THÈSE
Pour l'obtention du grade de
DOCTEUR DE L'UNIVERSITÉ DE POITIERS UFR des sciences fondamentales et appliquées
Unité de recherche pluridisciplinaire prairies et plantes fourragères - URP3F (Diplôme National - Arrêté du 7 août 2006)
École doctorale : Sciences pour l'environnement - Gay Lussac (La Rochelle) Secteur de recherche : Agronomie, production animales et végétales, agro-alimentaire
Présentée par : Amel Maamouri
Variabilité génétique de la luzerne cultivée en association avec une graminée fourragère
Directeur(s) de Thèse : Bernadette Julier
Soutenue le 26 juin 2014 devant le jury
Jury :
Président Rémi Lemoine Directeur de recherche, CNRS, Université de Poitiers Rapporteur Béatrice Denoyes-Rothan Ingénieur de recherche, INRA Aquitaine
Rapporteur Joëlle Fustec Enseignante-chercheuse, ESA, Angers Membre Bernadette Julier Directrice de recherche, INRA de Lusignan Membre Sandrine Flajoulot Ingénieur de recherche, Jouffray-Drillaud
Membre Gérard Duc Directeur de recherche, INRA de Dijon
Pour citer cette thèse :
Amel Maamouri. Variabilité génétique de la luzerne cultivée en association avec une graminée fourragère [En ligne]. Thèse Agronomie, production animales et végétales, agro-alimentaire. Poitiers : Université de Poitiers, 2014. Disponible sur Internet <http://theses.univ-poitiers.fr>
1
THESE
pour l’obtention du Grade de
DOCTEUR DE L’UNIVERSITE DE POITIERS (Faculté des Sciences Fondamentales et Appliquées)
(Diplôme National - Arrêté du 7 août 2006)
Ecole Doctorale : Sciences pour l'environnement - Gay Lussac
Secteur de Recherche : Agronomie, production animales et végétales, agro-alimentaire
Présentée par :
Amel MAAMOURI
************************Variabilité génétique de la luzerne cultivée en association avec une
graminée fourragère
************************
Préparée à l’Unité de Recherche Pluridisciplinaire Prairies et Plantes Fourragères INRA, 86600 LUSIGNAN
Bourse cofinancée par l’INRA et la région Poitou-Charentes Directrice de thèse : Bernadette JULIER
************************
Soutenue le 26 juin 2014 devant la Commission d’Examen ************************
JURY
M Rémi LEMOINE Professeur, Université de Poitiers Président Mme Béatrice DENOYES Ingénieur de Recherche, INRA Rapporteure Mme Joëlle FUSTEC Enseignante-Chercheuse, Groupe ESA Rapporteure Mme Bernadette JULIER Directrice de recherche, INRA Examinatrice Mme Sandrine FLAJOULOT Ingénieur de Recherche, Jouffray-Drillaud Examinatrice M Gérard DUC Directeur de Recherche, INRA Examinateur
3
!
"
#
#
#
#
$
$
$
$
% ## &
% ## &
% ## &
% ## &
5
Remerciements
La liste des remerciements qui va suivre se veut la plus exhaustive possible mais il n’y aura jamais suffisamment de place pour remercier tout le monde dans le détail, je m’excuse alors d’avance auprès de tous celles ou ceux que j’aurais omis et à qui j’exprime toute ma gratitude.
Je remercie tout d’abord Bernadette Julier pour m’avoir fait confiance et m’avoir permis de réaliser ces trois années de thèse qui m’ont enrichie et marqué une période de ma vie que rien ne pourra remplacer. Merci Bernadette de m’avoir donné l’occasion et encouragée à valoriser mes travaux et participer à un congrès à l’autre bout du monde. Merci pour toute l’aide que tu m’as apportée à mon arrivée, pour m’installer et après pour déménager. Merci de m’avoir orientée quand j’ai commencé à me perdre, merci pour ton soutien permanent et tes discussions fructueuses. Merci d’être toujours prête à répondre à mes questions même à distance. Je ne te remercierai jamais assez pour tout ce que tu as fait pour moi.
Mais aussi merci de m’avoir ouvert les portes de l’Unité de Recherche Pluridisciplinaire Prairies et Plantes Fourragères de Lusignan où j’ai rencontré des personnes sympas et connu des amis pour la vie. J’ai trouvé en ces personnes mes proches qui m’ont toujours manqué et étaient loin de moi sur un autre continent. J’ai trouvé en vous le père (Jojo et Philou ; tous les deux toujours de bonne humeur), la mère (Marilyn, Claudine et Françoise), les frères (Jean François, David, Philippe, Sébastien, Thierry, Rodrigue et Fabien), les sœurs (Bernadette, Amandine, Sabrina, Sandrine, Nathalie, Liliane, Isabelle et Magali) que j’ai beaucoup aimé et avec qui j’ai partagé des moments inoubliables. Je remercie la grande famille au sein de laquelle je me suis rapidement intégrée j’ai passé les plus belles années de ma vie et je ne me suis jamais sentie étrangère. Sans vous la réalisation de cette thèse aurait été beaucoup plus difficile.
Je remercie également Gaëtan Louarn avec qui j’ai travaillé sur l’une des expérimentations. Je te remercie pour les conseils pertinents et les discussions enrichissantes qui ont fait avancer les travaux.
Je tiens à remercier Béatrice Denoyes et Joelle Fustec d’avoir accepté d’être rapporteurs de cette thèse. Je remercie également Sandrine Flajoulot et Gérard Duc pour leur rôle d’examinateur et Rémi Lemoine de présider le jury.
Je remercie tous les membres de mes comités de thèse : Judith Burstin, Joelle Fustec, Sylvie Marhadour, Gaëtan Louarn et Vincent Béguier qui m’ont apporté leurs avis critiques et leurs conseils avisés dans l’orientation de cette thèse.
Mes remerciements sincères à Marc Chabosseau, Lilian Giry, François Gastal, Jean-Louis Durand, Françoise Macouin et Nathalie Bonnet pour toutes les décisions prises et les démarches administratives effectuées pour que je puisse mener à terme cette thèse.
Un très grand merci pour Joël Jousse « expert luzerne» responsable technique des essais (compétiluz et cartoluz). Merci Jojo pour ta rigueur et ta patience, pour les journées passées à entretenir les essais, à repérer la plante cible durant la récolte... Merci Jojo pour toutes les discussions et les conseils techniques sur la luzerne que tu as partagés avec moi. Merci d’être si agréable et si gentil.
Un très grand merci à Marilyn Vandier qui m’a accompagnée pendant ces trois ans et qui était très proche de moi. Nous nous sommes raconté beaucoup d’histoires. Merci d’avoir été là pour moi pour me soutenir, pour m’aider dans toutes les manips en plein champ quand j’étais couché par terre sous le soleil et la pluie pour faire les mesures sur les plantes. Merci pour toutes les discussions personnelles que nous avons eues. Je n’oublierai jamais le premier jour où je t’ai rencontré, j’ai senti qu’on allait s’entendre. Malgré ta timidité et ta discrétion, j’ai trouvé en toi une personne très mûre, très serviable et très responsable. Je ne te remercierai jamais assez pour tout ce que tu as fait pour moi, pour tous les services que tu m’as rendus.
Un très grand merci à Jean-François qui a contribué aux différentes manips au champ. Merci pour les batailles que nous avons menées et où je t’ai battu. Je me défoulais sur toi (t'étais mon punching ball) et Marylin en est témoin. Vraiment, merci de m’avoir supportée pendant ces trois ans grand frère. Nos batailles vont me manquer énormément. Merci de m’avoir dépannée quand les ordinateurs se retournaient contre moi et merci aussi à tes collègues informatiques Sébastien Pommiès et Jean-Marie Girardeau.
Un spécial remerciement pour Magali Caillaud qui m’a accueillie chez elle et hébergée pendant les derniers mois les plus critiques. Je ne te remercierai jamais assez, Mag, merci d’être si agréable avec moi. Je me suis sentie comme chez moi. Merci pour les fous rires et ta bonne humeur. Merci pour tes encouragements (et le compte à rebours, ça m’a bien fait avancer surtout que t’oubliais un ou deux jours lors du comptage !!!). Prends soin de toi et de Inata, vous allez me manquer.
Un spécial remerciement à Amandine Vogt, une vraie sœur, merci pour ton soutien moral et tes encouragements et toutes les discussions que nous avons eues en courant, c’était fatiguant mais indispensable pour moi. Merci de m’avoir écoutée. Tout ça va me manquer.
