: neutral peptide ... 144
Figure 5.2: Evolution of peptides concentration in (a) anionic and (b) cationic compartments as a function of time during EDFM. ... 153
xvii
Figure 5.3: RDA biplot between global peptide migrations to recovery compartments and FM properties after collinearity diagnostic ... 159 Figure 5.4: RDA biplots between anionic peptide migration and FM properties after collinearity diagnostic a) RDA 1 Vs RDA 2 and b) RDA 1 Vs RDA 3. ... 163 Figure 5.5: RDA biplot between cationic peptide migration and FM properties after collinearity diagnostics a) RDA 1 Vs RDA 2 and b) RDA 1 Vs RDA 3. ... 166 Figure 5.6: Various surface profiles having same Ra value. Adapted and modified from [123] ... 172 Figure 5.7: Schematic representation of anionic peptides repulsed from, and cationic peptides adhered and trapped on the peak and valley regions of the negatively charged membrane surface. ... 174 Figure 5.8: Schematic representation of migration of major small anionic peptides through FM (regardless of their pore size) to anionic recovery compartment, A) Migration of linear IDALNENK when it arrives parallel or perpendicular at the interface, B) migration of linear VLVLDTDYK when it arrives parallel or perpendicular at the interface and C) migration of both peptides when they have distorted conformations. ... 179 Figure 5.9: Case I: Schematic representation of migration of major large anionic peptides through FM having small pores to anionic recovery compartment. A) and C) Migration of linear TPEVDDEALEK and VYVEELKPTPEGDLEILLQK, respectively, when they arrive parallel or perpendicular at the interface, B) and D) migration of these large peptides when they have distorted conformations. Case II: Migration of these large peptides when they have linear or distorted conformations ... 180 Figure 5.10: Schematic representation of migration of major cationic peptides through FM (regardless of their pore size) to cationic recovery compartment, A) Migration of linear IPAVFK when it arrives parallel or perpendicular at the interface, B) migration of linear ALPMHIR when it arrives parallel or perpendicular at the interface and C) migration of both peptides when they have distorted conformations ... 183 Figure 5.11: Schematic representation of hydrophobic interactions occurring between (A) neutral and anionic peptides, and (B) neutral and cationic peptides ... 184 Figure 5.12: Schematic representation of hydrophobic and electrostatic interactions occurring between anionic and cationic peptides to form a globally positively charged aggregate... 184
xviii
List of abbreviations
ACE: Angiotensin-converting enzyme AEM: Anion-exchange membrane α-la: Alpha lactalbuminAPM : Global peptide migration to anionic recovery compartment A-RC: Anionic peptide recovery compartment
β-lg: Beta lactoglobulin BP: Bioactive peptide
CEM: Cation-exchange membrane
CPM: Global peptide migration to cationic recovery compartment C+RC: Cationic peptide recovery compartment
Da: Dalton
ED: Electrodialysis
EDFM: Electrodialysis with filtration membranes EMF: Electromembrane filtration
EMC: Electrophoretic contactor FM: Filtration membrane GPM: Global peptide migration
GRAVY: Grand average of hydropathy index IEM: Ion-exchange membrane
kDa: Kilo Dalton MF: Microfiltration MW: Molecular weight
MWCO: Molecular weight cut-off
mS: Milli-Siemens (electrical conductivity unit) NF: Nanofiltration
PAN: Polyacrylonitrile PES: Polyether sulfone
xix
PSD: Pore size distribution PVC: Polyvinyl chloride PVDF: Polyvinylidene fluoride Ra: Average roughness
RA: Relative abundance RDA : Redundancy analysis RO : Reverse osmosis
Rz : Average peak-to-valley height of the profile UF: Ultrafiltration
UPLC-MS-QTOF: Ultra-high-performance liquid chromatography-mass spectrometry-quadrupole time-of-flight
VIF: Variation inflation factor WPI: Whey protein isolate ZP: Zeta potential
List of amino acids with their code.
Amino acids Three letter code One letter code
Alanine Ala A Glycine Gly G Valine Val V Leucine Leu L Isoleucine Ile I Proline Pro P Phenylalanine Phe F Tyrosine Tyr Y Tryptophan Trp W Serine Ser S Threonine Thr T Cysteine Cys C Methionine Met M Asparagine Asn N Glutamine Gln Q Lysine Lys K Arginine Arg R Histidine His H
xx
Aspartate Asp D
xxi
To my beloved family, specially my Father and Mother, who are my source of motivation.
xxii
“Failure will never overtake me if my determination to succeed is strong enough”
xxiii
Acknowledgements
I would like to express my sincere gratitude to everyone, who supported and motivated me throughout my incredible Ph.D. journey.
First of all, I am very grateful to my thesis supervisor, Professor Laurent Bazinet, for believing in me and providing me the opportunity to work on this project. Thank you very much for your aspiring guidance, availability, and invaluable suggestions throughout. The way you manage time for your students for regular discussion despite your very busy schedule is praiseworthy. Without your continuous help, support, encouragement and motivation, the goal of this project would not have been realized. Also, thank you very much for the barbeque, and Christmas parties you hosted at your house every year. You are the best professor/supervisor any students could ask for. I am very proud and feel blessed to be one of your students.
