Direction des bibliothèques
AVIS
Ce document a été numérisé par la Division de la gestion des documents et des archives de l’Université de Montréal.
L’auteur a autorisé l’Université de Montréal à reproduire et diffuser, en totalité ou en partie, par quelque moyen que ce soit et sur quelque support que ce soit, et exclusivement à des fins non lucratives d’enseignement et de recherche, des copies de ce mémoire ou de cette thèse.
L’auteur et les coauteurs le cas échéant conservent la propriété du droit d’auteur et des droits moraux qui protègent ce document. Ni la thèse ou le mémoire, ni des extraits substantiels de ce document, ne doivent être imprimés ou autrement reproduits sans l’autorisation de l’auteur.
Afin de se conformer à la Loi canadienne sur la protection des renseignements personnels, quelques formulaires secondaires, coordonnées ou signatures intégrées au texte ont pu être enlevés de ce document. Bien que cela ait pu affecter la pagination, il n’y a aucun contenu manquant.
NOTICE
This document was digitized by the Records Management & Archives Division of Université de Montréal.
The author of this thesis or dissertation has granted a nonexclusive license allowing Université de Montréal to reproduce and publish the document, in part or in whole, and in any format, solely for noncommercial educational and research purposes.
The author and co-authors if applicable retain copyright ownership and moral rights in this document. Neither the whole thesis or dissertation, nor substantial extracts from it, may be printed or otherwise reproduced without the author’s permission.
In compliance with the Canadian Privacy Act some supporting forms, contact information or signatures may have been removed from the document. While this may affect the document page count, it does not represent any loss of content from the document.
Les origines parallèles du phénotype bleu chez le doré jaune (Sander vitreus)
Par Martin Laporte
Département de sciences biologiques Faculté des Arts et des Sciences
Mémoire présenté à la Faculté des études supérieures En vue de l'obtention du grade de M.Sc.
En sciences biologiques
Mai 2009-05-22
Université de Montréal Faculté des études supérieures
Ce mémoire intitulé:
ii
Les origines parallèles du phénotype bleu chez le doré jaune (Sander vitreus)
Présenté par: Martin Laporte
A été évalué par un jury composé des personnes suivantes:
François-Joseph Lapointe président -rapporteur Bernard Angers directeur de recherche Pierre Magnan co-directeur de recherche Louis Bernatchez membre du jury
Résumé
Le
statut taxonomique des dorés au phénotype bleu reste toujours ambiguë. Elle varie entre le statut de polymorphisme ou encore celui de sous-espèce. Des études antérieures ont révélé une absence de variabilité au niveau des marqueurs mitochondriaux entre les dorés de phénotype bleu. et ceux de phénotype jaune. De plus, il a été impossible de réassigner les individus provenant de différents lacs selon leur phénotype, ce qui suggère une apparition parallèle de la coloration bleue. Une analyse génomique a été effectuée sur des dorés de phénotype bleu et de phénotype jaune sur plusieurs lacs du Bouclier Laurentien afin de tester cette hypothèse. Les résultats démontrent que tous les dorés (peu importe la couleur de leur phénotype) d'un même lac sont tous plus près génétiquement les uns des autres par rapport aux dorés provenant des autres lacs. Cependant, les dorés au phénotype bleu et ceux au phénotype jaune représentent des lignées évolutives distinctes à l'intérieur de chacun des lacs. Ces résultats confirment que ces lacs ont été colonisés par une seule population de dorés jaunes et que les populations de dorés au phénotype bleu ont une origine polyphylétique. Par conséquent, la position taxonomique du phénotype bleu chez le doré est clarifiée et la variabilité morphologique observée entre les différentes populations de dorés de couleur bleue est partielle~ent expliquée. L'évolution parallèle du phénotype bleu représente un excellent modèle d'étude des mécanismes génétiques sous-jacents; la sélection divergente et l'isolement reproducteur.IV
Summary
The situation of the blue phenotype in the walleye remains ambiguous, varying between a polymorphism or a subspecies. Previous studies revealed no mitochondrial variability and failed to assign individuals from different lakes to a given phenotype, suggesting a parallel appearance of the blue phenotype. A genomic analysis was performed on blue and yellow phenotypes from severa! Laurentian Shield lakes to assess this hypothesis. Results indicate that, whatever their phenotype, individua!s from a given lake were more genetically similar than those from other lakes. However, blue and yellow phenotypes represent distinct evolutionary lineages within each Jake. These results confirm the co!onization of each lake by walleye and the polyphyletic origin of the blue phenotype. This clarifies the taxonomic position of the walleye complex and partially explains the morphological variability among blue phenotype populations. The parallel evo!ution of the blue phenotype represents an excellent mode! to study the mechanisms underlying divergent selection and reproductive isolation.
Table des matières
Résumé-- --- iii
Summary--- iv
Table des matières --- v
Listes des tableaux--- vii
Liste des viii Liste des s ix Remerciements--- xii 1. Introduction--- 1 1.1 Contexte scienti 2 1.1.1 biologique--- 3 1.1.2 locale--- 7 1.1.3 Flux génique--- 7 1.1.4 Mutation--- 8 1.1.5 Dérive 9 1.1.6 Niche Il 1.1.7 Types de sélection naturelle--- Il 1.1.7.1 Sélection directionnelle--- 12
1.1.7.2 Sélection stabilisante---~--- 13
1.1.7.3 Infériorité des hétérozygotes--- 14
1.1.8 Types de spéciation--- 15 1.1.8.1 La spéciation allopatrique--- 15 1.1.8.2 La spéciation péripatrique--- 16 1.1.8.3 La spéciation 17 1.1.9 Convergence évolutive--- 17 1.1.10 Évolution 18 1.1.10.1 Observée chez les populations 19 1.1.10.2 Sympatrique et issue d'invasion multiples--- 20
1.1.10.3 Sympatrique et ,issue de colonisation 21 1.2 Sujet de l'étude--- 22
1.2.1 Hi des dorés au phénotype bleu--- 22
1.2.2 Conservation des dorés au phénotype bleu--- 23
1.2.3 Hypothèse sur l'origine du phénotype bleu--- 23
1.2.4 Les origines du phénotype bleu--- 26
1.2.4.1 Observation prédisant un isolement reproducteur- 26 1.2.4.2 Observation prédisant une évolution 26 1.2.4.3 Observation prédisant une colonisation 27 1.2.5 Objectif de l'étude--- 27
2. Article: parallel origins of the blue phenotype in walleye---- 29
Accord des coauteurs--- 30
Abstract--- 31
Introduction--- 32
Materials and Methods--- 35
Results--- 38
Discussion--- 42
45 3. Discussion--- 47
3.1 Position taxonomique des dorés au phénotype b1eu--- 48
3.2 Niche écologique des dorés au phénotype bleu--- 49
3.3 Couleur bleue: cause ou conséquence de l'isolement reproducteur
?---
533.4 3.5 3.6
VI
Futur évolutif des populations au phénotype bleu: Espèce polyphylétique ou adaptation radiative ?---Gestion des populations au phénotype bleu---Le phénotype bleu : Indicateur de biodiversité