Je ne remercierai jamais assez tout le personnel technique de l’équipe génétique de l’unité qui de prêt ou de loin a contribué à la réalisation de cette thèse et a participé aux manips en plein champ des deux essais compétiluz et cartoluz : Joël, Marilyn, Jean-François, Claudine, Rodrigue, Fabien, Magali, David, Sébastien, Jérôme, Brigitte, Franck, Bernard, Pascal, Dominique. Merci pour votre patience lors des récoltes et la réalisation des manips minutieuses du comptage et de dépiautage des plantes de luzerne. Nous avons formé une armée qui a bien travaillé dans une bonne ambiance.
Un grand merci à l’équipe écophysiologie de l’unité avec qui nous avons suivi simultanément l’essai au champ de compétiluz dans une parfaite harmonie d’organisation : un très grand merci à Jean-Pierre Terrasson et Eric Rivault qui m’ont aidée dans les manips que nous avons effectuées en commun. Merci à Annie Epinchard de nous avoir aidés aussi.
7
Mes remerciements sincères s’adressent à toutes les personnes du laboratoire de biologie moléculaire de l’INRA de Lusignan qui m’ont accompagnée pendant les manips : Denise Cadier, Sabrina Delaunay, Philippe Cormenier pour leur implication dans les travaux de la partie cartographie, Françoise Durand, Sébastien Blugeon et Céline Bernier pour les nombreux échanges sur nos manips respectives et le fonctionnement du laboratoire.
J’adresse mes remerciements aussi à l’équipe du laboratoire chimie de l’INRA de Lusignan : Catherine Levèque pour avoir passé tous les échantillons de compétiluz au NIRS et fourni les résultats à temps, Corinne Melin et Nathalie Moynet d’avoir passé les échantillons de compétiluz sur l’analyseur d’azote, Véronique Menanteau. Je vous remercie Mesdames pour avoir participé aussi aux récoltes de compétiluz. Merci à François Gastal pour la coordination des analyses d’azote et son implication dans le fonctionnement de l’analyseur d’azote et à Philippe Barre pour le suivi des équations NIRS.
Merci aux scientifiques Philippe Barre, François Gastal, Isabelle Litrico, Marc Ghesquière, Jean-Paul Sampoux pour les discussions enrichissantes.
Je tiens également à remercier Nathalie Bonnet, Isabelle Boissou, Geneviève Pierre, Liliane Jean et Fabrice Domineau de gérer à la perfection la partie administrative. Merci à Nathalie, Liliane et Fabrice d’être si agréables et présentes dans les moments les plus difficiles.
Merci aux étudiants et stagiaires, Héléna, Aurélie, Khadija, Adrian, Arianne et Jonathan, qui ont permis de faire avancer ces travaux de thèse.
Merci à Sabrina, Stéphane, Julien, Laurence, Gaëlle (Zouza), Magali, Eva, Stéphane, David, Taisiia pour les apéros qu’on se faisait entre nous. Merci Sabrina pour tes sms pour me rappeler que nous avons cours de gym tous les mardis soir et merci pour ton soutien moral.
Merci à tous les autres stagiaires que j’ai pu rencontrer au cours de cette belle aventure : Ludovic, Monserrat, Florian, Mart, Sophie, Tiago, Laura, Fleur … et Vincent ! Bon courage pour ta thèse.
Merci à l’équipe de Volley de l’INRA, Aurélie, Florence, Helena, Agathe, Eric, David, Gaëtan, Didier, Christophe, Fabien, Jean-François, Abraham et Tiago. Merci aussi à l’équipe de course à pied, Pascal, Amandine, Isabelle, Bernadette, Carine, Christophe, Daniel et Philippe. Merci pour les moments sympas que nous avons passés ensemble.
Je tiens à remercier Patrick Despeignes qui m’a beaucoup dépannée pour ma Twingo (DDE orange) que je tiens à remercier également pour ne pas m’avoir lâchée au cours de ces trois ans. Merci Patrick pour ta bonne humeur. Merci également à Jérôme Fillon.
Merci à Sabrina Biais et Sylvie Perez responsables à l’école doctorale Gay Lussac de l’Université de Poitiers qui ont réalisé le suivi administratif de ma thèse.
Merci à ma famille en France, ma tante Latifa et ses enfants Akram, Insaf et Anis, qui m’a beaucoup encouragée et avec qui j’ai passé de très bons moments gravés à jamais. Tati, je n’oublierai jamais les heures passées au téléphone, ni les nuits blanches lors des vacances et
merci pour tous les bons plats que tu m’as préparés c’était un régal. Merci grand frère Kam, merci sœurette Nassouf, merci petit frère Nanou merci pour tout ce que nous avons partagé ensemble.
Merci à mes parents qui de loin ont toujours eu confiance en moi, supportée, encouragée à avancer. Merci de m’avoir appris à être patiente, d’aller jusqu’au bout de mes projets. Merci pour l’éducation que vous m’avez offerte qui m’a permis de s’intégrer facilement, d’apprendre à vivre loin de vous et de mes frères. Je suis qui je suis aujourd'hui grâce à vous. Merci à mes frères qui étaient toujours là pour moi quand j’avais besoin d’eux. Sans vous je ne serais jamais arrivée où je suis.
Pour boucler la boucle, merci à mon cher mari sur qui j’ai piqué mes crises de colère. Merci d’avoir su me supporter et me redonner courage lors de différents passages à vide. Merci d’avoir été présent pour moi.
9
Liste des abréviations
ADN: Acide DésoxyriboNucléique
ACP : Analyse en Composantes Principales
ACVF : Association des Créateurs de Variétés Fourragères
AFLP : Amplified Fragment Length Polymorphism
C: Carbon
CAPS : Cleaved Amplified Polymorphic Sequence
CIRAD : Centre de coopération Internationale en Recherche Agronomique pour le Développement cM : centiMorgan
DArT : Diversity Array Technology
DHS : Distinction, Homogénéité, Stabilité
EST : Expressed Sequence Tag
GEVES : Groupe d'Etude et de contrôle des Variétés et des Semences GL : Groupe de Liaison
HRM : High Resolution Melting
INN : Indice de Nutrition azotée
INRA : Institut National de Recherche Agronomique LOD : Logarithm of the odds ratio
N : Nitrogen
NF : Non Fixateur
Nl : concentration en azote des feuilles éclairées PCR : Polymerase Chain Reaction
QTL : Quantitive Trait Loci
RADP : Randomly Amplified Polymorphism DNA
RFLP: Restriction Fragment Length Polymorphism RIL : Recombinant Inbred Line
SC : Somme des Carrés
SNP : Single Nucleotide Polymorphism
SPIR : Specrométrie Proche InfraRouge SSR : Simple Sequence Repeats
11
Table des matières
1 Introduction ... 15
2 Synthèse bibliographique ... 19
2.1 Les associations légumineuses-graminées ... 19
2.1.1 Historique et situation actuelle ... 19
2.1.2 Intérêts et attendus des associations ... 20
2.1.3 Fonctionnement de l’association légumineuses-graminées ... 22