I am equally thankful to my thesis co-supervisor, Dr. Carole Lainé. Your valuable remarks and insightful suggestions during the project meetings and article writing were invaluable.
I am very grateful to Professor Jean-Christophe Vuillemard for accepting to pre-read my thesis. At the same time, I am sincerely thankful to Professor Yves Pouliot and Professor Michel Britten for accepting to be in the jury committee of this thesis.
Also, I would like to thank my research team. Thank you everyone for your moral support during my seminars. Your presence used to boost up my confidence during my presentations in French. I would like to extend my deepest appreciation to Jacinthe Thibodeau and Véronique Perreault for their continuous support during lab experiments, suggestions during presentations, help in writing abstracts and posters in French, and ordering lab materials. Jacinthe, thank you very much for being so kind to me since day 1. More than that, thank you for always being there as a support system during my difficult times, like a true friend. Thank you for everything.
xxiv
The technical help from the laboratory personnel is highly appreciated. Thank you very much Ms. Diane Gagnon, and Mr. Pascal Lavoie, from Department of Food Science, for your technical help in the lab and pilot lab. Also, the help of Dr. Pascale Chevallier from Laboratoire de Biomatériaux et de Bioingénierie is highly appreciated. Likewise, I am grateful to Gaétan Daigle, Statistical consultant from the Department of Mathematics and Statistics, Université Laval, for his generous help and guidance on advance statistical methods used in this study.
I am very thankful to my friends for making my stay in Québec full of memories. Particularly, my best friends, Dr. Sagar Bhatta, thank you very much for all your support, motivation and encouragement, and also for being that one friend with whom I can share my any secrets, and Mélanie Faucher, my Ph.D. journey wouldn’t have been the same without you, I will treasure our friendship for the whole life. I would like to pay my special regard to Dr. Shyam Suwal and his family. Shyam dai, you are like a brother to me. Thank you very much for your guidance since 2011. My deep appreciation goes to all the wonderful souls who have motivated me this or that way to reach my goal, Dr. Sergey Mikhaylin (you are such a motivator), Dr. Thibaud Geoffroy, Dr. Romari Meda, Léa, Carine and our small Nepali community.
At last, I am indebted to my family, my parents, sisters, and brothers, who, despite being far away by distance, have always been there with me through all my ups and downs. I would not have been here today without your love and support. I hope I have made you proud. Finally, my most favorite person, the love of my life, Rajib, thank you very much for your unconditional love and support. Thank you for believing in me, boosting up my confidence whenever I feel down and always making me realize that I can overcome any difficult situations. Life has been better and more beautiful with you.
xxv
Forewords
This PhD thesis was carried out within the framework of NSERC Industrial Research Chair on “Electromembrane processes, aiming the ecoefficiency improvement of biofood production lines”. Most of the research work of this thesis were carried out in the “Laboratoire de transformation alimentaire et procédés électromembranaires”, while some were performed at “Centre de recherche sur les matériaux avancés”.
This document consists of 5 chapters in total. The first chapter presents a literature review, which provides state of knowledge on existing research relevant to this study based on which research hypothesis was proposed. Furthermore, information on raw materials (such as whey protein hydrolysate), filtration membranes, and tools and techniques used within the framework of this study to achieve the research goal were also discussed in this section. The second chapter presents problematic, hypothesis and objectives of the present study. The third, fourth and fifth chapters correspond to the experimental works carried out throughout this project along with their results. These research works are either published or submitted in relevant international scientific journals.
The third chapter presents the article entitled “Use of redundancy analysis and multivariate regression models to select the significant membrane properties affecting peptide migration during electrodialysis with filtration membranes” published in “Separation and Purification Technology” 221 (2019), 114-125. The authors are Sabita Kadel (PhD Candidate: Conceptualization, experimental design and execution of experiments, result analysis and writing the manuscript), Mathieu Persico (Manuscript correction and revision), Jacinthe Thibodeau (Software handling), Carole Lainé (Thesis co-supervisor: Correction and revision of the manuscript) and Laurent Bazinet (Scientific supervision, conceptualization, results validation, correction and revision of manuscript). The results obtained in this chapter were presented in the form of “Oral presentation” at the 27th Annual meeting of North American Membrane Society (NAMS) which was held in Lexington, KY, USA in June 2018. Furthermore, this work presented in the form of “Poster presentation”
xxvi
won second prize in Green Food Tech conference, which was held in Quebec City, QC, Canada in October 2018.
The fourth chapter presents the article entitled “How Molecular Weight Cut-Offs and Physicochemical Properties of Polyether Sulfone Membranes Affect Peptide Migration and Selectivity during Electrodialysis with Filtration Membranes” published in “Membranes” 9 (2019). The authors are Sabita Kadel (PhD Candidate: Conceptualization, experimental design and execution of experiments, result analysis and writing the manuscript), Geneviève Pellerin (Execution of experiments), Jacinthe Thibodeau (Software handling), Véronique Perreault (Execution of experiment), Carole Lainé (Thesis co-supervisor: Correction and revision of the manuscript) and Laurent Bazinet (Scientific supervision, conceptualization, results validation, correction and revision of manuscript). The results obtained in this chapter were presented in the form of “Oral presentation” in 28th
Annual meeting of North American Membrane Society (NAMS) which was held in Pittsburgh, PA, USA in May 2019. Furthermore, this work presented in the form of “Poster presentation” won third prize in STELA Symposium, which was held in Quebec City, QC, Canada in May 2019.
