Modulation de l'activité transcriptionnelle du récepteur aux estrogènes [bêta] par le facteur de transcription SCL/Tal-1
106
0
0
Texte intégral
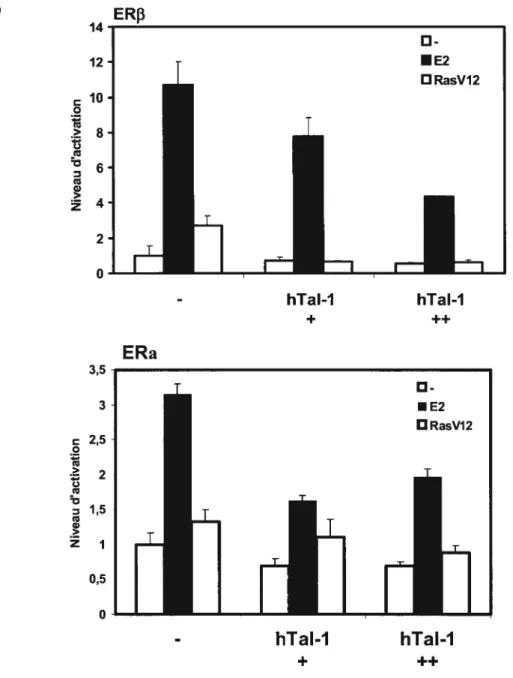
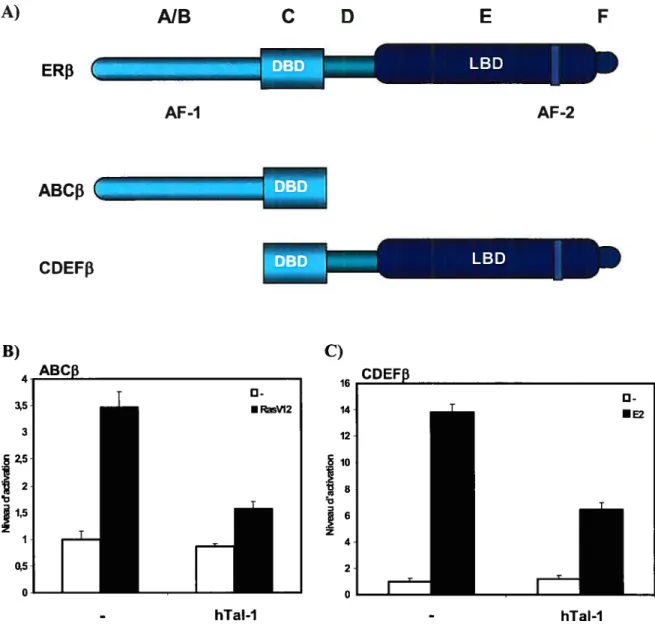