2.1.3.1 Partage de la lumière en association ... 23
2.1.3.2 Fixation et transfert de l’azote atmosphérique dans les associations graminées-légumineuses ... 24
2.1.3.3 Compétition pour d’autres éléments : eau, autre minéraux ... 28
2.1.4 Caractères impliqués dans le fonctionnement d’une association ... 28
2.1.5 Choix variétal et maintien des deux espèces ... 30
2.2 Présentation de la luzerne et généralités sur la fétuque élevée ... 31
2.2.1 La luzerne ... 31
2.2.1.1 Origines et expansion géographique ... 31
2.2.1.2 Evolution et taxonomie ... 32
2.2.1.3 Morphologie, physiologie et cycle de développement ... 35
2.2.1.4 Diversité disponible ... 38
2.2.1.5 Importance et types de culture ... 40
2.2.2 La fétuque élevée ... 41
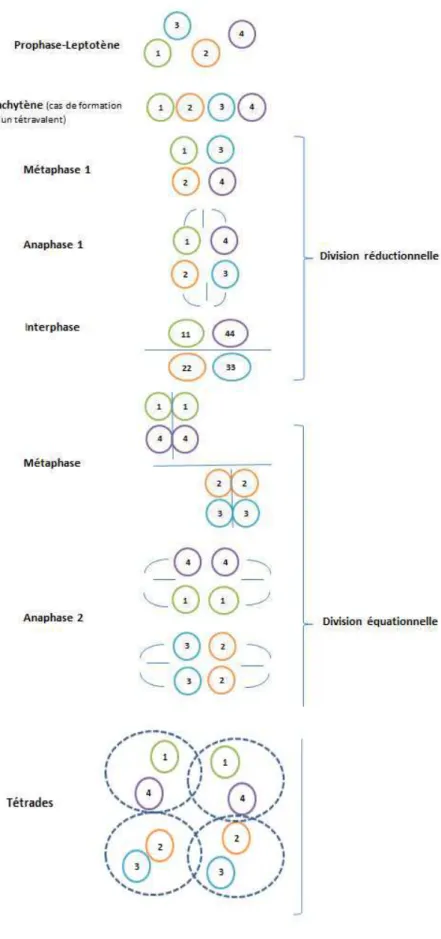
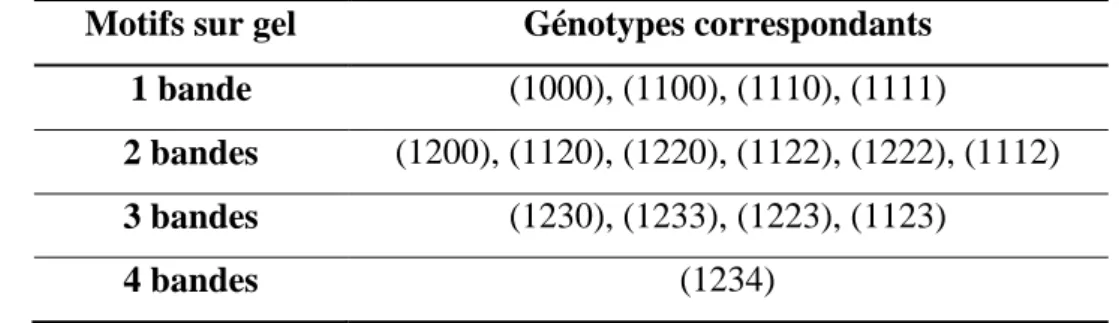
2.3 Recherche de zones du génome impliqués dans le déterminisme d’un caractère quantitatif chez les polyploïdes ... 42
2.3.1 Génétique des tétraploïdes ... 42
2.3.2 Construction des cartes génétiques ... 46
2.3.3 Cartographie de locus à effets quantitatifs ... 50
2.3.4 Recherche de QTL chez le genre Medicago ... 51
3 Matériel et méthodes ... 55
3.1 Caractérisation de la variabilité génétique de la luzerne pour l’aptitude à être cultiver en mélange avec la fétuque élevée ... 55
3.1.1 Matériel végétal ... 55
3.1.2 Notations phénotypiques ... 60
3.1.3 Analyses statistiques ... 62
3.1.3.1 Analyse des deux mélanges de luzerne et de fétuque ... 63
3.1.3.2 Les caractéristiques des génotypes de luzerne en mélange sont-elles les mêmes qu’en monoculture ? ... 64
3.1.3.3 Est-ce que les caractéristiques des génotypes de luzerne en isolé sont prédictifs des caractéristiques des mêmes génotypes en monoculture et en mélange ? 66 3.2 Déterminisme génétique de l’aptitude à la culture en mélange de la luzerne ... 67
3.2.1 Matériel végétal ... 67
3.2.2 Génotypage et cartographie génétique ... 67
3.2.2.1 Génotypage avec les marqueurs SSR ... 68
3.2.2.2 Génotypage avec les marqueurs DArT ... 69
3.2.2.3 Ségrégation des marqueurs ... 69
3.2.2.4 Construction de la carte ... 71
3.2.3 Phénotypage de la population de cartographie ... 72
3.2.3.1 Dispositif expérimental... 72
3.2.3.2 Notations réalisées ... 73
3.2.3.3 Analyses statistiques ... 74
3.2.4 Détection de QTL ... 75
4 Caractérisation de la variabilité génétique de la luzerne pour l’aptitude à être cultivée en mélange avec la fétuque élevée ... 77
4.1 Analyse des deux mélanges de luzerne et de fétuque ... 77
4.1.1 Variabilité génétique des caractères mesurés sur les génotypes de luzerne cultivés en mélange avec la fétuque élevée ... 77
4.1.2 Variation phénotypique des caractères mesurés sur les deux génotypes de la fétuque élevée cultivés en mélange avec la luzerne ... 81
4.1.3 Interaction entre la luzerne et la fétuque élevée ... 86
4.1.4 Discussion ... 88
4.1.4.1 La variabilité génétique des caractères de la luzerne ... 88
4.1.4.2 Variation phénotypique de la fétuque élevée ... 90
4.1.4.3 Interaction entre la luzerne et la fétuque élevée ... 91
4.2 Les caractéristiques des génotypes de luzerne en mélange sont-elles les mêmes qu’en monoculture et peuvent-elles être prédites par les caractéristiques des mêmes génotypes en isolé?.. ... 96
13
4.2.1 Les caractéristiques des génotypes de luzerne en mélange sont-elles les mêmes
qu’en monoculture ? ... 96
4.2.1.1 Variabilité génétique de la luzerne en monoculture ... 96
4.2.1.2 Comparaison des caractéristiques des génotypes de luzerne en mélange avec celles en monoculture ... 100
4.2.2 Est-ce que les caractéristiques des génotypes de luzerne en isolé sont prédictifs des caractéristiques des mêmes génotypes en monoculture et en mélange ? ... 105
4.2.2.1 Variabilité génétique de la luzerne en isolé ... 105
4.2.2.2 Comparaison des caractéristiques des génotypes de luzerne en isolé avec celles en mélange et en monoculture ... 108
4.2.3 Discussion ... 117
4.2.3.1 Variabilité génétique de la luzerne en isolé et en monoculture ... 117
4.2.3.2 Comparaison des caractéristiques des génotypes de luzerne en mélange et en monoculture ... 119
4.2.3.3 Les caractéristiques en isolé prédictives de celles en mélange et en monoculture ... 121
5 Détection de QTL de caractères liés à la culture en association de la luzerne avec la fétuque ... 125
5.1 Réalisation des cartes génétiques ... 125
5.1.1 Ségrégation des marqueurs SSR ... 125
5.1.2 Ajout de marqueurs DArT ... 126
5.2 Analyse des données phénotypiques ... 127
5.2.1 Variabilité génétique de la luzerne dans les différents traitements pour la biomasse et la hauteur ... 127
5.2.2 Analyse de l’effet du génotype de luzerne, du traitement et de l’interaction traitement x génotype de luzerne sur la biomasse et la hauteur ... 129
5.2.3 Analyse de corrélations entre la biomasse et la hauteur dans un même traitement 131 5.2.4 Analyse des corrélations pour la biomasse et la hauteur entre les deux traitements mélange et monoculture ... 132
5.2.5 Analyse des corrélations pour la biomasse et la hauteur entre les traitements isolé et mélange et les traitements isolé et monoculture ... 133
5.3 Détection de QTL ... 136
5.4 Discussion ... 148
5.4.1 Cartographie génétique de la population F1 ... 148
5.4.3 Détection de QTL ... 152 6 Conclusion et perspectives ... 157 Références bibliographiques ... 163 Annexes ... 181 Résumé ... 220 Abstract ... 220
15
1
Introduction
La luzerne cultivée (Medicago sativa) est une légumineuse fourragère pérenne, autotétraploïde et allogame appartenant à la famille des Fabacées qui constitue un taxon végétal de grande importance. Elle est tolérante à la sécheresse et aux hautes températures et est cultivée essentiellement pour une exploitation en fauche, sur des sols non acides et non hydromorphes. Elle est exploitée, depuis les débuts de l’agriculture, pour sa partie aérienne riche en protéines et destinée à l’alimentation animale (Julier et Huyghe, 2010). Grâce à sa capacité à fixer l’azote atmosphérique, la luzerne peut s’adapter aux situations de faibles ressources en azote minéral du sol. Ce qui lui confère un double intérêt économique par une réduction de la fertilisation azotée et par conséquent des engrais azotés de synthèse associée à une production de protéines, et écologique par la limitation de lixiviation des nitrates vers la nappe phréatique.