The fifth chapter presents the article entitled “How physicochemical properties of filtration membranes impact peptide migration and selectivity during electrodialysis with filtration membranes: Development of predictive statistical models and understanding of mechanisms involved”, submitted in “Journal of Membrane Science” for publication. The authors are Sabita Kadel (PhD Candidate: Conceptualization, experimental design and execution of experiments, result analysis and writing the manuscript), Gaétan Daigle (Statistical software and analysis) Geneviève Pellerin (Execution of experiments), Jacinthe Thibodeau (Software handling), Véronique Perreault (Execution of experiment), Carole Lainé (Thesis co-supervisor: Correction and revision of the manuscript) and Laurent Bazinet (Scientific supervision, conceptualization, results validation, correction and revision of manuscript).
xxvii
Finally a general conclusion and key findings of the present research work and future perspectives of this study are presented.
1
Introduction
The importance of food-derived peptides has been increasingly acknowledged in recent decades because of their proven bioactivities such as antihypertensive, antithrombotic, antimicrobial, antioxidative, immunomodulatory, opioid, mineral binding, among others, beyond their normal nutritional value [1–4]. Such health-promoting effects of those bioactive peptides (BPs) make them potential ingredients of functional foods, nutraceuticals, or
pharmaceutical products. Consequently, there is a burgeoning number of
food/pharmaceutical companies that are inclined to incorporate BPs in the formulation of such products [1]. However, production, separation, and concentration of BPs at commercial scale remained delayed due to the lack of suitable large-scale technologies. Since, BPs are initially present in the native protein sequence in inactive form, proteolysis of protein must be carried out to liberate them [5]. This results in the production of a complex protein hydrolysate composed of a large number of active and inactive hydrolyzed protein fractions having various sizes, amino acid compositions, functional properties and biological functions [6]. Consequently, an appropriate and efficient fractionation technique is required to selectively separate these active peptides from the complex mixture and concentrate them in order to yield the enriched bioactive peptide fractions or a specific bioactive peptide, with improved biological activity and/or nutritional values.
Different methods exist for the fractionation and concentration of bioactive peptides such as chromatographic techniques [7–9], electrophoretic-based isoelectric focusing [10] and membranes-based processes [11–14]. Chromatographic techniques, in spite of effectively separating and purifying the peptides, are mainly limited to lab scale due to unacceptable economics. Furthermore, the use of chemical solvents, time-consuming, applicable for small volume only and complex operation process are other cons associated with these techniques [15]. Similarly, regardless of higher sensitivity, quality and purity of the final product, isoelectric focusing (IEF) is not applicable at industrial scale because of high processing cost, limited to small sample volume, and too long run time (24h) [10,16]. Among all, membrane-based processes are considered as the potential one for peptides separation at commercial
2
scale because of their acceptable economy (thanks to their low processing cost and relatively low use of energy), no use of solvents making it an environment friendly (green) technology, easy to scale up and use of mild working conditions such as ambient temperature (therefore, helps to maintain the biological activity and structural integrity of peptides) [11]. Membrane-based processes such as pressure-driven ones (mainly ultrafiltration (UF) and nanofiltration (NF)) have been regularly used for the separation and concentration of bioactive peptides from various sources of protein hydrolysate such as casein [17], whey proteins [18], alfalfa leaf [19], etc. Furthermore, UF and NF are considered to be the best technologies available for the enrichment of bioactive peptides at commercial level. However, limited selectivity (cannot separate peptides having the same molecular size but different charge), and occurrence of concentration polarization and membrane fouling resulting in the declination of permeate flux and the modification of membrane transport selectivity are the major limitation associated with these technologies [15]. To address these inconveniences of pressure driven membrane processes, electrically driven membrane separation processes such as electromembrane filtration (EMF) and electrophoretic membrane contactor (EMC) were introduced. Nevertheless, requirement of high electric field to obtain higher rate of peptides transport and low performance due to strong electrostatic repulsion occurring between peptide and membrane at their interface are the major limitations of EMF and EMC, respectively [20,21]. Therefore, a recently developed electromembrane process, called electrodialysis with filtration membranes (EDFM), which was patented by Bazinet et al. [22], has gained popularity in the field of charged molecules separation because of their better selectivity and less or minimum fouling compared to UF or NF, and less energy requirement unlike EMF.