?---55 57 58 4. Références--- 60
Liste des tableaux
Table 2.1 Distribution of sampled individuals from the six study lakes.--- 36
Table 2.2 Distribution of the number of loci obtained from the eight combinations of selective primers used with the AFLP method. The number of variable loci is in parentheses.--- 37
Table 2.3 Results of the reassignment test performed on blue and yellow phenotypes. The analysis was performed on 10 groups (yellow form from lakes A, C, D, E, and F; blue from lakes A, B, C, and D; saugers from lake E; refer to Table 1 for lake identifications). Only the results of blue phenotype are presented. Sample size is in parentheses.--- 41
viii
Liste des figures
Figure 1.1 Hypothèse concernant les différentes origines possibles du phénotype bleu. Les dorés au phénotype bleu sont représentés par les lignes bleues, ceux au phénotype jaune par des lignes jaunes et les points représentent un ancêtre commun.--- 25
Figure 2.1 Location of populations sampled for the present study (Québec, Canada). --- 36
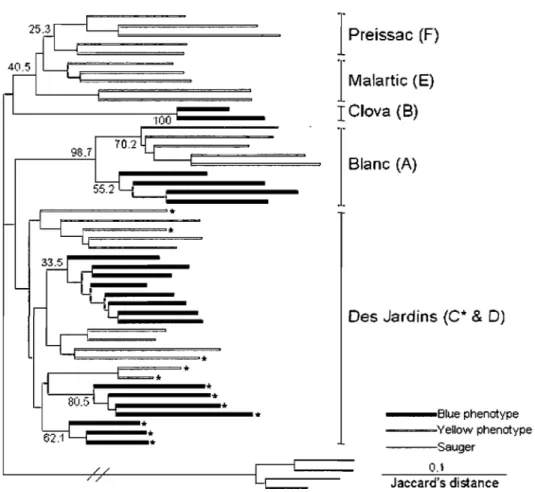
Figure 2.2 Relationships among the 51 individuals (26 yellow phenotypes, 21 blue phenotypes, and 4 sauger) as inferred from NJ computed on the Jaccard's distance. Bootstraps values are indicated. Asterisks represent the individuals caught in the western part of Lake Des Jardins.--- 40
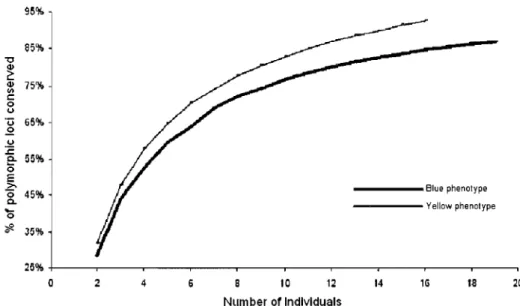
Figure 2.3 Percentage of polymorphie loci as a function of the number of individuals per phenotype. The analysis was performed on the 35 walleye captured in lakes where both blue and yellow phenotypes were sampled.--- 42
Liste des abréviations
A a ADN AFLP COSEPAC COSEWIC DNA dNTP ESU FQRNT GRILHO
Hl H2 H3 L Km mM MRNF mtDNA NE ng NJ nM NSERC PCR ppm Allèle numéro 1 Allèle numéro 2 Acide désoxyribonucléiqueAmplified fragment length polymorphism
Comité sur la situation des espèces en péril au Canada Committee on the Status of Endangered Wildlife in Canada Deoxyribonucleic acid
Deoxynucleotide triphosphates Evolutionary significant Unit
Fonds Québécois de la recherche sur la Nature et les Techonologies
Groupe de recherche interuniversitaire en limnologie et en environnement aquatique Hypothèse nulle Hypothèse 1 Hypothèse 2 Hypothèse 3 Litre Kilomètre Millimolaire
Ministère des Ressources Naturelles et de la Faune du Québec Mitochondrial deoxyribonucleic acid
Taille efficace de la population Nanogramme
Neighbor joining Nanomolaire
Natural Sciences and Engineering Research Council of Canada Polymerase chain reaction
x
pmol Picomolaire
Qc Québec
SSCP Single strand conformation polymorphism
Taq The,rmus aquaticus
TBE Tris-Borate-EDTA
U Unité
~l Microlitre
UV Rayon ultra violet IOX Dix fois concentré 0.5X 0,5 fois concentré
% Pourcentage
« Le
bleu est une plongée inconsciente et interminable»
XII
Remerciements
Je tiens tout d'abord à remercier mon directeur de recherche, Bernard Angers, pour tous ses conseils et son expertise qu'il a mis à ma disposition. C'est avec beaucoup d'ouverture d'esprit qu'il m'a laissé choisir mon sujet de maîtrise, me permettant ainsi d'évoluer à ma façon au sein d'une ambiance intellectuellement stimulante. Je n'oublierai jamais le souci de la précision des mots qu'il m'a inculqué, ni l'importance de la rigueur qu'un scientifique se doit de maintenir autant face à ses pairs que face au grand public. J'aimerais aussi le remercier pour les expériences qu'il m'a fait vivre et qui m'ont permis peu à peu de me débarrasser du syndrome de l'imposteur, trop souvent observé chez les nouveaux étudiants en recherche.
Je remercie aussi mon codirecteur de recherche, Pierre Magnan, pour nos rencontres et nos discussions qui m'ont permis d'avancer dans le cadre de ce projet. Je tiens aussi à le remercier pour sa confiance à mon endroit qui nous amène maintenant vers de nouveaux horizons.
Je ne pourrais passer sous silence les collègues de mon laboratoire qui m'ont soutenu, tous à leur façon, lors des épreuves qu'un projet de maîtrise peut contenir. Merci à vous tous (Frédéric Cyr, Émilie Castonguay, Daniel Cossios, Claude-Olivier Sylva-Beaudry, Philippe Girard, Rachel Massicotte, Joëlle Boizard et Mathieu Chouteau).
Ce projet n'aurait jamais vu le jour sans l'aide de collaborateurs qui ont partagé leur donnés ou qui ont permis la capture des dorés. Merci à Yves Paradis, Jean-Pierre Hamel, Louis Houde, Daniel Nadeau et Jocelyn Mercier du Ministère des Ressources Naturelles et de la Faune (MRNF) du Québec; Stéphanie Pellerin; Guillaume Bourque; Richard Monfette du magazine Sentier Chasse-Pêche; Jacques Mercier de la Station de Biologie des Laurentides de l'Université de Montréal; Magella et Linda Larochelle de la Pourvoirie Rothmans; et Marc Lucotte de l'Université du Québec à Montréal.
Ce projet à été financé grâce à la participation des Fonds Québécois de la Recherche sur la Nature et les Technologies (FQRNT), du conseil de recherches en sciences naturelles et en génie (CRSNG) du Canada et de la Faculté des Études Supérieures de l'Université de Montréal (bourse de la Banque Nationale).
Je terminerai cette liste en remerciant ma famille et mes amis qui me sont très chers. Leur support inconditionnel m'a grandement aidé et je leur en serai toujours reconnaissant.
1.1 CONTEXTE SCIENTIFIQUE
L'évolution biologique se défini par les changements de fréquences alléliques et de combinaisons de loci retrouvés chez les différentes populations. Un allèle est une variante d'un locus qui correspond à une section précise du . génome. La fréquence allélique fluctue selon l'influence de quatre forces évolutives fondamentales: les mutations, la dérive génique, le flux génique et la sélection naturelle. Les processus neutres incluent les trois premières forces qui pennettent l'apparition de nouveaux allèles (mutations) et la variation des différentes variantes alléliques (dérive et flux génique). Ces processus sont fonction du hasard, pennettant de présumer que l'histoire du vivant serait différente si on la recommençait.
L'ensemble du génome est affecté par les processus neutres. Cependant, seuls les loci associés à la transcription (les gènes) et ceux qui y sont physiquement reliés peuvent être affectés par la sélection naturelle. Cette dernière pennet l'adaptation en supprimant les allèles qui désavantagent les individus face aux conditions biotiques et abiotiques de l'environnement. Cette sélection d'allèles s'effectue à partir du phénotype, l'interface entre le génotype et son environnement.
L'apparition d'un nouveau phénotype peut conduire à de nouvelles interactions entre le génotype et son environnement. Ultimement, la spécialisation
3
que permet un nouveau phénotype envers une nouvelle ressource peut mener à la spéciation (i.e.: formation de nouvelles espèces). L'étude de l'origine des phénotypes est donc une importante source de modèles biologiques permettant la compréhension des processus de spéciation.