Ayant développé des mécanismes d’adaptation à la concurrence pour la lumière au niveau de ses parties aériennes, la luzerne est cultivée en association avec des plantes non fixatrices comme les graminées. Les bénéfices des associations légumineuses-graminées, principalement la production fourragère en quantité et en qualité (énergie et protéines) et l’autonomie en azote, sont assurés par le maintien des deux espèces au fil des repousses. Chaque espèce doit donc être apte à être cultivée en association avec l’autre, ce qui signifie que chaque espèce contribue à une part significative de la production et permet la production de l’autre espèce. Elle inclut aussi le maintien des deux espèces associées dans des proportions adéquates pour atteindre les finalités agronomiques (production de biomasse, teneur en protéines), économiques (faibles intrants azotés) et environnementales. Cependant, cette culture est considérée comme un cas particulier de culture en situation de compétition où les effets des espèces sur les ressources induisent une réponse négative chez au moins l’une des deux espèces en présence (Louarn et al., 2010). La capacité à maintenir les deux espèces dans une association et leur productivité est affectée par les différences morphologiques, essentiellement de taille et de physiologie des partenaires puisque ces différences conditionnent la capture des éléments nutritionnels présents dans l’environnement et indispensables à la croissance des plantes dans ces peuplements hétérogènes.
Au-delà du choix des espèces à associer, le choix des variétés et mêmes des génotypes au sein d’une variété s’avère crucial. En condition de culture associée, d'importantes différences de performance existent entre diverses variétés d'une même espèce (Zannone et al., 1986; Guy, 1989). Les variétés répondent à leur environnement de culture et leur comportement en peuplement plurispécifique n’est en général pas le même qu'en culture monospécifique. Néanmoins, la sélection des génotypes et des variétés se fait en conditions de culture pure ce qui peut être limitant pour identifier les variétés adaptées aux mélanges d’espèces. De plus, les caractéristiques des cultures qui se révèlent être utiles en culture monospécifique peuvent être moins pertinentes en association; il y a donc un besoin de savoir quels sont les caractères qui procurent les informations pertinentes pour les mélanges. Afin de réussir une association d’espèces, une meilleure connaissance des interactions entre les deux espèces associées est requise (Baudoin et al., 1997). Les caractères utiles dans une association sont ceux qui traduisent les relations de complémentarité et de compétition entre les espèces associées (Davis and Woolley, 1993). Par conséquent, le degré d’interaction entre les espèces est un élément essentiel et l’effet d’un caractère particulier d’une espèce sur l’autre espèce est à considérer (Lithourgidis et al., 2011). Parmi les caractères utiles dans une association, ceux qui sont liés à l’interception du rayonnement sont majeurs, puisqu’ils conditionnent la productivité, la pérennité et la qualité du fourrage et qu’ils conditionnent indirectement la fixation et l’assimilation de l’azote. Généralement dans les associations, deux ressources sont particulièrement disputées : la lumière dont l’acquisition dépend de la morphogenèse aérienne et l’azote qui est absorbé par le système racinaire. Les caractères liés à la dynamique de l’azote sont également importants.
Dans ce contexte, cette thèse s’inscrit dans un programme de recherche sur les associations légumineuses-graminées qui a été mis en place à l’INRA de Lusignan visant de mieux comprendre ces interactions entre plantes. Son objectif général est d’étudier la variabilité génétique de la luzerne cultivée en association avec une graminée fourragère. La démarche développée comprend deux étapes. La première étape consiste à identifier et hiérarchiser les caractères d’adaptation à la compétition de la luzerne en association avec la fétuque élevée (Festuca arundinacea), et la caractérisation de leur diversité génétique à l’échelle inter et intraspécifique. La deuxième étape traite de l’analyse du contrôle génétique de ces caractères. Afin de répondre à ces deux objectifs, deux dispositifs expérimentaux ont été mis en place. Le premier traitant du premier objectif comprend un ensemble de génotypes issus de variétés et populations de luzerne et deux génotypes de fétuque élevée. Le deuxième dispositif comprend
17
une population F1 issue d’un croisement de deux parents apparentés faisant l’objet d’une population de cartographie et de détection de QTL. L’étude des interactions entre génotypes d’espèces différentes devient rapidement complexe et volumineuse au niveau expérimental. Le niveau d’étude choisi est celui qui permettra d’analyser des mécanismes d’interactions entre plantes, en complément d’évaluations agronomiques menées en dehors de cette thèse. Pour ces raisons, nous avons choisi de réaliser, au sein des deux dispositifs, des mini-peuplements (ou îlots) dans lesquels la caractérisation de chaque plante est possible. Cela permet d’étudier une large gamme de génotypes de luzerne associé à une diversité représentative de la fétuque. Dans cette étude, on exclut l’analyse de la compétition pour l’eau et pour les minéraux autres que l’azote.
19
2
Synthèse bibliographique
Cette synthèse bibliographique comporte trois grandes parties, la première partie traitera des associations légumineuses-graminées. La deuxième partie rappellera les caractéristiques de la luzerne et quelques généralités sur la fétuque. La troisième partie présentera la méthodologie de cartographie génétique chez les autotétraploïdes et l’étude du déterminisme génétique des caractères quantitatifs.
2.1 Les associations légumineuses-graminées
2.1.1 Historique et situation actuelle
Pour des raisons de coût de production et d’impacts environnementaux, les espèces fourragères pérennes doivent fournir une part croissante de la ration alimentaire des animaux (Lemaire et al., 2005b). Les graminées et les légumineuses présentent des caractéristiques de fonctionnement différentes et complémentaires en termes de croissance, de production et de qualité. Lorsque ces deux types d’espèces sont cultivés séparément en monocultures, et dans le cas des espèces annuelles, leurs atouts sont valorisés par les rotations biennales et triennales des cultures avec la légumineuse en tête de rotation. Pour les cultures pérennes, la co-culture de graminées et de légumineuses est la seule façon de bénéficier pleinement de leurs avantages particuliers. D’ailleurs, les mélanges d’espèces dans les systèmes fourragers sont connus, depuis longtemps, pour leurs intérêts économiques et écologiques. Jusqu’aux années 1950, la culture des légumineuses était importante dans l’agriculture française et européenne. Cependant, avec l’intensification des élevages et des pratiques agricoles permise par l’utilisation des engrais azotés de synthèse et avec l’accroissement des surfaces de céréales et oléagineux, la part des légumineuses dans les rotations a baissé (Du et al., 2008). En même temps, les prix très faibles du tourteau de soja, sous-produit de la production d’huile en Amérique, n’ont pas incité à la culture de légumineuses en France ou en Europe, conduisant à une dépendance protéique préoccupante depuis plusieurs décennies. Pour les productions fourragères, à partir des années 1960, période de la « Révolution Fourragère », la prairie monospécifique de graminées (à base de ray-grass, fétuque élevée ou de dactyle) abondamment fertilisée se généralise. Récemment, les limites de ce système agricole se sont faites évidentes : forte consommation énergétique liée à l’utilisation d’engrais azotés de synthèse, pollution des eaux en nitrates, dépendance protéique avec des marchés internationaux instables, coûts de production élevés. Suite à la prise en compte de ces
préoccupations environnementales et aux changements des contextes agricoles, l’intérêt pour les cultures de légumineuses a été renouvelé visant à économiser l'azote grâce à leur capacité de fixation symbiotique de l’azote atmosphérique (Capitaine et al., 2008). Dans le cas des cultures fourragères pérennes, les légumineuses sont utilisées en mélange avec des graminées dans ce qu’on appelle des prairies multispécifiques. Cette modalité se rapproche de la composition en espèces des prairies permanentes qui sont toujours multispécifiques. La culture en association, définie comme la culture simultanée de deux espèces dont une légumineuse, sur la même parcelle et pendant une durée significative de leur cycle de croissance (Willey, 1985), correspond à un cas particulier de la prairie multispécifique.