Figure
+7
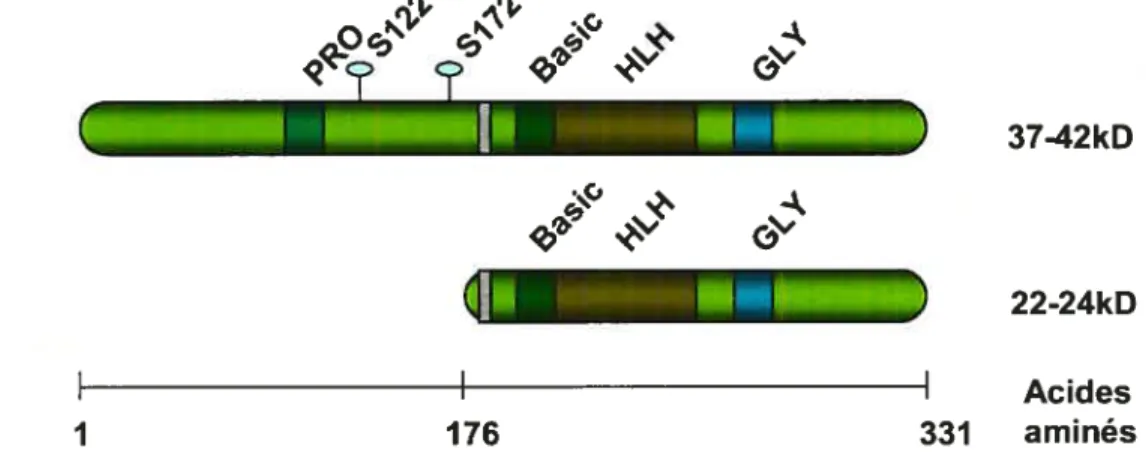
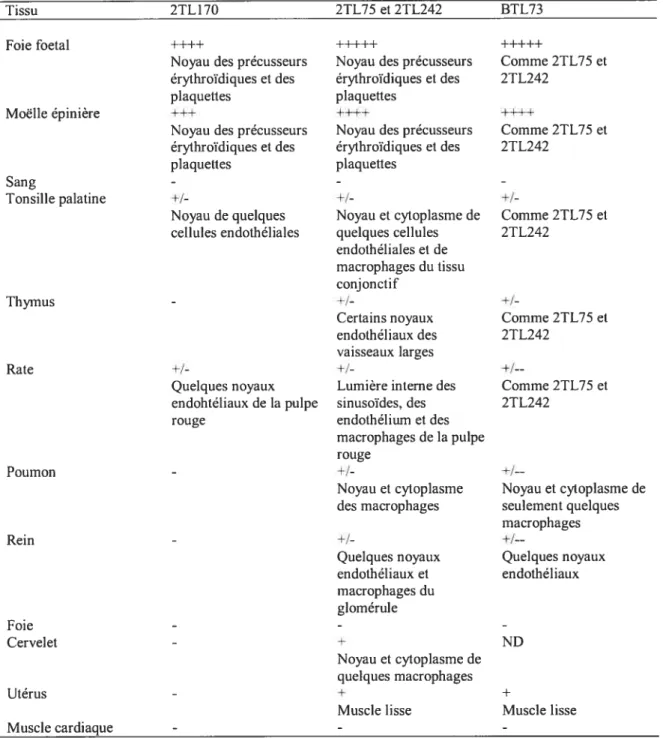
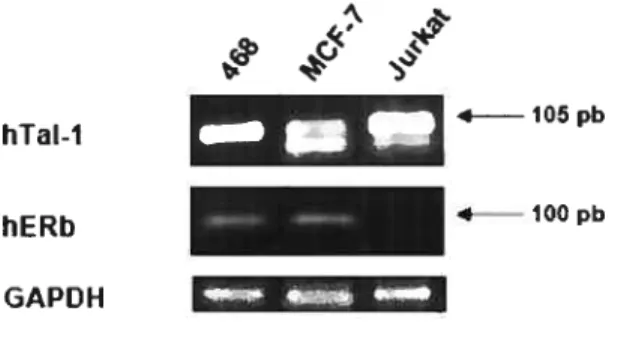
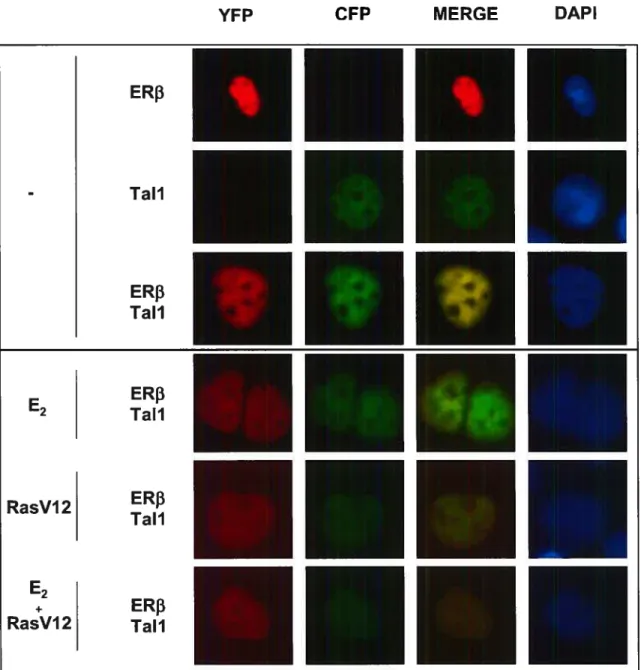
Documents relatifs