Dans ce mémoire, nous décrirons les origines multiples d'un phénotype à l'aide d'une comparaison du génotype de différents individus. Avant de décrire le sujet de cette étude, les différents concepts et mécanismes évolutifs pouvant être reliés à l'apparition de ce phénotype seront expliqués.
1.1.1 ESPÈCE BIOLOGIQUE
Selon le concept biologique, une espèce biologique est un ensemble de loci en évolution, définitivement isolé des autres ensembles de loci. Cette définition est fortement inspirée de celle de Mayr (1970). Le terme «gène» a cependant été modifié pour «loci» puisqu'une espèce est beaucoup plus que l'ensemble de son ADN codant. Un gène est un type de loci, mais plus important encore, toute partie de l'ADN est un locus. Puisque les loci neutres sont aussi isolés que les gènes lors des processus de spéciation, ils seront propres à l'espèce.
Auparavant, cet ADN non codant était perçu comme de l'ADN peu intéressant au niveau évolutif. Il est de plus en plus reconnu que le paysage génomique d'une espèce influence son évolution. Par exemple, i) les régions
promotrices agissent comme des activateurs ou des répresseurs à la transcription, influençant la production des différentes protéines (Carninci et al. 2006), ii) les
introns permettent l'épissage alternatif chez les eucaryotes, permettant la production d'un grand nombre de protéines à partir d'un nombre beaucoup plus petit de gènes (Duan et al. 2009) et iii) les éléments transposables se multiplient de façon indépendante dans le génome, permettant aussi de multiplier certains exons à travers le paysage génomique (Esnault et al. 2000). L'évolution de cet
ADN non codant a donc une influence majeure sur l'évolution des différentes espèces.
Afin de mettre en évidence la pertinence de ce concept biologique de l'espèce, l'existence des espèces biologiques doit avant tout être démontrée. Imaginons un monde du vivant sans espèce. Ce monde formerait un seul groupe où tous les organismes pourraient se reproduire entre eux. Les génotypes les mieux adaptés à certains milieux apparaîtraient et disparaîtraient de façon stochastique sans qu'aucun mécanisme ne les concentre à l'intérieur de groupes distincts et spécialisés à leur environnement. La simple observation des différents mécanismes d'isolement reproducteur (pré et post·copulatoire) p~rmet d'affirmer qu'un tel monde biologique n'est pas représentatif de la réalité. L'espèce biologique n'est donc pas une création de l'homme. Le concept nominaliste, défini par Occam, statuant que l'espèce est une abstraction créée par l'homme ne s'applique donc pas à l'espèce biologique.
5
L'importance évolutive que sous-entend le concept biologique de l'espèce doit aussi être expliquée. Les véhicules transportant un ensemble d'allèles sont les individus. Il n'existe pas dans la nature deux organismes sexués parfaitement identiques (même les jumeaux monozygotes comportent des mutations somatiques). Par conséquent, la seule façon de décrire collectivement un groupe d'organismes est l'utilisation des différentes méthodes statistiques. Les différentes statistiques sont des abstractions mentales calculées à partir d'entités bien réelles. Par exemple, la moyenne de la longueur des ailes d'une espèce d'oiseau est une abstraction mentale calculée à partir des différentes tailles d'ailes qui sont bien réelles. Conséquemment, les variations observables au sein d'une espèce ne sont pas des illusions créées par un phénomène universel, mais bel et bien des entités réelles, soumises aux forces évolutives. Ce raisonnement est antagoniste à la pensée typologique issue de la philosophie grecque qui considère que l'essence d'une espèce (Le. un phénomène universel pouvant être décrit par un groupe de descripteurs statiques) est bien réelle, mais que les variations qui la composent sont illusoires. Le concept typologique de l'espèce est donc opposé à toute pensée évolutive et ne considère pas le caractère unique de chacun des organismes (Mayr, 1970). Ce concept ne s'applique donc pas à ce qui est observé chez les espèces biologiques.
Bien qu'il considère l'espèce et les variations des organIsmes qui la composent comme des entités réelles, le concept biologique de l'espèce a ses limites. La plus importante d'entre elles est le fait qu'il soit restreint aux
organismes sexués (Vrba, 1985). Selon ce concept, chaque organisme asexué (ne se reproduisant pas avec le reste du vivant) devrait être considéré comme une espèce. Afin de remédier à ce problème, certains ont proposé de regrouper les individus en fonction de leur degré de similarité génétique et phénotypique. Cette solution est problématique, puisqu'il n'existe aucun moyen objectif de déterminer le degré de divergence nécessaire pour nommer «espèce », un groupe
d'organismes. En réponse à ce problème, Templeton (1989) a proposé d'utiliser les mécanismes de cohésion (i.e.: les mécanismes empêchant l'isolement reproducteur tel que l'absence de différenciation morphologique et l'utilisation de ressources similaires) et Ridley (1993) d'utiliser la position écologique afin de définir les espèces. Cependant, ces efforts intellectuels notables n'ont malgré tout jamais réussi à considérer toute la complexité biologique observée chez les différentes populations. Par conséquent, il n'existe toujours pas de définition de l'espèce qui est universelle à l'ensemble du vivant.
Puisqu'aucune définition incluant tous les phénomènes biologiques ne fait l'unanimité et parce que l'organisme modèle s'y porte bien (un vertébré sexué), le concept biologique de l'espèce sera utilisé à l'intérieur de ce document. Un vocabulaire commun à l'ensemble de la communauté scientifique sera donc utilisé, ce qui permettra une meilleure compréhension des autres concepts et mécanismes expliqués plus loin dans ce travail.
7
1.1.2 POPULATION LOCALE
Les espèces sont organisées en populations, à l'intérieur desquelles les individus ont une chance plus élevée de se reproduire entre eux qu'avec les individus de n'importe quelle autre P9pulation (Hartl et Clark 1997). Ces populations constituent des sous unités d'ensembles d'allèles, évoluant de façon plus ou moins indépendante, selon l'étanchéité de l'isolement reproducteur qui les sépare. L'isolement géographique des populations est le mécanisme d'isolement le plus important en spéciation (Mayr, 1970). Cependant, deux populations vivant au même endroit peuvent évoluer de façon indépendante à l'aide d'un isolement reproducteur qui n'a pas été initié par un isolement géographique. Les mécanismes d'isolement reproducteur observés entre les populations peuvent s'éteindre et permettre un retour à l'homogénéisation des deux ensembles d'allèles. Ultimement, un isolement reproducteur entre deux ou plusieurs populations mène à la spéciation. Les populations sont donc les unités évolutives permettant d'étudier les différents stades de la spéciation.
1.1.3 FLUX GÉNIQUE
Le flux génique est la proportion d'allèles se transmettant au sein d'une population, mais provenant d'autres populations. Les allèles migrants peuvent être nouveaux ou déjà présents au sein de la population. S'ils sont nouveaux, ils auront un effet similaire aux mutations (voir section 1.1.4). C'est-à-dire qu'ils
apparaîtront au hasard au sein de la population. Si l'allèle existe déjà, son arrivée modifiera les fréquences alléliques du locus et le flux génique produira un effet stochastique similaire à la dérive génique (voir section 1.1.5).
Contrairement aux autres forces évolutives (excepté le cas de la sélection stabilisante, voir section 1.1.7.2), le flux génique ralentit la divergence des populations (Hartl et Clark, 1997). Le transfert d'allèles homogénéise donc les populations et la spéciation en cours est ralentit ou neutralisée. Une absence de flux génique témoigne de la présence d'un isolement reproducteur entre les deux populations, indiquant ainsi que les populations divergent entre elles.