2.1.2 Intérêts et attendus des associations
Les motivations des agriculteurs vis-à-vis des prairies multispécifiques et particulièrement des cultures en association légumineuses-graminées sont multiples. Pour un éleveur, la sécurité d’un système fourrager est assurée via un rendement total suffisant en quantité et en qualité par rapport aux besoins des animaux, tout en limitant les coûts de production. Avec des besoins des animaux variant peu au cours de l’année, une répartition régulière de la production fourragère est un atout important. Globalement, la production fourragère est importante au printemps, modérée en automne et faible en été surtout lorsque la sécheresse est intense. Cependant, les espèces ont des capacités de production saisonnière différentes, par exemple, certaines graminées démarrent tôt au printemps, la luzerne est capable de production significative en été. Les cultures d’associations légumineuses-graminées permettent de valoriser ces potentiels contrastés de production saisonnière. A ces différences s’ajoutent des critères de résistance ou de tolérance aux stress abiotiques (sécheresse, humidité excessive, basses températures…) se traduisant par des repousses contrastées des espèces face à ces aléas ce qui permet l’étalement de la pousse au long de l’année et atténue la variabilité interannuelle. Les légumineuses cultivées en association avec des graminées peuvent être une alternative à l’introduction d’azote minéral (Fustec et al., 2010). Par une réduction importante voire une absence de fertilisation azotée palliée par l’introduction de légumineuse, l’association permet une économie budgétaire, une économie d’une grande quantité d’énergie fossile ainsi que de l’azote assurant une diminution des risques de lixiviation des nitrates dans le sol et notamment la pollution de la nappe phréatique, aussi une réduction de l’émission des gaz à effet de serre (Peoples et Baldock, 2001). Ces systèmes à bas niveau d’intrants doivent néanmoins continuer à être performants en termes de production fourragère qui doit être
21
maintenue au cours du temps, c’est ainsi que la notion de pérennité de la prairie ou de l’association constitue une composante essentielle dans la multifonctionnalité de celles-ci (Huyghe et Litrico, 2008). La préoccupation des agriculteurs lorsqu’ils cultivent des associations est plus particulièrement le maintien des deux espèces dans des proportions adéquates. Des conditions favorables à l’une ou à l’autre des espèces tendent à déséquilibrer le peuplement, jusqu’à obtenir de fait une monoculture. La pérennité des deux espèces dans les associations, au fil de l’exploitation de la prairie est donc un objectif qui vise un maintenir un niveau de production de fourrage de qualité équilibré en protéines et en énergie sur plusieurs années. L’intérêt nutritionnel des cultures fourragères pour les animaux est exprimé au travers de la composition biochimique. Le fourrage produit par les cultures en associations de graminées et de légumineuses est décrit pour avoir une valeur nutritive « correcte » (Bignon, 2007) grâce à la combinaison des qualités des deux partenaires. Si les graminées fourragères produisent une biomasse de haute valeur énergétique, les légumineuses fourragères confèrent au fourrage une bonne teneur en protéines (17 à 25% en moyenne). Une telle production limite les besoins d’achats de protéines sur le marché et contribue à une autonomie protéique des exploitations. Les agriculteurs visent à améliorer leurs rendements par un choix variétal permettant de tirer la meilleure production avec un moindre coût de fertilisants. Le mélange d’espèces possède d’autres atouts. L’introduction de biodiversité dans les systèmes de culture permet une meilleure maîtrise des pathogènes et ravageurs, dont les attaques deviennent à moindre effet (Plantureux et Amiaud, 2008; Teasdale, 1996). Cette diversité a un effet bénéfique sur une lutte contre les adventices, positionnant la culture des associations comme une solution alternative suite à la réduction sévère du nombre de désherbants homologués sur les légumineuses comme par exemple la luzerne (Lebois et al., 2008). Au total, les associations réduisent le recours aux pesticides, assurant une diminution des coûts de production pour l’agriculteur (Follis, 1993). Indirectement, ce type de culture assure des fonctions de restructuration du sol par une exploration meilleure avec différents profils racinaires, de diminution de l’érosion, de la battance et du ruissellement et d’amélioration de l’activité biologique du sol (Swift et al., 2004).
En résumé, les services écosystémiques que doivent fournir des prairies d’association se traduisent par une notion de multifonctionnalité dans l’écosystème cultivé. Il s’agit de trouver un équilibre entre des fonctions productives et économiques (production en quantité et en qualité) et des fonctions non productives telles que la préservation de l’environnement (gestion des ressources naturelles – sols, eau, réduction des pesticides, valorisation de la
biodiversité) ou les préoccupations sociales (occupation des terres, protection des paysages, flexibilité d’exploitation, pérennité de l’exploitation) (Hervieu, 2002). Les associations légumineuses-graminées contribuent ainsi à élaborer un système agricole qui respecte l’environnement et s’intègre bien dans un contexte d’agriculture durable.
2.1.3 Fonctionnement de l’association légumineuses-graminées
L’objectif des cultures en association est de valoriser les différences écologiques, nutritionnelles et agronomiques des espèces associées en termes de production en quantité et en qualité et de bénéfices environnementaux. Le devenir d'une plante (production, teneur en azote, survie) dans un couvert mixte dépend de sa capacité à capter et à utiliser les ressources fournies par l'environnement : rayonnement solaire, N, eau, etc. Dans ces peuplements hétérogènes, les espèces associées sont en interaction. La production de ces espèces et leur équilibre dépendent de leurs capacités d’être complémentaires dans leur exploitation des ressources dans le milieu qu’elles partagent. Il s’agit de limiter la compétition pour l’acquisition des ressources et favoriser la facilitation entre les différentes composantes. Pour prévoir le devenir des constituants des cultures associées, le contrôler, ou créer de nouvelles associations plus productives, il est nécessaire de connaître les interactions dans différentes conditions du milieu où la performance d’un individu dépend non seulement de son génotype, mais aussi de l’identité spécifique de ses voisins.
La culture en association peut être considérée comme un cas particulier de culture en situation de compétition. La compétition est définie par une interaction entre individus d’espèces identiques ou différentes pour une ou plusieurs ressources disponibles en quantités finies étant limitantes pour la croissance. Elle correspond à une situation où les effets des espèces sur les ressources induisent une réponse négative (réduction de la survie, de la croissance ou de la reproduction) chez au moins certains individus (Begon et al., 1996). Dans un milieu donné, les individus les plus compétitifs sont qualifiés d’agressifs et leur intensité de compétition est déterminée par leurs caractéristiques morphologiques et physiologiques. Il existe deux types de compétition pour les ressources limitantes : la compétition symétrique (les plantes se partagent équitablement les ressources ou proportionnellement à leur taille) et la compétition asymétrique (un des individus possède un avantage disproportionné en terme d’acquisition des ressources par rapport aux autres individus) (Bauer et al., 2004). Une utilisation différente des ressources permettra aux espèces d’échapper à la compétition et contribuera à une meilleure efficience des espèces. La complémentarité entre espèces se traduit ainsi par leur
23
capacité à acquérir et utiliser différentes ressources ou à utiliser la même ressource à des moments et des endroits différents. C’est un mécanisme essentiel permettant de renforcer la diversité résultant de la coexistence de différents groupes fonctionnels : des espèces conservatrices (avec de faibles taux de croissance, faible surface foliaire spécifique) et des espèces d'exploitation (avec des taux de croissance rapides, une surface foliaire spécifique élevée) en particulier dans des environnements difficiles (Gross et al., 2007). Les interactions entre individus qui induisent des effets positifs existent aussi. Les plantes peuvent faciliter d'autres plantes de deux manières ; directement, par l'amélioration des caractéristiques de l’environnement qu’elles partagent (en modifiant les caractéristiques du substrat, ou en augmentant la disponibilité d'une ressource, classiquement l’azote apporté par une légumineuse) ; ou indirectement, en éliminant des concurrents potentiels (par l'introduction d'autres organismes utiles tels que les microbes du sol, mycorhizes, ou pollinisateurs, ou en offrant une protection contre les herbivores) (Callaway, 1995). La balance entre la compétition, la facilitation et la complémentarité varie en fonction de la taille, l’âge et la densité des espèces en interaction ainsi que des conditions du milieu et les deux mécanismes peuvent coïncider (Callaway et Walker, 1997).