EDFM consists of an electrodialysis (ED) cell and filtration membranes (stacked between ion exchange membranes) as the major components. Therefore, peptide separation is basically due to the combined effect of charge selectivity of ED cell and size exclusion capabilities of filtration membranes (FMs). Furthermore, it is noteworthy to mention that the applied electric field strength is the only driving force for peptide migration and no transmembrane pressure is applied during the process [11]. This process is considered as the
3
major discovery for peptides separation, thanks to its promising performances such as high selectivity (can separate peptides having same molecular size but different charge) and obtention of peptides with improved bioactivities [11,23]. Within a decade and half, this process has been successfully used at the lab and semi-pilot scale to separate and concentrate bioactive peptides from a wide source of food protein hydrolysates such as β-lg, soy, whey protein, flaxseed, alfalfa white protein, fish, etc. [11,24]. But this process has not yet commercialized due to unavailability of EDFM cell at commercial scale. To commercialize this process, it is paramount to have a clear understanding of the parameters that affect process performances (such as peptide migration and selectivity), so that EDFM cell can be designed and manufactured accordingly. The major parameters that significantly impact EDFM process and its performances are physicochemical characteristics of feed solution (pH, conductivity and concentration), operating conditions of the EDFM system (electric field strength and feed flow rate), and nature and physicochemical properties of filtration membranes. All these parameters interact and influence each other, resulting in a complex system of EDFM process. Up to our knowledge, research works have been carried out by manipulating the first two sets of parameters [25–29]. However a very limited consideration has been given to comprehend the impact of physicochemical properties of FMs on EDFM performances.
During EDFM, charged peptide migrates through FM to its respective anionic or cationic peptide recovery compartments. Therefore, the type of interactions occurring between FM and peptide at the interface, due to their physicochemical properties, must have considerable impact on EDFM performance. Consequently, this study was aimed at investigating the major FM properties that significantly affect peptide migration and selectivity during EDFM by using statistical analyses: redundancy analysis (RDA) and multivariate regression models, and understanding the mechanisms involved. Thus, the FMs tested in this study were characterized in terms of their surface charge, conductivity, hydrophilic/hydrophobic nature of surface and pores, thickness, roughness, porosity, and pore size distribution. Furthermore, a well-characterized whey protein hydrolysate was used as the model feed solution, so that different types of peptide-membrane interactions occurred at the interface, either facilitating
4
or hindering peptide migration and selectivity, and mechanisms behind them could be understood/explained.
In this doctoral thesis, the first part will be a literature review divided into four sections: the first section will be about bioactive peptides obtained by the tryptic hydrolysis of two major whey proteins (β-lactoglobulin and α-lactalbumin), which were used as feed in this research work, the second section will discuss different existing methods for the separation and purification of bioactive peptides, the third section will present a brief overview on various parameters that affect EDFM process with the main focus on physicochemical properties of FMs and finally, the last section provides a quick summary of the advanced statistical analyses used in this research. The literature review will be followed by problematic, hypothesis and main objectives. Then, the results obtained in this research work will be presented in the following three chapters. Finally, the general discussion, conclusion and perspective of the research project will be presented.
5
6
1.1 Bioactive peptides
1.1.1 Generalities
Bioactive peptides (BPs) are short specific protein fragments, which are derived from food proteins. They remain inactive in precursor protein sequences, but when released by peptidase (trypsin, pepsin, thermolysin, etc.) enzymatic hydrolysis, they play active roles in physiological body functions in addition to their nutritional values. The size of BPs usually varies from 3 to 20 amino acids [5]. Upon consumption, according to the amino acid composition and sequence, BPs exhibit a wide range of bioactivities such as antimicrobial, opioid, immune-modulatory, antihypertensive, antithrombotic, mineral binding, antioxidative, agonistic, prebiotic and hypocholesterolemic effects, on human body systems (cardiovascular, immune, gastrointestinal, and nervous system). Few examples of BPs include lactostatin (Ala-Leu-Pro-Met-His (f142-146), and Ile-Ile-Ala-Glu-Lys (f71-75) obtained by enzymatic hydrolysis (using trypsin enzyme) of β-lactoglobulin (β-lg), a whey protein, showed hypocholesterolemic properties [30], M6 peptide (Lys-Arg-Val-Ile-Thr-Tyr) obtained by pepsin treatment of porcine myosin B, exhibited antihypertensive effect [2], α-lactalbumin (α-la) derived peptide namely Trp-Leu-Ala-His-Lys f(104-108) released by trypsin hydrolysis is known to possess angiotensin-converting enzyme (ACE) inhibitory activity, etc. [3]. Many BPs have been found to possess multifunctional bioactivities, which means that a specific peptide sequence shows two or more different biological activities. For example, α-Lactorphin, obtained by pepsin hydrolysis of bovine milk protein (α-la) shows opioid, and ACE-inhibitory effects [31]. In addition, contrary to synthetic molecules, which have been found to have health risk and toxicity, food-derived peptides are considered to be safer and have lower secondary effects to humans [2]. Hence, the search for food-derived bioactive peptides has increased dramatically in recent years.
Bioactive peptides can be obtained from a wide range of plant and animal protein sources such as eggs, milk, meat, fish, wheat, rice, sunflower, flaxseed soy, pulses (lentil, chickpea, beans, and peas), etc. By far, milk proteins represent the most important source of BPs and
7
hence, widely studied. Consequently, in recent years, BPs derived from milk have grabbed the attention because of their potential application in the formulation of physiologically beneficial nutraceuticals, health promoting functional foods or other pharmaceutical products. Among milk-derived peptides, there is an increased interest in BPs obtained by the hydrolysis of whey proteins as they exhibit a wide array of bioactivities that affects cardiovascular, immune and nervous systems. In the context of this research, peptides obtained from the tryptic hydrolysis of whey proteins mainly β-lactoglobulin and α-lactalbumin will be discussed in the following section.