1.1.4 MUTATION
Les mutations sont les changements de la séquence nucléique qui apparaissent au hasard, suite aux erreurs de copie du matériel génétique lors de la division cellulaire (les mutations peuvent aussi être issues de contact avec des sources mutagènes telles que les UV). Elles sont les seules véritables productrices de variabilité allélique. Chez les animaux pluricellulaires, seules les mutations qui surviennent à l'intérieur des lignées germinales sont héritables. L'apparition de mutations héritables permet la divergence des populations en créant de nouveaux allèles.
9
La plupart des mutations n'ont aucun effet direct sur le phénotype des organismes. Par exemple, dû à la dégénérescence du code génétique, les mutations survenant au niveau d'un gène codant peuvent modifier ou non la séquence d'acides aminés de la protéine, selon leur position au niveau d'un codon. Les mutations survenant dans des régions non codantes (par exemple, dans les sites promoteurs ou d'épissage) peuvent aussi influencer le phénotype
1.1.5 DÉRIVE GÉNIQUE
La dérive génique se défini par la fluctuation des fréquences alléliques, causée par le hasard de la reproduction. Ces fluctuations sont causées par les erreurs d'échantillonnage produites entre chaque génération. Puisque la taille des populations n'est pas infinie (en terme de nombre d'individus la composant), la probabilité qu'un allèle neutre se retrouve à la génération suivante est proportionnelle à sa fréquence dans la population. Un allèle rare a donc moins de chance de se transmettre qu'un allèle abondant. Les fluctuations causées par le hasard mènent à la disparition d'allèles et ultimement à la fixation du locus par un seul d'entre eux. La dérive génique est donc une force aléatoire qui diminue irrémédiablement la diversité allélique à l'intérieur d'une population.
L'intensité de la dérive génique dépend de la taille efficace (NE) de la population. La taille efficace est un paramètre théorique défini comme la taille d'une population idéale conduisant à la dérive observée. Ce paramètre est
influencé par le sexe ratio, le nombre de reproducteurs, la taille historique et la variance du succès reproducteur. La probabilité de fixation d'un nouvel a11èle d'un locus neutre est de
I12NE.
Selon cette équation, les populations avec de plus petites tailles efficaces perdent plus rapidement leur diversité allélique.En diminuant la diversité allélique à l'intérieur d'une population, la dérive génique augmente la diversité allélique entre les populations. Les fluctuations alléliques qu'elle cause sont issues du hasard, affectant de façon indépendante chacune des populations. Imaginons différentes populations complètement isolées les unes des autres et possédant toutes un locus neutre composé d'un même nombre d'allèles aux fréquences alléliques équivalentes. Les chances que toutes ces populations fixent le même allèle sont de IInombre allèlesnombre de populations. Par exemple, 3 populations composées de 3 allèles ont moins de 4% de chance de fixer le même allèle pour un locus donné. Puisqu'il existe plusieurs populations par espèce, plusieurs loci par génome et plusieurs allèles par locus, les chances de retrouver deux populations identiques sont pratiquement nulles. L'ensemble de la biodiversité serait donc principalement expliqué par cette force évolutive (Kimura, 1983; Lynch 2006). Elle déterminerait préalablement la variation génétique sur laquelle la sélection naturelle aurait par la suite un effet
Il
1.1.6 NICHE ÉCOLOGIQUE
Hutchinson (1957) décrit la niche écologique comme une enveloppe où chaque dimension de l'espace est une ressource (alimentaire, abris ... ) de l'environnement. Ces ressources sont limitées dans l'espace et dans le temps. Elles sont des conditions qui délimitent la position occupée par une espèce dans son environnement. Deux groupes évolutifs observés en sympatrie ne peuvent théoriquement pas occuper la même niche écologique. La compétition pour les mêmes ressources obligerait les moins adaptés à opter pour une solution alternative (telle que l'adaptation à une nouvelle ressource) ou bien à disparaître du territoire occupé par l'espèce la plus adaptée. Si la dérive génique est une force majeure expliquant la biodiversité, la sélection naturelle est la force évolutive permettant la spécialisation des populations à une niche écologique.
/./.7 TYPES DE SÉLECTION NATURELLE
La sélection naturelle modifie l'abondance des caractères héritables qui offrent des avantages différents en fonction de l'environnement. Les caractères les moins avantageux à l'intérieur d'une population pour un environnement donné auront plus de chance d'être réduits en abondance, ce qui augmentera la fréquence des caractères les plus avantageux à la génération suivante. Bien que l'unité de sélection soit le génotype de l'organisme, la cible de la sélection est son phénotype (Mayr, 1970). Le phénotype est l'interface qui permet à l'organisme
d'avoir un contact avec son environnement. C'est donc le phénotype qui est contraint à la sélection et les effets de cette force se répercutent ensuite sur le génotype. Dans certains cas précis (caractère monolocus), la cible et l'unité de sélection sont directement reliées. Un tel cas de figure sera utilisé afin de simplifier les effets de la sélection sur le génotype. Un locus à deux allèles (A et a) d'une population composée d'organismes diploïdes nous servira de modèle. Les différents génotypes possibles (AA, Aa et aa) seront comparés en fonction de leur valeur adaptative (i.e.: le nombre d'organismes générés et devenus sexuellement fertiles). Plus la valeur adaptative d'un caractère est grande, moins il y aura de chance qu'il disparaisse par sélection au sein d'une population.
1.1.7.1 Sélection directionnelle
Ce type de sélection favorise un caractère au détriment des autres. Un' des homozygotes sera donc favorisé par rapport à l'hétérozygote et à l'autre homozygote (i.e. : aa :S Aa < AA). Un cas typique de sélection directionnelle est la résistance des insectes face aux insecticides (Rathor et Wood, 1985). Les organismes possédant le ou les traits adaptés à ces conditions inhospitalières seront les seuls à se reproduire, multipliant ainsi la présence de ce ou ces traits de résistance aux insecticides à la génération suivante. Une population adaptée à cet environnement hostile sera donc obtenue à partir de quelques mutants déjà résistants à l'insecticide. La sélection directionnelle peut être antagoniste chez des
13
populations habitant des environnements différents, favorisant ainsi la divergence et ultimement la spéciation entre ces populations.
1.1.7.2 Sélection stabilisante
La sélection stabilisante élimine les formes homozygotes d'un locus. L'hétérozygote est par conséquent favorisé (i.e. : aa < Aa > AA). Un cas reconnu est l'anémie falciforme (drépanocytose) chez l'humain. L'anémie falciforme est la mutation d'un gène de la globine qui cause une déformation des globules rouges (Neel, 1949). Un individu portant deux allèles mutés produira seulement des globules rouges déformés, ce qui causera sa mort prématurée. La moitié des globules rouges d'un hétérozygote seront cependant normaux. Le porteur survivra, mais subira des anémies chroniques et des crises périodiques de douleur. Un homozygote portant deux allèles sauvages (i.e.: non mutés) ne sera pas atteint par aucun des problèmes reliés à la forme de ses globules rouges. Sous ces seules pressions, il y a une forte sélection directionnelle en faveur des homozygotes sauvages. Cependant, dans les zones de paludisme (maladie mortelle provoquée par l'infection des globules rouges par un plasmodium), les hétérozygotes sont avantagés. Les globules rouges déformés ralentiraient la circulation sanguine, ce qui protégerait face aux affections neurologiques causées par le paludisme.
La sélection stabilisante est une force homogénéisante car elle contribue à
les processus d'hybridation menant à la fusion des groupes lors d'un contact entre les populations en voie de spéciation. Lorsque deux populations spécialisées à des environnements contrastant colonisent simultanément un environnement intermédiaire, l'hybride a de bonnes chances d'être mieux adapté, ce qui favorisera la fusion des deux groupes parentaux.