2.1.3.1Partage de la lumière en association
L’interception du rayonnement et plus précisément de la radiation photosynthétiquement active est certainement un moteur important de la compétition entre les plantes (Abberton et Marshall, 2005). La compétition pour cette ressource est un mécanisme majeur qui contrôle la croissance des plantes et conditionne l’acquisition des autres ressources (Keddy, 2001). L’architecture de la canopée et les propriétés optiques des feuilles et du sol se comptent parmi les facteurs affectant l’interception de la lumière (Sinoquet et Caldwell, 1995). La morphogenèse aérienne, et en particulier la hauteur des plantes, la distribution relative de leurs surfaces foliaires par rapport aux plantes voisines et leur efficacité d’interception du rayonnement solaire liée en particulier à leur inclinaison influencent le partage de la lumière et de la répartition de la demande en eau (Louarn et al., 2012). Les graminées ont généralement des feuilles plus inclinées que les légumineuses et ont besoin d’un nombre plus important de feuilles pour absorber une certaine radiation (Chamblee et Colins, 1988). La compétition pour la lumière est très asymétrique (Schwinning et Weiner, 1998). La lumière est fournie de façon directionnelle et sa disponibilité diminue exponentiellement depuis le haut de la canopée. La nature asymétrique de la compétition pour cette ressource fait que
l’essentiel de la ressource est capté par la plante dominante et devient non disponible pour la plante dominée limitant fortement sa croissance (Ford et Diggle, 1981). Dans un couvert végétal dense, l’inégalité de taille même pour les plantes du même âge est considérée comme la première cause de compétition pour la lumière (Schwinning et Weiner, 1998). Pour une même hauteur et une même surface foliaire, la légumineuse avec des feuilles érigées est souvent plus efficace à intercepter le rayonnement que la graminée (Louarn et al., 2010). Dans les associations avec des légumineuses de plus grande taille que la graminée (cas luzerne – graminée), la compétition pour la lumière se fait au profit de la croissance de la légumineuse et tend à exclure la graminée de l’association (Chamblee et Colins, 1988). L’effet d’ombrage peut affecter la croissance de la graminée et sa suppression de l’association. En mélange, l’ombrage peut affecter l’une ou l’autre des deux espèces à certains stades (Faurie et al., 1996). La compétition pour la lumière commence dès l’émergence des feuilles. A ce stade, une absorption maximale de la radiation par les feuilles est indispensable et la graminée (cas de la fétuque élevée) peut montrer une certaine compétitivité vis-à-vis de la luzerne (Chamblee et Colins, 1988). Dans les environnements contraints, l’ombrage, conséquence de la compétition pour la lumière, peut réduire le volume des racines et rend par conséquent la plante moins compétitive pour absorber l'eau et les minéraux (Cruz et Sinoquet, 1994). Dans un peuplement, les individus de petite taille reçoivent de la lumière avec un rapport rouge clair/ rouge sombre plus faible et une intensité de bleu diminuée ce qui induisent une élongation foliaire plus importante (Berntson et Wayne, 2000). Cette élongation est une réponse plastique à l’ombrage causé par les individus les plus hauts (Dodd et al., 2005).
L’étude de la compétition pour la lumière à l’échelle de la plante individuelle est possible grâce aux modèles disponibles en écophysiologie pour les cultures pures (caractérisation de l’architecture) avec un certain degré de complexité pour les systèmes multispécifiques lié à des différences spatiales marquées de la densité et de l’inclinaison des feuilles et dont les travaux sont en cours (Malézieux et al., 2009).
2.1.3.2Fixation et transfert de l’azote atmosphérique dans les associations
graminées-légumineuses
En association avec les graminées, la capacité de la légumineuse à fixer le N2 atmosphérique est responsable de son plus grand intérêt agronomique surtout que les carences en azote sont souvent responsables de baisses de rendement chez les graminées. Les légumineuses sont
25
capables de réaliser une symbiose avec les organismes procaryotes des genres Rhizobium et Bradyrhizobium dits « diazotrophes » qui possèdent une enzyme appelée la nitrogénase capable de réduire le diazote de l’air (N2) (forme très stable chimiquement et présente en grande quantité) en NH4+ (forme absorbée par les plantes à partir du sol). La fixation symbiotique fait intervenir des réactions complexes entre la bactérie et la plante hôte (Ledgard et Steele, 1992). A un type de légumineuse ne correspond qu’un type de bactérie (Rhizobium melliloti pour la luzerne). La capacité de fixation dépend de la variété et même du génotype de la légumineuse ainsi que de la souche bactérienne. Cette capacité de fixation diffère également selon les espèces et entraine une dépense d’énergie plus importante de la part de la légumineuse (2.5 mg de C/mg de N fixé contre 1.6 mg de C/mg de N provenant des nitrates selon (Hartwig, 1998)). Certains facteurs peuvent réduire cette activité fixatrice tels qu’une forte disponibilité de N dans le sol (principalement les ions NH4+ et NO3- qui inhibent l’activité de la nitrogénase de la bactérie) ainsi que certaines situations environnementales limitantes tel qu’un stress hydrique (Jayasundara et al., 1998). Cette capacité dépend aussi des besoins de la plante et les plantes avec un faible taux de croissance ont des besoins faibles en N, et donc fixent une quantité réduite de N2.
Dans les couverts mixtes, la présence d'une composante légumineuse (capable de fixer le N2 atmosphérique) contribue à enrichir le sol par une entrée d’azote gratuite disponible pour la communauté végétale (Hauggaard-Nielsen et al., 2009). Ainsi, dans le cas des associations graminées-légumineuses, cet enrichissement peut être classé comme un processus intervenant dans la facilitation via une augmentation de la ressource qui bénéficie plus à la graminée compagne. Dans les associations trèfle blanc et ray-grass anglais, la majorité du rendement en azote de la graminée proviendrait de la fixation symbiotique de N2 (Zanetti et al., 1997). Le transfert de N de la légumineuse vers la graminée peut recouvrir deux formes : un transfert apparent et un transfert réel et varie fortement selon l’espèce de légumineuse, le mode de culture et le temps (Hogh-Jensen, 2006). Le transfert apparent résulte de la différence de compétitivité entre la graminée et la légumineuse pour le N du sol. Il y a une spécialisation de la graminée pour le N minéral et de la légumineuse pour le N2 atmosphérique. La graminée ne bénéficie pas ainsi directement du N2 fixé. En prélevant le N du sol, la graminée « oblige » la légumineuse à fixer le N2 atmosphérique pour satisfaire ses besoins et la légumineuse laissera ainsi une part plus importante du N du sol disponible pour la graminée que si celle-ci était cultivée en pur. On parle de complémentarité de niches pour l’azote. Le second type de transfert concerne directement le devenir du N2 fixé et son transfert réel à la graminée
associée. Le processus de transfert du flux net de la légumineuse en direction de la graminée s’appelle la rhizodéposition. Ce processus comprend deux principaux mécanismes : l’exsudation par les racines de composés solubles riches en N (acides aminés, protéines, hormones excrétées par les racines) et le turn-over racinaire ou décomposition des tissus morts de la légumineuse (racines, nodosités, feuilles…), ce qui rend le N organique disponible par minéralisation (Fustec et al., 2009). L'importance relative de ces deux mécanismes et la quantité de N mise à disponibilité dépendent des espèces de légumineuses ; l’exsudation racinaire est plus importante pour le trèfle que pour la luzerne (Louarn et al., 2010). Le mécanisme de rhizodéposition a été décrit sur des cultures pures et des associations de trèfle violet, trèfle blanc et ray-grass anglais où les rhizodépôts ont atteints 40% de N chez le trèfle et la part de N de la graminée issu de la légumineuse a atteint 50% (Hogh-Jensen, 2006). Lorsqu’il y a fixation, le transfert est principalement lié aux échanges locaux de N entre les deux partenaires avec des dynamiques de disponibilité différentes (immédiats pour les exsudats ou sur le long terme pour les résidus dont la minéralisation dépend fortement de la température et la disponibilité en eau). Pour les associations pérennes à base de luzerne, ce n’est qu’à partir de la deuxième année de culture que ces transferts agissent sur la nutrition azotée de la graminée (Tomm et al., 1994; Tomm et al., 1995). Dans le cas des pâtures, un transfert supplémentaire indirect du N transformé dans les parties aériennes des plantes consommées par les animaux, par l’intermédiaire des excréments est également possible (environ 20 % de l’azote fixé par le trèfle blanc mis en disponibilité pour le ray-grass anglais) (Ledgard, 1991).