1.1.2 Bioactive peptides from whey proteins
1.1.2.1 Generalities
Whey proteins are one of the major bovine milk proteins representing about 20% of total milk proteins. They comprise of six main proteins: β-lactoglobulin (β-lg) and α-Lactalbumin (α-la) in a high concentration (~80% of whey protein), and proteins such as serum albumin, immunoglobulins, lactoferrin, and a peptide fraction called proteose-peptones in a small quantity [32]. In the nutritional front, whey proteins are proved to have exceptional nutritional quality among all plant and animal-based food protein including another major milk protein, i.e., casein (represents 80% of total milk proteins). Indeed, whey proteins are rich in human-essential amino acids (AAs) such as lysine, methionine, and tryptophan, which cannot be produced by human body itself. In addition, they also contain branched chain AAs (leucine, isoleucine, and valine) that plays vital roles in metabolism, blood glucose homeostasis and neural function. Furthermore, they are an important source of sulfur-containing AAs such as cysteine and methionine, that plays potent roles in enhancing immune functions and one-carbon metabolism [32]. Peptides obtained by the enzymatic hydrolysis of whey proteins have demonstrated a wide range of bioactivities such as antihypertensive, antimicrobial, antioxidative, opiate, and immunomodulatory [33]. Furthermore, in the last two decades, several functional foods formulated with the use of whey-derived bioactive peptides as ingredients/supplements has been commercialized in the international market [34].
8
Whey proteins are well-characterized in terms of peptide sequences. In the context of this research project, only the peptides derived from β-lg and α-la by tryptic hydrolysis will be further discussed, since they are taken as the model protein hydrolysate during electrodialysis with filtration membrane (EDFM) process.
1.1.2.2 β-Lactoglobulin
1.1.2.2.1 Structure and conformation
β-lg is the most abundant whey protein present in bovine milk. It represents around 12 % of total bovine milk protein and ~55% of whey protein [35,36]. Two genetic variants of β-lg (A and B) are normally present in most breeds of cow at high frequency, and are the most common ones in industrial preparation [37,38]. Both variants are composed of 162 amino acids with a molecular weight of ~18 kDa, an isoelectric point of 5.13 and an isoionic point of 5.35 [39]. The primary structure of Type A β-lg with its amino acid sequence is shown in Figure 1.1. The only difference in the amino acid sequence of variants A and B is the presence of glycine (Gly) and alanine (Ala) at positions 64 and 118 respectively for A against aspartic acid (Asp) and valine (Val) for B. One β-lg molecule consists of five cysteines (Cys): Cys66, Cys106, Cys119, Cys121 and Cys 160, four of them involved in the formation of two disulfide bonds (Cys66-Cys160 and Cys106-Cys119, see Figure 1.1) contributing to the stabilization of its compact globular structure [40] and one cysteine at Cys121 resulting in free sulfhydryl group [37,40]. Therefore, the reduction of these two disulfide bonds following thermal or pressure treatments destabilizes the structure of β-lg.
β-lg is a highly structured protein and contains 43% of β-sheets, 10-15% of α-helices and 47% of unordered structures including turns [36,41]. It comprises of nine strands of β-sheets (A-I) located at different positions: 17-25 (A), 42-48 (B), 54-62 (C), 63-74 (D), 81-86 (E), 89-97 (F), 102-108 (G), 118-123 (H) and 147-150 (I). Eight of them: A-D and E-H are arranged to form antiparallel β-barrel with one more β-Sheet and the COOH tail, respectively, which folds into calyx. The α helix is situated on the surface of the molecule at the position 129-143 [42]. Furthermore, at room temperature, β-lg exists as a dimer (MW,~36 kDa) at the
9
pH values between 5.5 and 7.5 (this includes the pH of natural milk which is 6.7-6.8), as a octamer (MW, ~144 kDa) in the pH range of 3.5 and 5.2, while it dissociates to monomer at pH values below 3.5 and above 7.5 [36,37].
Figure 1.1: Primary structure of β-lg A (Adapted and modified [42])
1.1.2.2.2 Peptides derived from tryptic hydrolysis of β-lg
In its native state, β-lg is very resistant to proteolysis due to its globular structure. However, its susceptibility to enzymatic hydrolysis could be improved after its denaturation by means of temperature, pressure, etc. Hydrolysis of β-lg has been predominantly carried out with the enzyme, trypsin, which may contain chymotrypsin in a lesser amount [43,44]. Trypsin generally cleaves at the terminal side of lysine and arginine, and chymotrypsin at the C-terminal side of leucine, phenylalanine, tyrosine, and tryptophan. Table 1.1 presents the
10
peptide sequences obtained by tryptic hydrolysis (containing some amounts of chymotrypsin) of β-lg along with their physicochemical properties such as molecular weight, isoelectric point, global charge at pH 7 and GRAVY (grand average of hydropathy index) score. GRAVY score qualifies and quantifies the hydrophobic or hydrophilic character of each peptide. A positive GRAVY score indicates that the peptide is globally hydrophobic, whereas a negative GRAVY score represents a globally hydrophilic peptide.