1.1.7.3 Infériorité des hétérozygotes
Pour un locus donné, ce type de sélection favorise l'homogénéisation des allèles à l'intérieur des différents individus. Elle est donc caractérisée par la faiblesse des hétérozygotes (i.e.: aa > Aa < AA). Les populations subissant une telle sélection tendront vers l'une et/ou l'autre des formes homozygotes en fonction de leur valeur adaptative respective et de la dérive génique.
Un mécanisme similaire peut aussI se produire sur plusieurs loci afin d'obtenir une combinaison de caractères permettant d'augmenter la valeur adaptative de l'individu. Les individus possédant un phénotype intermédiaire seront désavantagés par rapport à l'une ou l'autre de ces stratégies évolutives. La sélection divergente peut donc permettre la divergence des populations habitant le même type d'environnement. Encore plus intéressant, elle peut diviser une population en deux groupes distincts si un isolement reproducteur (par homogamie par exemple) se produit en sympatrie entre les deux caractères différents.
15
1.1.8 TYPES DE SPÉCIATION
La spéciation est la formation de deux ou plusieurs nouvelles espèces à partir d'une seule. Ces espèces sont issues de populations qui ont divergé les unes par rapport aux autres. Le processus de spéciation est complété lorsque l'isolement reproducteur entre les populations est définitif. Différents types de spéciation ont été définis en fonction des circonstances initiales dans lesquelles s'est initiée la divergence. Les deux premiers types de spéciation sont issus d'un isolement géographique (Mayr, 1970), tandis que le troisième peut être causé par un isolement écologique ou encore par l'établissement instantané d'un isolement reproducteur.
1.1.8.1 La spéciation allopatrique
La spéciation allopatrique se produit lorsqu'une population est séparée géographiquement en deux populations de taille équivalente par une barrière. Chacune des nouvelles populations débute sa propre histoire évolutive. Ces histoires évolutives divergeront tant et aussi longtemps que la barrière les maintiendra géographiquement isolées. Les deux populations accumuleront des différences génétiques et phénotypiques avec le temps. Ces différences sont issues de processus neutres et de processus adaptatifs. Arrivées à un certain stade, les différences deviendront si importantes que la barrière n'aura plus besoin d'exister pour maintenir l'isolement reproducteur, marquant ainsi, la fin de la
spéciation. La vallée du Rift qui serait responsable de la séparation des hominidés du reste des primates est un des nombreux exemples de spéciation allopatrique. La dérive des continents a aussi permis des barrières géographiques menant à de multiples spéciations.
1.1.8.2 La spéciation péripatrique
Ce type de spéciation est semblable à la spéciation allopatrique, puisqu'une barrière géographique sépare la population en deux. Cependant, la taille (en nombre d'individus) d'une des deux populations est largement plus petite que l'autre. Ce détail, en apparence anodin, a des conséquences majeures sur l'évolution des populations. Rappelons que tous les loci sont affectés par les processus neutres et que les fluctuations causées par le hasard affectent davantage les petites populations. Tous les loci auront donc un taux de fixation plus élevé chez les petites populations. Bien que les allèles avantageux aient plus de chance d'être supprimés par hasard à l'intérieur d'une petite population, certains d'entre eux risquent aussi d'être conservés et se fixeront beaucoup plus rapidement. L'importance de la sélection dépendra de la taille de la population. Plus une population est petite, plus la dérive génique est intense et moins la sélection naturelle aura un effet substantiel sur la divergence des populations. Les plus petites populations se distinguent donc davantage par des variations de fréquences alléliques dues au hasard que par leur perte d'allèles due à une adaptation. Cette acquisition accélérée de différences génétiques et phénotypiques permet aux deux
17
populations péripatriques d'atteindre plus rapidement un isolement reproducteur définitif que ce qui serait observé lors d'une spéciation allopatrique. Plusieurs cas de spéciation péripatrique sont observés dans les archipels (Mayr, 1970).
1.1.8.3 La spéciation sympatrique
Ce type de spéciation se produit lorsqu'une population se divise en deux groupes évolutivement distincts, sans préalablement avoir recours à un isolement géographique. Par exemple, la production d'un tétraploïde auto fécond ou de tétraploïdes féconds peut produire un isolement reproducteur instantané. Il est aussi possible que la spéciation sympatrique ne soit pas causée par une modification chromosomique. Une telle situation a forcément recours à la sélection naturelle afin de permettre l'établissement de l'isolement reproducteur entre les deux nouvelles populations. La proximité entre l'habitat et le site de reproduction et une préférence pour les partenaires sexuels les plus ressemblants sont les deux tendances majeures observées lors de ces cas de spéciation sympatrique (Coy ne et Orr, 2004).
1.1.9 CONVERGENCE ÉVOLUTNE
La convergence en évolution est la présence, chez deux espèces différentes, de caractères similaires issus d'une adaptation au même type d'environnement, mais n'étant pas hérités d'un ancêtre commun. Ces caractères
similaires proviendraient d'une même réponse évolutive, obtenue de façon indépendante, par deux espèces plus ou moins soumises au même problème adaptatif. Le loup de Tasmanie (Thylacinus cynocephalus) était un marsupial très ressemblant au loup commun (Canis lupus), un euthérien. Les ailes des oiseaux et des chauves-souris ont été acquises de façon indépendante. Le colibri à gorge rubis (Archilochus colubris) et les papillons de la famille des sphingidés ont développé un bec long et un vol stationnaire adaptés pour puiser le nectar de certaines fleurs. Ces espèces distantes au ni veau phylogénétique se sont spécialisées indépendamment à des niches écologiques similaires, ou du moins à
une même ressource disponible. Ces observations démontrent qu'il est possible que des unités évolutives distinctes développent des adaptations similaires malgré un bagage génétique différent.
1.1.10 ÉVOLUTION PARALLÈLE
L'évolution parallèle est une forme de convergence qui s'est produite à
plusieurs reprises, à l'intérieur d'un même type d'écosystème et à partir d'une même structure (homoplasie) chez des organismes de lignée évolutive relativement proches l'une de l'autre (Futuyma, 1986). L'évolution parallèle est liée à des mécanismes adaptatifs puisque la répétition de changements évolutifs corrélés à l'environnement n'est généralement pas causée par le hasard (Endler, 1986). On peut retrouver des modèles d'évolution parallèle chez des populations vivant en allopatrie et en sympatrie. Les modèles de populations en sympatrie
19
peuvent par contre être issus de mécanismes différents. L'évolution parallèle engendre une répétition du même phénomène de divergence de populations permettant ainsi d'étudier les mécanismes adaptatifs à la base de la spéciation. La colonisation des lacs d'origine glaciaire en Amérique du Nord serait une cause importante d'évolution parallèle chez les :poissons d'eau douce (Skùlason et Smith, 1995).
1.1.10.1 Observée chez des populations allopatriques
Ce type d'évolution parallèle se produit lorsque toutes les populations situées dans un environnement alternatif développent un même phénotype alternatif. L'apparition de ce phénotype est associée à la colonisation d'une nouvelle niche écologique en allopatrie. Le cas des gammares de cavernes
(Gamma rus minus) illustre bien cette situation. Les populations vivant dans les rivières souterraines ont développé de longues antennes, une absence de pigmentation et une perte de vision, contrairement au phénotype commun présent dans les rivières extérieures (Kane et al., 1992). Des populations de gammares
n'étant pas connectées au même réseau hydrographique démontrent le même patron de divergence phénotypique corrélé à
r
environnement des populations. La barrière séparant les deux populations peut être d'ordre géographique ou écologique. Même si la migration peut être possible entre les deux populations, le flux génique doit par contre rester faible. Le faible taux de survie des migrants à un environnement hostile et/ou l'infériorité des hybrides permettrait de diminuerl'importance du flux génique et favoriserait la divergence entre les deux populations qui ne seraient pas isolés géographiquement.