En association, la fixation symbiotique du N2 atmosphérique par les légumineuses est linéairement liée à l'accumulation de la biomasse chez ces plantes (Høgh-Jensen et Schjoerring, 2001; Corre-Hellou et al., 2006). Nyfeler et al. (2011) ont démontré que les mélanges avec 40 à 60% de légumineuses (espèces Trifolium) maximisent l'acquisition et la transformation de N en biomasse. De plus, la croissance est déterminée par le partage de la lumière entre la graminée et la légumineuse. Cette compétition pour la lumière détermine donc indirectement la quantité de N2 fixé en déterminant le potentiel de croissance de la légumineuse. Elle influence la demande en N de chaque composante et le taux de N absorbé par les plantes. Au final, les transferts de N dépendent de la fixation de N2 atmosphérique par la légumineuse régie par les phénomènes de compétition pour la lumière et pour l’azote entre les deux partenaires (Soussana et Tallec, 2010). Tous ces échanges restent encore mal caractérisés dans leur dynamique et dans la quantification des flux. L’analyse du transfert
27
d’azote reste difficile à étudier, mais progresse grâce à l’utilisation de traçage isotopique 15N (Mahieu et al., 2007; Mahieu et al., 2009).
Les interactions entre les légumineuses et les graminées en association été largement décrites (Haynes, 1980). En mélange, les différences morphologiques des parties racinaires entre les deux espèces et même au sein d’une espèce sont marquées ; la légumineuse avec des racines pivotantes atteignant les horizons profonds du sol (cas de la luzerne) et la graminée généralement avec un système racinaire fasciculé à enracinement peu profond, ce qui n’est pas le cas de la fétuque élevée à enracinement profond. Des différences phénologiques et un asynchronisme des cycles de développement entre les deux espèces favorisent par exemple la graminée au printemps et la légumineuse comme la luzerne en été lorsque les températures sont élevées (Davies, 2001). Ces disparités auraient des conséquences fortes pour la compétition et pourraient conduire à une relative séparation des niches exploitées par les deux espèces durant leur croissance ainsi qu’une diminution de la sensibilité aux contraintes ce qui contribue à une complémentarité des ressources. Par conséquent, la nature dynamique de l’équilibre en association permet le maintien des deux espèces au cours du temps. En début du cycle, les plantes doivent établir rapidement leur système racinaire qui permet de capter les ressources hydriques et minérales et particulièrement le pivot chez les légumineuses, principal lieu de stockage des réserves qui conditionne la repousse après une coupe (Avice et al., 1997; Berg et al., 2009; Volenec et al., 1996). L’acquisition de la ressource est proportionnelle aux besoins nécessaires pour la croissance ultérieure et le N du sol est disponible pour les deux espèces associées. A cette phase d’installation, la légumineuse repose sur l’azote minéral du sol pour satisfaire ses besoins, du fait du temps nécessaire pour la formation de nodosités actives (Voisin et al., 2002). Grâce à sa croissance racinaire plus rapide, les graminées sont généralement plus compétitives, à ces stades végétatifs, pour l’acquisition de N du sol que les légumineuses (Haynes, 1980). En cours du cycle, cette compétition interspécifique n’est limitante pour la croissance que lorsque la disponibilité en N minéral est faible. Dans des situations où la légumineuse tire son N de la fixation symbiotique uniquement, la complémentarité de niche joue un rôle important pour la graminée, qui contrairement à sa culture pure où elle est en compétition avec elle-même, a accès à une plus grande quantité de N. En comparaison aux cultures pures de graminées ou de légumineuses, la quantité de N prélevée en mélange d’espèces est bien supérieure (Corre-Hellou et al., 2006).
2.1.3.3 Compétition pour d’autres éléments : eau, autre minéraux
Comme pour l’azote, les plantes interagissent entre elles pour la capture des autres minéraux du sol et pour l’eau. La morphologie racinaire dans le sol est cruciale dans la compétition pour l’eau et les autres minéraux. Le besoin en eau des plantes dans une association est étroitement lié à l’interception et au partage de lumière dans le couvert (Lemaire, 1995). L’eau commence à être un facteur limitant lorsque le stock du sol n’assure plus les besoins de l’un ou des constituants de l’association. Ce stress que peut subir chaque composante de l’association peut varier selon les capacités de prélèvement d’eau de chacune ainsi que de leur capacité à placer leurs racines dans les horizons du sol où l’eau est présente. Cette capacité est déterminée par la vitesse et la profondeur d’enracinement, la densité et la distribution du système racinaire ainsi que de la durée de vie des racines (Ozier-Lafontaine et al., 1998). En situation de stress hydrique et à même profondeur d’enracinement, les graminées grâce à leur morphologie racinaire présentent des capacités à extraire l’eau plus importantes que les légumineuses (Davies, 2001). Le stress hydrique provoque une accumulation de N minéral dans le sol qui agit négativement sur la fixation symbiotique de N2 atmosphérique. Dans ces conditions, la légumineuse repose sur le N du sol pour satisfaire ses besoins et sa fixation symbiotique diminue (Ledgard et Steele, 1992). En association, cette situation affectera la nutrition azotée de l’espèce associée par une augmentation des phénomènes de compétition pour l’azote du sol. Les légumineuses à système racinaire pivotant puissant telle que la luzerne les rends plus compétitives pour l’absorption de l’eau que la plupart des graminées. Comme pour l’azote les graminées apparaissent plus compétitives pour les autres minéraux tels que le phosphore et le potassium (Davies, 2001). Pour le phosphore, les exigences des graminées sont moindres que celles des légumineuses expliquant en partie la plus grande compétitivité des graminées en conditions limitantes (Voisin et al., 2002).
2.1.4 Caractères impliqués dans le fonctionnement d’une association
L’étude de la compétition et la facilitation entre les espèces associées mobilise plusieurs disciplines : l’agronomie, l’écologie (Malézieux et al., 2009) et l’écophysiologie. Les caractères liés à l’interception du rayonnement sont majeurs, puisqu’ils conditionnent directement la productivité, la pérennité et la qualité du fourrage et qu’ils conditionnent indirectement la fixation et l’assimilation de l’azote (Lemaire et al., 2005a). En culture pure, il apparaît clairement que la hauteur de la tige, le port, le nombre de tiges et de ramifications ainsi que la surface foliaire contribuent à la performance des individus en situation de
29
compétition (Allirand, 1998). Par ailleurs, la hauteur des plantes est considérée comme un caractère pertinent pour caractériser l’aptitude compétitrice (Real et al., 2000). La hauteur influence aussi d’autres dimensions telles que le diamètre des tiges (Hakl et al., 2012). La hauteur d’une plante doit être considérée en relation avec les hauteurs des plantes voisines puisque être plus haute que la voisine confère un avantage compétitif en terme d’acquisition de la lumière (King, 1993; Falster et Westoby, 2005). Les caractères liés à la dynamique de l’azote sont également importants, puisqu’ils conditionnent directement l’intérêt des associations. Deux types de mesures sont utilisés pour la gestion d’une culture prairiale : mesure de la teneur en N de la biomasse aérienne et mesure du N minéral du sol. La mesure du statut azoté des parties aériennes est généralement définie à partir de l’Indice de Nutrition Azoté (INN) (Farruggia et al., 2004). Cet indice se base sur la courbe de dilution de l’azote : lors de la croissance d’une culture, la teneur en azote évolue au cours du temps en lien avec la biomasse totale produite, le changement du rapport entre feuilles (riches en azote) et tissus de soutien (pauvres en azote), et la disponibilité en azote du sol (Gastal et Lemaire, 2002). Il est possible de définir une courbe critique de dilution de l’azote qui permet de connaitre, pour toute biomasse, la teneur en azote minimale correspondant à une croissance optimale. Cette courbe sert de référence au diagnostic de statut azoté (Figure 1).
Figure 1 Principe du diagnostic de nutrition azotée par analyse de plante : a) courbe d’azote
critique et calcul de l’indice azote, b) quantité d’azote prélevé correspondant à la teneur critique.
Le rapport entre la teneur en azote mesurée sur un échantillon et la teneur en azote critique pour la même biomasse définit l’INN (N observé/N critique). S’il est inférieur à 1, la croissance de la culture est limitée par l’azote. Sa validité est cependant limitée aux cultures
pures et beaucoup de précautions doivent être prises en cultures associées, particulièrement celles impliquant des légumineuses (Cruz et Lemaire, 1987). Récemment, il a été montré que l’INN était, chez les graminées, directement corrélé à la teneur en azote des feuilles bien éclairées du couvert (Figure 2) (Gastal et al., 2001). Cet indicateur fonctionne aussi bien en pur qu’en prairies multi-spécifiques et permet un suivi régulier non destructif de l’état azoté d’une plante.
Figure 2 Corrélation entre l’Indice de nutrition azotée (INN) et la concentration en azote des feuilles éclairées des graminées (Nl), INN = 0.264 Nf – 0.106 (Gastal et al., 2001).