11
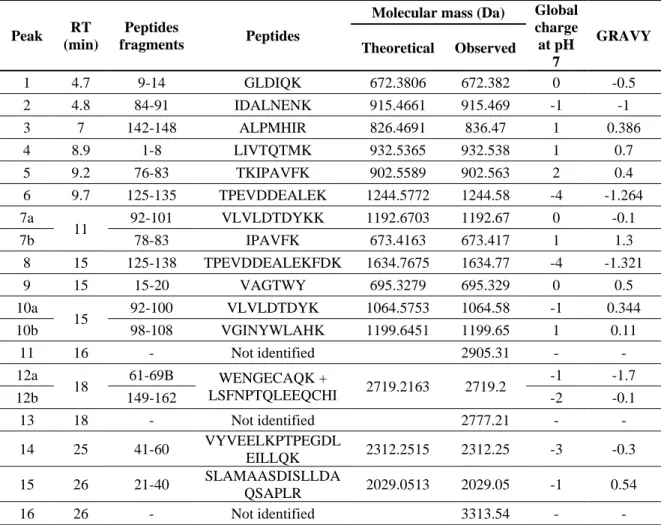
Table 1.1: Peptides derived from the tryptic hydrolysis of β-lg (Adapted and modified from [45,46])
No. Peptides
fragment Amino acids sequence
Molecular Weight pI Global charge at pH 7 GRAVY score 1 1–8 LIVTQTMK 932.5 8.75 1 0.7 2 9–14 GLDIQK 672.3 5.84 0 0.1 3 15–20 VAGTWY 695.3 5.49 0 0.5 4 21–32 SLAMAASDISLL 1190.7 3.8 -1 1.4 5 33–39 DAQSAPL 700.3 3.8 -1 -0.3 6 33–40 DAQSAPLR 856.4 5.84 0 -0.8 7 41–42 VY 280.2 5.49 0 - 8 41–57 VYVEELKPTPEGDLEIL 1943.8 3.91 -0.2 21-40 SLAMAASDISLLDAQSAPLR 2029.7 4.21 -1 0.5 9 41–60 VYVEELKPTPEGDLEILLQK 2313.2 4.25 -3 -0.3 10 43–57 VEELKPTPEGDLEILL 1795 3.91 -4 -0.1 11 43–60 VEELKPTPEGDLEILLQK 2050.9 4.25 -3 -0.5 12 61-70A + 61–69A WENDAECAQKK + WENDAECAQK 2125 4.25 -1 -2 -2.1 61-70B + 61–69B WENGBECAQKK + WENGBECAQK 2400 4.6 0 -1 -1.7 13 61–70A + 149–162 WENDAECAQKK + LSFNPTQLEEQCHI 2906.5A 4.57 -1 -2 61–70B + 149–162 WENGBECAQKK + LSFNPTQLEEQCHI 2848.7B 4.91 0 -2 14 61–69A+ 149–162 WENDAECAQK + LSFNPTQLEEQCHI 2778.3A 4.25 -2, -2 -1.7 61–69B + 149–162 WENGBECAQK + LSFNPTQLEEQCHI 2720.4B 4.48 -1, -2 -0.1 15 71–75 IIAEK 572.3 6 0 0.7 16 76–82 TKIPAVF 774.5 8.41 1 1 17 78–82 IPAVF 545.3 5.52 0 2.3 18 83–91 KIDALNENK 1043.7 6.07 0 -1.3 19 84–91 IDALNENK 915.4 4.37 -1 -1 20 92–101 VLVLDTDYKK 1192.8 5.93 0 -0.1 21 125–135 TPEVDDEALEK 1244.8 3.83 -4 -1.3 22 125–138 TPEVDDEALEKFDK 1635.2 4.02 -4 -1.3 23 136–138 FDK 408.2 5.84 0 -1.5 24 142–148 ALPMHIR 836.4 9.8 1 0.4 25 146–148 HIR 424.5 9.76 2 -1.1 26 149–162 + 149–162 LSFNPTQLEEQCHI + LSFNPTQLEEQCHI 3315 4.47 -0.5 27 104–124 LFCMENSAEPEQSLACQCLVR 2369.8 4.25 0.13
No. 8 exhibits coelution of two peptides (have same retention time). The letters A and B in subscript represents variants A and B of β-lg. The + sign in between two peptide sequences (peaks 12, 13, 14 and 26) indicated the presence of a disulfide bond between them and hence, they are inseparable.
12
1.1.2.2.3 Bioactivities of β-lg derived peptides
The bioactivities of peptides obtained from the hydrolysis of β-lg using different enzymes such as trypsin, chymotrypsin, pepsin, thermolysin, etc. have been widely studied. They have been found to possess potential physiological and regulatory functions in the human body. Table 1.2 listed the peptides with their observed bioactivities, obtained by hydrolysis of β-lg using different enzymes.