1.1.10.2 Sympatrique et issue d'invasions multiples
Ce second type d'évolution parallèle se produit lorsque deux populations isolées l'une de l'autre entrent de nouveau en contact. Ce contact produit un phénotype alternatif chez au moins une des deux populations, permettant ainsi de coloniser une nouvelle niche écologique et d'éviter la compétition intraspécifique. Cet évènement doit se reproduire indépendamment et à différents endroits pour qu'il y ait évolution parallèle. Il a été démontré que les populations sympatriques du grand corégone (Coregonus clupeaformis) provenaient de deux refuges
glaciaires différents (Bernatchez et Dodson, 1990) et que celles de l'épinoche à trois épines (Gasterosteus aculeatus) seraient issues de deux incursions
temporelles différentes, depuis l'océan vers le continent (McPhail, 1984). Ces populations
ont
possiblement acquis une certainè forme de mécanisme d'isolement avant d'entrer de nouveau en contact. L'isolement reproducteur chez les animaux peut être issu d'une différence de sites ou de temps de reproduction. L'apparition d'un phénotype alternatif est souvent reliée à la deuxième population arrivée sur les lieux. L'avantage compétitif que peut procurer un nombre d'individus plus élevé chez la population déjà en place expliquerait cette observation (Hubbell, 2001). Le phénotype alternatif associé à la colonisation d'une nouvelle niche écologique serait donc le résultat de mécanismes adaptatifs21
permettant le maintien de deux populations génétiquement distinctes en sympatrie.
1.1.10.3 Sympatrique et issue d'une colonisation unique
Ce type d'évolution parallèle n'est pas relié à un isolement reproducteur préalablement formé en allopatrie. La divergence des populations doit se produire sur place, à différents endroits, de façon indépendante et suite à l'apparition d'un phénotype alternatif qui sera corrélé à une nouvelle niche écologique disponible. Contrairement au cas précédent, les mécanismes adaptatifs n'empêchent pas la fusion entre deux populations déjà partiellement différenciées, mais permettent l'apparition de deux populations à partir d'une seule. Le corégone européen
(Coregonus lavare/us) démontre un tel patron de différenciation à l'intérieur des
lacs Norvégiens (0stbye et al., 2006). Un modèle similaire pourrait être
observable à l'intérieur du Bouclier Laurentien où certaines populations de doré jaune (Sander vitreus) présentent un phénotype alternatif (bleu) selon un patron
1.2 SUJET DE L'ÉTUDE
1.2.1 HISTORIQUE DES DORÉS AU PHÉNOTYPE BLEU
Découverts dans les lacs Érié et Ontario, les dorés au phénotype bleu sont considérés comme une sous-espèce du doré jaune (Scott et Crossman, 1973; Trautman, 1981). Cependant, Hubbs (1926) considérait les dorés de couleur bleue de ces lacs comme une espèce à part entière (doré bleu - S. vitreus glaucum). Cette divergence d'opinion, basée sur des observations morphologiques et comportementales, n'a jamais été résolue puisque ces populations se sont effondrées vers 1959. La surexploitation par la pêche commerciale (Campbell, 1987), l'introduction d'éperlans arc-en-ciel (Osmerus mordax) (Regier et al., 1969) et l'eutrophisation du lac Érié (Campbell, 1987) sont les trois principales causes de sa disparition. Les derniers spécimens ont été capturés en 1965 (Anonyme, 1983) et on le considère disparu depuis 1985 (Campbell, 1987; COSEPAC, 2003). Cependant, la présence de dorés au phénotype bleu a de nouveau été signalée dans les lacs du Bouclier Laurentien (Paradis, 2004; Paradis et Magnan, 2005). Suite à cette information, le débat concernant le statut taxonomique du phénotype bleu a été réanimé.
23
1.2.2 CONSERVATION DES DORÉS AU PHÉNOTYPE BLEU
La curiosité que suscite un poisson d'une couleur peu commune dans les eaux tempérées du Québec a contribué à la popularité des dorés de couleur bleue auprès des pêcheurs sportifs. De plus, avant même de connaître les informations fondamentales à la gestion de cette « nouvelle » ressource, les pourvoyeurs sont de plus en plus nombreux à utiliser ces dorés de couleur bleue comme un outil de promotion. Puisque l'augmentation drastique de la pêche sportive des années 1980 a contribué au déclin de plusieurs populations de dorés jaunes (Ellison et Franzin, 1992), il devient alors critique de déterminer la position évolutive des dorés possédant le phénotype bleu à l'intérieur des lacs du Bouclier Laurentien. Cette information est nécessaire à l'établissement d'une stratégie de gestion appropriée afin de conserver une forme alternative d'un des poissons d'eau douce avec l'une des plus grandes importances économiques en Amérique du Nord (Billington et al., 1992).
1.2.3 HYPOTHÈSES SUR L'ORIGINE DU PHÉNOTYPE BLEU
L'origine d'un phénotype peut être la conséquence de différents phénomènes. Nous pouvons expliquer l'apparition du phénotype bleu par quatre hypothèses différentes (voir figure 1).
80-Telle que la couleur bleue des yeux chez l'humain, la coloration bleue serait un polymorphisme et n'impliquerait aucune structure populationnelle. Un doré au phénotype bleu pourrait donc être aussi près génétiquement d'un doré au phénotype jaune que d'un autre doré au phénotype bleu.
81- Les dorés au phénotype bleu proviendraient d'un isolement reproducteur survenu avant la colonisation post-glaciaire des lacs du Bouclier Laurentien. Toutes les populations de phénotype bleu seraient donc plus près génétiquement les unes des autres de celles au phénotype jaune. Dans ce cas, on parlerait de sous espèces ou d'espèces différentes.
82- Les dorés au phénotype bleu proviendraient d'une évolution parallèle issue de contacts indépendants entre deux populations provenant de refuges glaciaires différents (i.e. : évolution parallèle reliée à une invasion multiple). Une population de phénotypes bleus et une population de phénotypes jaunes habitant un même lac ne seraient donc pas reliées par un ancêtre commun immédiat. Ces populations seraient plutôt reliées aux autres populations provenant du même refuge glaciaire. L'apparition du phénotype bleu ne serait pas corrélée aux différents refuges glaciaires, mais plutôt à l'ordre d'arrivée des populations à l'intérieur des lacs.
83- Les dorés au phénotype bleu proviendraient d'une évolution parallèle issue de populations où la divergence génétique s'est entièrement produite sur
25
place, suite à la colonisation post-glaciaire (i.e.: évolution parallèle reliée à une colonisation unique). Une population de phénotypes bleus proviendrait de la population de phénotypes jaunes habitant le même lac. Il y aurait donc présence d'un ancêtre commun immédiat pour tous les dorés (hormis les dorés noirs) d'un même lac.
HO
H1
Lac 1 Lac 2 Lac 1 Lac 2H2
H3
Lac 1 Refuge 1 Lac 2 Lac 2 Refuge 2 Lac 1Figure 1.1 : Hypothèses concernant les différentes origines possibles du phénotype bleu. Les dorés au phénotype bleu sont représentés par les lignes bleues, ceux au phénotype jaune par des lignes jaunes et les points représentent un ancêtre commun
Lac 1 Lac 2 Lac 1 Lac 2 Lac 1 Lac 2
/.2.4 LES ORIGINES DU PHÉNOTYPE BLEU
1.2.4.1 Observation prédisant un isolement reproducteur
Il est peu fréquent d'observer deux groupes différenciés par plusieurs caractères morphologiques au sein d'une seule population panmictique (exceptés les caractères différenciant les mâles et les femelles). Puisque les dorés au phénotype bleu se distinguent par leur sécrétion de sandercyanine à la surface de leur épiderme (Yu et al., 2008), par leur croissance plus lente et aussi par leur
espace interobital plus étroit (Hubbs et LagIer, 1964; Paradis et Magnan, 2005), l'hypothèse du polymorphisme est moins vraisemblable. De plus, il a été démontré que les dorés au phénotype bleu préfèrent une eau plus profonde et plus froide que ceux au phénotype jaune (Parsons, 1967). Il semblerait que cette préférence soit conservée lors des périodes de reproduction (Stone, 1948). Une telle observation permet d'envisager que les dorés au phénotype bleu constitueraient probablement un groupe évolutif distinct de ceux au phénotype jaune.