2.1.5 Choix variétal et maintien des deux espèces
Même si la culture en association graminées-légumineuses est d’usage commun, le maintien d'une proportion appropriée de chaque espèce sur la durée de vie de la prairie (3 à 5 ans, voire plus) reste un défi. Le choix des espèces est d'une importance majeure. Le degré d’interaction entre les espèces associées est un élément essentiel à prendre en compte lorsqu’on choisit les génotypes à associer. Ce choix influence également la décision de l’introduction d’intrants ou non, et le choix de densité de chaque espèce (Haynes, 1980). Pour les mélanges binaires, l’association trèfle blanc (Trifolium repens) - ray-grass anglais (Lolium perenne) est souvent utilisée sous un climat frais et humide tempérée pour le pâturage. L'association de la luzerne (Medicago sativa) avec la fétuque élevée (Festuca arundinacea) ou dactyle (Dactylis glomerata) est plus adapté aux zones sèches sous les climats tempérés ou méditerranéens, pour la fauche. Dans la plupart des cas, les pratiques agricoles, telles que la densité de semis,
% N d e s fe u ill e s é cl a ir é e s
31
la fertilisation azotée et de la fréquence de coupe, sont utilisés pour modifier la proportion des espèces. Le choix des variétés qui sont inclus dans un mélange peut contribuer à modifier l'équilibre entre les espèces. L'effet du génotype sur l'interaction entre les légumineuses et les graminées a beaucoup été étudié sur le mélange trèfle blanc - ray-grass anglais. La capacité compétitive du trèfle blanc s'explique principalement par la taille des feuilles, les variétés à grandes feuilles ont une plus grande capacité concurrentielle que les variétés intermédiaires ou à petites feuilles (Annicchiarico, 2003; Williams et al., 2003). Pour les mélanges luzerne - graminée, sous des conditions de culture qui sont favorables à la survie de la luzerne (sol, rythme de coupe), la luzerne tend à exclure la graminée (Chamblee and Colins, 1988), car elle possède une meilleure capacité à intercepter la lumière. De plus, l’enrichissement du sol en N par la luzerne dans la première année, ce qui défavorise la graminée (Ta et Faris, 1987; Hardarson et al., 1988; Burity et al., 1989). L’effet de la variété de luzerne sur le devenir d’une association luzerne-graminée a été étudié par (Zannone et al., 1983; Zannone, 1985; Mosimann et al., 2007).
2.2Présentation de la luzerne et généralités sur la fétuque élevée
2.2.1 La luzerne
2.2.1.1Origines et expansion géographique
L'histoire de la luzerne est liée essentiellement à l’histoire de l'élevage dans le bassin Méditerranéen. Elle est reconnue et domestiquée par l’homme antique comme étant une plante précieuse. Elle est un facteur essentiel de l'évolution des civilisations modernes. Elle a été identifiée, il y a près de 9 000 ans, dans le croissant fertile, en Asie mineure sur les hauts plateaux du Caucase, en Iran et en Turquie comme un fourrage facile à cultiver et à conserver, productif et très apprécier par les animaux. Les espèces apparentées sont dispersées en Asie centrale jusqu’à la Sibérie. Introduite en Grèce en 479 av. J.-C., la luzerne est diffusée en Europe méditerranéenne et en Afrique de l’Est puis du Nord par les conquêtes d’où elle s’est répandue sur tous les continents (Mauriès, 1994).
Il est admis que la luzerne est la plante la plus ancienne cultivée uniquement pour son fourrage. Lors des fouilles archéologiques en Turquie, on a découvert des tablettes hittites où il est fait mention que les animaux s’alimentaient de luzerne. Des preuves historiques témoignent d’une propagation à grande échelle de la luzerne à partir de la région nord-ouest de la Perse (ancien Iran) à la Ie millénaire av. J.-C. et en plaine de Mésopotamie (Irak);
carrefour entre l’Asie, l’Afrique et l’Europe. Le commerce marin, étant développé à l’Est de la Méditerranée, a contribué à cette propagation. Au IVe siècle av. J.-C., la luzerne était introduite en Grèce avec les invasions des armées des Mèdes (peuple de l’Iran ancien), ces derniers l’ont implantée afin de nourrir leurs animaux. Pour les 200 années suivantes, on n’a jamais de références sur la luzerne. C’est avec l’Empire romain, qu’on a entendu parler de nouveau de la luzerne (Genier et al., 1992). Durant le Ie siècle av. J.-C., les romains l’ont cultivé en Italie et en Andalousie en Espagne. A la même époque, elle a été cultivée dans les oasis du nord d’Afrique, le sud de la France, en Suisse. Son expansion s’est poursuite vers l’est, en 126 av. J.-C., depuis le Turkestan russe vers la Chine en introduisant des races de chevaux par le général Zhang Quian. Il y a aussi eu une introduction de la luzerne en Espagne lors des invasions arabes par le Nord de l’Afrique (Hendry, 1923). La chute de l’Empire romain a marqué l’interruption de la culture la luzerne en Europe qui est restée probablement à l’état spontané (Michaud et al., 1988). A partir du XVIe siècle, selon Klinkowski cité par (Michaud et al., 1988) la luzerne s’est propagée de l’Espagne en France en 1550, en Hollande en 1565, en Angleterre en 1650, en Allemagne en 1750, au Suède en 1770 et en Russie lors du XVIIIe siècle. A partir de cette époque, la luzerne a été distribuée à travers le monde entier à partir de l’Europe vers les nouvelles terres comme l’Amérique, l’Australie et la Nouvelle-Zélande.
La luzerne est une des plantes fourragères les plus répandues dans le monde. Son plus grand développement se trouve dans les zones tempérées chaudes : Etats-Unis, Europe, Amérique du Sud, Asie, Japon, Australie, Nouvelle-Zélande, Afrique du Nord et Argentine (Barnes et al., 1990; Mauriès, 1994).
2.2.1.2Evolution et taxonomie
Comme décrit auparavant, la luzerne serait originaire de Médie (région allant du nord-ouest de l'Iran jusqu’à l’Azerbaïdjan) et l’appellation « medica » fait référence aux Mèdes d’où le nom scientifique pour le genre de la luzerne Medicago. Quant à l’appellation « alfalfa » utilisé en espagnol et en américain, elle proviendrait du nom arabe « al-fi fi a » ou « al-fac-façah » signifiant père des aliments qui auraient été muté en « alfalfa » en Espagne médiévale.
Sinskaya (1950) était l’un des premiers taxonomistes qui a intégré les caractères des légumineuses et la couleur des fleurs dans la classification des complexes de Medicago sativa (Gunn et al., 1978) mais l’analyse la plus complète du genre Medicago est l’étude
33
taxonomique de Lesins et Lesins (1976) (Quiros et Bauchan, 1988). D’ailleurs, une unification de la nomenclature a été réalisée par (Heyn, 1963), (Lesins et Lesins, 1979) puis par (Small et Brookes, 1984; Small et Brookes, 1990) qui ont réduit le nombre de taxons et les ont ramenés au rang de sous-espèces. Ces sous-espèces sont d’ailleurs confirmées par les phylogénies réalisées à l’échelle du genre Medicago (Bena et al., 1998; Bena, 2001).
Le genre Medicago appartient à l’ordre des Fabales, la superfamille des Légumineuses, la famille des Fabaceae, la sous-famille des Faboïdeae (Doyle et Luckow, 2003) et la tribu des Trifolieae (Choi et al., 2004b). La tribu Trifolieae comprend trois genres Medicago, Trigonella et Melilotus et regroupe les plupart des légumineuses fourragères comme la luzerne cultivée et les trèfles (Figure 3). Les trois genres sont très proches.
Figure 3 Dendrogramme représentant les relations phylogénétiques entre espèces de
légumineuses (Zhu et al., 2005).
En se basant sur les caractères morphologiques décrivant la forme de fleurs, la couleur des inflorescences, la forme des gousses, la forme des gousses et des graines, le genre Medicago a été classé en quatre sous-genres : Lupularia, Medicago, Orbicularia et Spirocarpos. Ces quatre sous-genres comprennent un total de plus de 60 espèces différentes dont les 2/3 sont des annuelles et 1/3 des pérennes (Quiros et Bauchan, 1988). Le sous-genre Medicago est composé d’espèces pérennes et comprend quatre sections : Arboreae, Marinae Grossh.,