Table 1.2 :Bioactive peptides obtained by the enzymatic hydrolysis of bovine β-lg (Adapted and updated from [47,48])
Bioactivities Peptide fragment Enzymes used
Antihypertensive
ALPMHIR, LAMA, LDAQSAPLR, VFK Trypsin VAGTWY, IIAEK, IPAVFK, GLDIQK, YPFPGPI Trypsin+Pepsin
CMENSA, ALPMH, VLDTDYK, VAGTW Pepsin, then Trypsin+Chymotrypsin Antidiabetic
VAGTWY, IPAVFK, IPAVF, TPEVDDEALEK,
VLVLDTDYK Trypsin
LKPTPEGDL, LKPTPEGDLEIL, IPAVFKIDA Pepsin
Opioid YLLF Trypsin
Antimicrobial
VAGTWY, AASDISLLDAQSAPLR, IPAVFK,
VLVLDTDYK Bovine trypsin
TPEVDDEALEK Trypsin+ Chymotrypsin
KVAGT, VRT, IRL, PEGDL, LPMH, EKF Porcine pepsin
Hypocholesterolemic IIAEK, ALPMH, GLNIQK Trpsin
Antioxidant
MHIRL, YVEEL, WYSLAMAASDI, FNOTQ,
LQKW, LDTYKK Thermolysin
VAGTWY Trypsin
WYSL Alcalase
1.1.2.3 α-lactalbumin
1.1.2.3.1 Structure and conformation
α-Lactalbumin (α-la) is the second major whey protein present in bovine milk. It represents 3.5% of total milk protein and 20% of whey protein. It is a globular protein. It consists of 123 amino acids, has a molecular weight of 14.2 kDa, an isoelectric point of 4.2 and an isoionic point of 4.5 [49]. It is a good source of essential amino acids: tryptophan and cysteine. There are two predominant genetic variants in α-la; A and B, B being the most
13
abundant one. The variant A differs from B with a single amino acid in the tenth position consisting of an arginine (variant A) instead of a glutamine (variant B). It consists of eight cysteines that form four intramolecular disulfide bonds (Cys6–Cys120, Cys61–Cys77, Cy73–Cys91, and Cys28–Cys111), which stabilizes its globular structure. The primary structure of Type A α-la with its amino acid sequences is shown in Figure 1.2.
The structure of α-la is divided into two domains: α-domain (accounts for 44% of total structure), which is largely helical, and β-domain (accounts for 11% of total structure) that consists of β-sheets (residue 35-85). The α-domain comprises of four helices at different positions: 5-11, 23-24, 86-98 and 105-109 and domain composed of three antiparallel β-sheets at positions 41-44, 47-50 and 55-56.
14 Figure 1.2: Primary structure of α-la A (Adapted from [49])
15
1.1.2.3.2 Peptides derived from tryptic hydrolysis of α-la
Table 1.3 listed the peptides obtained by the tryptic hydrolysis of α-la.
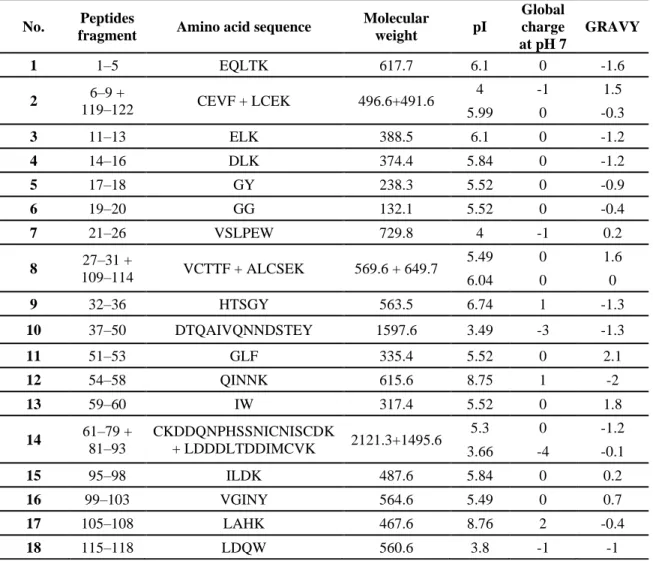
Table 1.3: Peptides derived from the enzymatic hydrolysis of α-la (Adapted and updated from [46])
No. Peptides
fragment Amino acid sequence
Molecular weight pI Global charge at pH 7 GRAVY 1 1–5 EQLTK 617.7 6.1 0 -1.6 2 6–9 + 119–122 CEVF + LCEK 496.6+491.6 4 -1 1.5 5.99 0 -0.3 3 11–13 ELK 388.5 6.1 0 -1.2 4 14–16 DLK 374.4 5.84 0 -1.2 5 17–18 GY 238.3 5.52 0 -0.9 6 19–20 GG 132.1 5.52 0 -0.4 7 21–26 VSLPEW 729.8 4 -1 0.2 8 27–31 + 109–114 VCTTF + ALCSEK 569.6 + 649.7 5.49 0 1.6 6.04 0 0 9 32–36 HTSGY 563.5 6.74 1 -1.3 10 37–50 DTQAIVQNNDSTEY 1597.6 3.49 -3 -1.3 11 51–53 GLF 335.4 5.52 0 2.1 12 54–58 QINNK 615.6 8.75 1 -2 13 59–60 IW 317.4 5.52 0 1.8 14 61–79 + 81–93 CKDDQNPHSSNICNISCDK + LDDDLTDDIMCVK 2121.3+1495.6 5.3 0 -1.2 3.66 -4 -0.1 15 95–98 ILDK 487.6 5.84 0 0.2 16 99–103 VGINY 564.6 5.49 0 0.7 17 105–108 LAHK 467.6 8.76 2 -0.4 18 115–118 LDQW 560.6 3.8 -1 -1
The + sign in between two peptide sequences (No. 2, 8 and 14) indicated the presence of a disulfide bond between them and hence, they are inseparable.