1.2.4.2 Observation prédisant une évolution parallèle
La dispersion de deux groupes évolutivement distincts devrait se traduire par deux distributions géographiques différentes. La présence du phénotype bleu est cependant toujours associée à celle du phénotype jaune (communications avec
27
la MRNF-Abitibi). Il est donc peu probable qu'une séparation survenue avant la colonisation post-glaciaire soit à l'origine de l'apparition du phénotype bleu. Nous serions plutôt devant un cas d'évolution parallèle, où l'apparition de la coloration bleue s'est produite à différents endroits après la colonisation postglaciaire.
1.2.4.3 Observation prédisant une colonisation unique
Afin qu'il puisse y avoir une évolution parallèle avec une invasion multiple, il doit y avoir deux populations préalablement isolées géographiquement qui entrent de nouveau en contact. Une étude génétique indique que les dorés jaunes (l'espèce) du Bouclier Laurentien proviendraient d'un seul refuge glaciaire (Billington et al., 1992). Puisque la colonisation s'est produite suite au retrait des glaces, il y a environ 12000 ans et qu'aucune raison ne permet de croire à une seconde colonisation produite ultérieurement, l'évolution parallèle issue d'une invasion multiple de refuges différents ne semble pas être à l'origine du phénotype bleu.
1.2.5 OBJECTIF DE L'ÉTUDE
L'objectif de cette étude est de déterminer si les populations au phénotype bleu sont apparues à partir d'une évolution parallèle issue d'une colonisation unique. La comparaison de génotypes d'individus au phénotype jaune et
d'individus au phénotype bleu provenant de 6 lacs du Bouclier Laurentien servira à démontrer l'objectif en trois étapes. Il sera démontré (i) qu'il y a un isolement reproducteur entre les dorés au phénotype bleu et ceux au phénotype jaune d'un même lac, (ii) que les populations au phénotype bleu ont émergé suite à la colonisation post-glaciaire et (iii) que les lacs échantillonnés ont été colonisés une seule fois par les dorés jaunes (l'espèce).
29
ABSTRACT
The situation of the blue phenotype in the walleye remains ambiguous, varying between a polymorphism or a subspecies. Previous studies revealed no mitochondrial variability and faited to assign individuals from different lakes to a given phenotype, suggesting a parallel appearance of the blue phenotype. A genomic analysis was performed on blue and yellow phenotypes from several Laurentian Shield lakes to assess this hypothesis. Results indicate that, whatever their phenotype, individuals from a given Iake were more genetically similar than - those from other lakes. However, blue and ~ellow phenotypes represent distinct evolutionary lineages within each lake. These results confirm the colonization of each lake by walleye and the polyphyletic origin of the blue phenotype. This clarifies the taxonomie position of the walleye complex and partially explains the morphologicai variability among blue phenotype populations. The parallel evolution of the blue phenotype in represents an excellent model to study the mechanisms underlying divergent selection and reproductive isolation.
KEYWORDS
32
INTRODUCTION
Phenotypic change in c10sely related lineages frequently follows a common pathway in response to similar environmental pressures (Endler, 1986), resulting in the evolution of similar but independently derived ecotypes (Le.: distinct populations adapted to an alternative environment). This is a common feature in fishes from north temperate lakes (Taylor, 1999; Skùlason and Smith, 1995). This process, termed parallel evolution (Futuyma, 1986), pro vides a system of replicates that can be statistically assessed to study the adaptive mechanisms involved in speciation (Schluter, 2000).
According to the "trigger" of this process, parallel evolution can be separated into two different models: the "multiple invasions" and the "unique colonization" models. The multiple invasions model occurs wh en two or more genetically distinct populations come into contact with each other at multiple locations. The phenotype of one or both populations may then change similarly but independently in each location. Such a process, which has been reported in numerous species complex (Bernatchez and Dodson, 1990; McPhail, 1984), provides invaluable information about the mechanism underlying reinforcement between sympatric populations (Coyne and Orr, 2004).
In the unique colonization model, the appearance of the alternative phenotype occurred when a part of the ancestral population diverged from a
distinct population. This process may occur at different locations and provide similar phenotypes if an environmental configuration lead to reproductive isolation between the phenotypes (e.g., 0stbye et al., 2006). Because divergence
occurred strictly in sympatry, such a scenario is an ideal opportunity for studying mechanisms underlying divergent selection and reproductive isolation.
The bJue phenotype of the walleye, Sander vitreus (formerly Stizostedion vitreum), seems to be a typical case of parallel evolution. First described in Lake
Erie and Lake Ontario (Hubbs, 1926), it was extensively exploited until its extinction in 1965 (COSEWIC, 2003). It is distinguishable from the corn mon walleye by its col our, size, growth rate, morphology, and behavioural characteristics such as a preference for deeper and cooler water (Stone, 1948; Trautman, 1981; Campbell, 1987; Paradis and Magnan, 2005).
Different studies based on behavioural and morphological traits have investigated the taxonomie position of the blue phenotype. Hubbs (1926) first referred to the blue phenotype from the Great Lakes (commonly named blue walleyes or blue pike) as a distinct species (Sanqer glaucus), but their position
was retrograded to subspecies (Sander vitre us glaucus) after individuals
considered intermediate between the two groups were observed (Scott and Crossman, 1973). Stone (1948) concluded that the blue and yellow phenotypes from lakes Erie and Ontario were distinct species with different subspecies in each of these lakes. Recently, a genetic study based on one nuclear and two
34
mtDNA genes could not cluster ail the blue phenotypes of the Great Lakes into a single evolutionary unit (Morrison and King, unpublished data). They suggested that the blue phenotype may represent a morphotype (i.e.: distinct population with a different form), distinguishable at the population level. Moreover, the blue phenotypes outside the Great Lakes were not assigned subspecies status because they lack "typical" blue walleye traits (Hubbs and LagIer, 1964; Campbell, 1987), although no direct evidence supports this conclusion (Scott and Crossman, 1973). Populations with blue phenotypes have recently been described in Laurentian Shield lakes (Paradis, 2004). Sorne "typical" blue walleye traits, such as different head morphology and slower growth, were observed in the blue individuals of these populations (Paradis and Magnan, 2005).
Several pieces of évidence suggest sympatric parallel evolution from a unique colonization event in the blue walleye. i) Many morphological and behavioural characteristics are correlated with the blue phenotype (Stone, 1948; Paradis and Magnan, 2005), although there is large variability among populations. For instance, reclassification based on morphological characters allowed more efficient discrimination of both phenotypes for a given lake than when several populations of Laurentian Shield lakes were considered (Yves Paradis, Ministère des Ressources Naturelles et de la Faune [MRNF] du Québec, personal communication). ii) The geographic distribution of the blue phenotype is discontinuous and included into the one of the yellow phenotype. Furthermore, the blue phenotype is only found in sympatry with the yellow phenotype
(Jean-Pierre Hamel, Louis Houde, Daniel Nadeau and Jocelyn Mercier, MRNF, personal communications). Together, these observations may reflect the local and independent origin of the blue phenotype. iii) Based on mitochondrial DNA, the postglacial dispersal of walleye in the Laurentian Shield may originate from a single refuge (Billington et al., 1992). The presence of a single lineage allows one
to rule out the hypothesis of contact between divergent populations.
The objective of this study was to determine. the origines) of the blue phenotype occurring in Laurentian Shield lakes using a genomic approach. To test our prediction of sympatric parallel evolution of the blue phenotype from a unique colonization event, our specifie hypotheses were: (i) within a given lake, blue and yellow phenotypes represent distinct lineages, (ii) the blue phenotype appeared independently from the yellow phenotype across lakes and (iii) walleye originate from the same founder group.
MATERIALS AND METHODS
Sampling
A total of 51 walleye from five lakes located in the Laurentian Shield (Figure 2.1) were sampled; 26 and 21 specimens were unambiguously assigned to the yellow or the blue phenotype, respectively, according to the colour of their epidermis. The remaining 4 individuals belong to saugers (Sander canadenses)
36
and served as an outgroup. For a given lake, the number of individuals varied from two to 14 (Table 2.1) .
....
NORTHo
Lake • City 250 KM CANADA 5km DesJardins ®(C&D)Figure 2.1: Location of populations sampled for the present study (Québec, Canada).
Table 2.1: Distribution of sampled individuals from the six study lakes. Walleye
Sauger Total Geographie coordinates
Code Lake na me l:ellow blue
A Blanc 5 4 0 9 47°44'50"N-73° 12'01"W B Clova 0* 2 0 2 48°07'07"N-75°31' 19"W C Des Jardins W. 6 8 0 14 46°37' 37"N-78° 19'27"W D Des Jardins E. 5 7 0 12 46°38'45"N-78° 15'43"W E Malartie 5 0 4 9 48° 15'26"N-78°07'51
"w
F Preissac 5 0 0 5 48°22'II"N-78°19'38"W Total 26 21 4 51* Walleyes with yellow phenotype were also present in this lake.
Genetic analysis
DNA was extracted with standard phenol-chloroform procedures (Sambrook et al., 1989). Genetic variability at the scale of the genome was
surveyed using the AFLP procedure according to Vos et al. (1995). DNA
fingerprints were performed with eight selective combinations (Table 2.2). The presence of loci was assessed with a denaturing polyacrylamide gel stained with
silver nitrate (Bassam et al., 1991). An mtDNA analysis on the control region
was done using the CR-3-F (CATAATTNKATMTCAAGTGCATAA) and T-phe-R (GTGTTATGCTTTAGTTAAGC) primers. PCR reactions were conducted from a template of 12.5 !lI (1.25 !lI of IOX Taq buffer (Feldan-Bio Inc.,
St-'Laurent, Québec); 2.5 nM . L-I of each dNTP; 10 pmol of each primer; 0.2 Unit of Taq polymerase and 100 ng of DNA) with 45 cycles of 3 steps (15 sec. at
92°C, 15 sec. at 48°C, and 15 sec. at 68°C). The segment variability was screened using the SSCP method (Sheffield et al., 1993). PCR products were separated on
6% non-denaturing acrylamide gels (37:5: 1 ratio acrylamide-bisacrylamide) for 12 hours at 20 watts at 4°C in 0.5 X TBE.
Table 2.2: Distribution of ~he number of loci obtained from the eight combinations of selective primers used with the AFLP method. The number of variable loci is in parentheses.
MseI EcoRI Total Sauger Walle~e Yellow Blue
CAA AGC 51 34 43 43 43 CAC ACC 22 18 18 18 17 CAC AGC 49 37 45 45 44 CAC AGG 31 23 25 25 24 CTG ACC 26 17 22 22 21 CTG ACG 27 16 26 26 26 CTG AGC 40 28 33 33 31 CTG AGG 29 24 25 24 25 Total 275 (209) 197 (30) 237 (149) 236 (145) 231 (117) Statistical analysis
To determine whether the phenotypic groups are genetically distinct, phylogenetic relationships among individuals were inferred. A Jaccard distance matrix (J accard, 1900) was computed from the presence-absence of loci for the 51 individuals. Relationships were inferred with the neighbor-joining (NJ) method (Saitou and Nei, 1987) using the POPULATIONS 1.2.30 software
38
(Langella, 1999). Bootstraps with 1000 replicates on loci were perforrned with the PHYLIP 3.6 software (Felsenstein, 1989).
Reassignment tests were perforrned to validate the result of the phylogenetic tree. The groups were defined according to the lake, the two phenotypes and the monophyletic groups in the phylogenetic tree. The AFLPOP 1.1 software (Duchesne and Bernatchez, 2002) was used to compute the maximum likelihood of each individual for each group. The allele frequency was estimated assuming Hardy-Weinberg equilibrium. Because AFLP produce dominant markers, the "zero" and the "1" frequency values were set to a correction value of 0.01 and 0.99900001 respectively.
To compare the genetic diversity between phenotypes, the proportion of polymorphie loci as a function of the number of individuals was estimated. This allows one to take into account the different sample sizes of the two phenotypes. For a given number of individuals and for each phenotype, the number of polymorphie loci was averaged across 1000 permutations on individuals of the same phenotype. The R 2.4.1 software was used to compute the permutation.
RESULTS
A total of 275 loci were screened among the 51 individuals (Table 2.2). Of these loci, 237 were present in walleye, with the rest being specifie to the sauger. Walleye have 24 pairs of chromosomes (Gold et al., 1980), providing a coverage
l.
of approximately 10 loci per chromosome; 149 (63%) of the 237 loci are variable in the walleye (Table 2.2).
Similar numbers of loci were detected in the yellow (236) and the blue (231) phenotypes. Within yellow and blue phenotypes, 61 % of the 236 loci and 51 % of the 231 loci, respectively, are variable (Table 2.2). No diagnostic allele (present in aIl individuals of a given group) between the yeIlow and blue phenotypes was detected over alliakes. A single private allele (present in a single group) was detected for the yellow phenotype and none for the blue phenotype. However, II private alleles were observed when waIleye were separated by lake, suggesting a higher differentiation among lakes than between phenotypes.
The NJ tree (Figure 2.2) revealed that the yellow and blue phenotypes do not represent monophyletic groups, allowing us to rule out the hypothesis that separation of the two phenotypes preceded the postglacial dispersal. Individuals cluster by lake (or region) and within a given lake, and individuals of the blue phenotype are generally more similar to each other and c1uster together. This result indicates that the separation between the blue and yellow phenotypes occurred in each lake from a single colonization event. The mtDNA results support this scenario: there was no variation among yellow phenotypes except for one individual from Preissac Lake. Interestingly, three distinct groups of blue phenotypes were detected in the Des Jardins Lake, one including aIl blue individuals from the eastern part of the lake while those from the western part
40
clustered into two groups. This result strongly supports the hypothesis of multiple origins of the blue phenotype, even within a single lake.
.Jç~
~
98.7 55.2N
3~ri
-
l..i
·'00 70.2'1..r
* L..J-
t<
/ / / /l
Preissac (F)l
Malartic (E)IClova (8)
l
Blanc (A)Des Jardins (C* & 0)
---Blue phenotype
---Vellow phenotype
---Sauger
0.1
Jaccard's distance
Figure 2.2: Relationships among the 51 individuals (26 yellow phenotypes, 21 blue phenotypes, and 4 sauger) as inferred from NJ computed on the Jaccard's distance. Bootstraps values are indicated. Asterisks represent the individuals caught in the western part of Lake Des Jardins.
A maximum likelihood reallocation test was first performed on two groups with the aim of discriminating blue and yellow phenotypes from ail lakes. Large proportions of both the blue (20%) and yellow (38%) phenotype were incorrectly reassigned. This confirms the NJ results, suggesting that the separation of the blue phenotypes occurred after the colonization of the yellow phenotype and independently in each lake. The test was performed again on the