1.1.2.3.3 Bioactivities of α-la derived peptides
Peptides obtained by the hydrolysis of α-la have been found to possess a wide range of bioactivities. Table 1.4 presents the peptides derived from enzymatic hydrolysis of α-la using different enzymes with their resulting bioactivities.
16
Table 1.4: Bioactive peptides obtained by enzymatic hydrolysis of bovine α-la (Adapted and updated from [47,48])
Bioactivities Peptide fragment Enzymes used
Antihypertensive
VGINYWLAHK, FFVAPFPEVFGK, YGL Trypsin+Pepsin+Chymotrypsin
LAHKAL Pepsin+Trypsin
WLAHK, VGINYWLAHK Trypsin
Antioxidant WYSLAMAASDI Corolase PP
Antomicrobial
EQLTK. GYGGVSLPEWVCTTF + ALCSEK Porcine trypsin CKDDQNPH+ISCDKF Bovine chymotrypsin
Antidiabetic
WLAHKAL, WLAHKALCSEKLDQ, LAHKALCSEKL, TKCEVFRE, IVQNNDSTEYGLF, ILDKVGINY
Pepsin
Opioid YGLF Pepsin
1.2 Bioactive peptides separation and purification methods
The protein hydrolysate, obtained after protein hydrolysis, contains bioactive peptides in low concentrations, along with other inactive hydrolyzed protein fractions. Therefore, an appropriate and efficient fractionation technique is crucial to concentrate and selectively separate those bioactive peptides, in order to yield the enriched bioactive peptide fractions or a specific bioactive peptide, with improved biological activity and/or nutritional values. Various technologies such as selective precipitation, chromatographic, electrophoresis, and membrane-based methods have already been used successfully at the lab scale [50,51]. Among these, chromatographic techniques and pressure driven membrane processes have been used at industrial scale for the separation and concentration of bioactive peptides. These methods separate peptides based on one or more physicochemical property such as charge, size, polarity or hydrophobicity, etc. of a peptide. Furthermore, a combination of two or more technologies is desired to achieve the separation of a specific peptide with a high degree of purity. For instance, membrane-based methods are generally combined with other technologies such as chromatography, to fractionate and isolate a specific bioactive peptide from the crude hydrolysate at a pilot scale production. However, the commercial scale
17
production of enriched and/or pure bioactive peptide is still challenging due to the lack of suitable large-scale technologies.
1.2.1 Chromatographic techniques
1.2.1.1 Generalities
Chromatographic techniques separate peptides based on their differential affinity to mobile phase (liquid or gas) and stationary phase (liquid or solid) according to their physicochemical properties. Therefore, peptide having high affinity to mobile phase moves with a high velocity through chromatographic column and becomes the first one to be eluted. Conversely, peptide having high affinity to stationary phase moves with a low velocity and becomes the last one to be eluted through the column. Consequently, this difference in migration velocity results in separation of respective peptides from a complex mixture [52].
Different chromatographic techniques that have been commonly used in recent years for peptides fractionation and purification are reversed phase high performance liquid chromatography, ion-exchange chromatography, size exclusion chromatography and fast protein liquid chromatography. Each of these techniques separate peptides based on their physicochemical properties such as molecular weight, charge and polarity or hydrophobicity. In terms of separation effects, they are efficient in selectively fractionating peptide of interest. However, prior to any chromatographic experiments, membrane filtration, centrifugation or sieving of protein hydrolysate is generally required.
1.2.1.2 Reversed phase high performance liquid chromatography
Since 1970’s, reversed phase high performance liquid chromatography (RP-HPLC), has been extensively used for fractionation and purification of peptides. It separates peptides based on their polarity. It consists of hydrophobic stationary phase/column, which is silica-based with non-polar carbon grafted on it, and polar mobile phase, which is generally the blend of organic and aqueous solvents (usually water and methanol or acetonitrile) [53,54]. When a
![Table 1.2 :Bioactive peptides obtained by the enzymatic hydrolysis of bovine β-lg (Adapted and updated from [47,48])](https://thumb-eu.123doks.com/thumbv2/123doknet/2914954.75959/40.918.122.786.390.756/table-bioactive-peptides-obtained-enzymatic-hydrolysis-adapted-updated.webp)
![Table 1.4: Bioactive peptides obtained by enzymatic hydrolysis of bovine α-la (Adapted and updated from [47,48])](https://thumb-eu.123doks.com/thumbv2/123doknet/2914954.75959/44.918.124.779.160.451/table-bioactive-peptides-obtained-enzymatic-hydrolysis-adapted-updated.webp)
![Figure 1.4: Schematic representation of concentration polarization and fouling during pressure driven membrane separation processes (Adapted and modified from [46])](https://thumb-eu.123doks.com/thumbv2/123doknet/2914954.75959/53.918.144.707.198.533/schematic-representation-concentration-polarization-pressure-membrane-separation-processes.webp)
![Figure 1.14: Schematic representation of zeta potential of filtration membranes (Adapted and modified from [118])](https://thumb-eu.123doks.com/thumbv2/123doknet/2914954.75959/76.918.132.738.105.637/figure-schematic-representation-potential-filtration-membranes-adapted-modified.webp)