Dietary conjugated α-linolenic acid (CLNA) did not improve glucose tolerance in a
neonatal pig model
--Manuscript
Draft--Manuscript Number:Full Title: Dietary conjugated α-linolenic acid (CLNA) did not improve glucose tolerance in a neonatal pig model
Article Type: Original Contribution
Keywords: conjugated linolenic acid; n-3 fatty acid; insulin resistance; pig Corresponding Author: Christian-Alexandre Castellano, Ph.D.
Research Center on Aging - Université de Sherbrooke Sherbrooke, QC CANADA
Corresponding Author Secondary Information:
Corresponding Author's Institution: Research Center on Aging - Université de Sherbrooke Corresponding Author's Secondary
Institution:
First Author: Christian-Alexandre Castellano, Ph.D. First Author Secondary Information:
Order of Authors: Christian-Alexandre Castellano, Ph.D. Jean-Patrice Baillargeon, M.D Mélanie Plourde, Ph.D. Sandie I. Briand Paul Anger, Ph.D. Alain Giguère J. Jacques Matte, Ph.D. Order of Authors Secondary Information:
Abstract: Purpose: There is an increased interest in the benefits of conjugated α-linolenic acid (CLNA) on obesity-related complications such as insulin resistance and diabetes. The aim of the study was to investigate whether a 1% dietary supplementation of mono-CLNA isomers (c9-t11-c15-18:3 + c9-t13-c15-18:3) improved glucose and lipid metabolism in neonatal pigs. Methods: Since mono-CLNA isomers combine one conjugated two-double bond system with an n-3 polyunsaturated fatty acid (PUFA) structure, the experimental protocol was designed to isolate the dietary structural characteristics of the molecules by comparing a CLNA diet with three other dietary fats: 1) conjugated linoleic acid (c9-t11-18:2 + t10-c12-18:2; CLA), 2) non-conjugated n-3 PUFA and 3) n-6 PUFA. Thirty-two piglets weaned at 3 weeks of age were distributed into the four dietary groups. Diets were isoenergetic and food intake was controlled by a gastric tube. After 2 weeks of supplementation, gastro-enteral (OGTT) and parenteral (IVGTT) glucose tolerance tests were conducted. Results: Dietary supplementation with mono-CLNA did not modify body weight/fat or blood lipid profiles (p>0.82 and p>0.57, respectively) compared with other dietary groups. Plasma glucose, insulin and C-peptide responses to OGTT and IVGTT in the CLNA group was not different from the three other dietary groups (p>0.18 and p>0.15, respectively). Compared to the non-conjugated n-3 PUFA diet, CLNA-fed animals had decreased liver composition in three n-3 fatty acids (18:3n-3; 20:3n-3; 22:5n-3) (p<0.001). Conclusions: These results suggest that providing 1% mono-CLNA is not effective in improving insulin sensitivity in neonatal pigs.
Suggested Reviewers: Hélène Poirier
Darshan S. Kelley
CLNA and glucose tolerance in neonatal pigs European Journal of Nutrition – v2013-06-21
1
Title: Dietary conjugated α-linolenic acid (CLNA) did not improve glucose tolerance in a neonatal pig model
1
2
Running title: CLNA and glucose tolerance in neonatal pigs
3
4
Christian-Alexandre Castellano1,2*, Jean-Patrice Baillargeon3, Mélanie Plourde1,3, Sandie I. Briand4,5, Paul Angers6, Alain
5
Giguère7 and J. Jacques Matte7
6
7
1Research Center on Aging, Health and Social Sciences Center, Geriatrics Institute, Sherbrooke, QC, Canada
8
2Department Physiology and Biophysics, Université de Sherbrooke, Sherbrooke, QC, Canada
9
3Department of Medicine, Division of Endocrinology, Université de Sherbrooke, Sherbrooke, QC, Canada
10
4Naturia Inc., Sherbrooke, QC, Canada
11
5Institut national de santé publique du Québec, Montréal, QC, Canada
12
6Department of Food Sciences and Nutrition, Faculty of Agriculture and Agri-Food Sciences, Université Laval, QC,
13
Canada, G1K 7P4.
14
7
Dairy and Swine Research and Development Centre, Agriculture and Agri-Food Canada, Sherbrooke, QC, Canada
15
16
17
18
19
*Corresponding author:20
Research Center on Aging, 1036 Belvedere South, Sherbrooke, (QC, Canada) J1H 4C4.
21
Tel: 1-819-780-2220 - Fax: 1-819-829-714122
E-mail: [email protected]23
24
2
Abstract
25
26
Purpose: There is an increased interest in the benefits of conjugated α-linolenic acid (CLNA) on obesity-related
27
complications such as insulin resistance and diabetes. The aim of the study was to investigate whether a 1% dietary
28
supplementation of mono-CLNA isomers (c9-t11-c15-18:3 + c9-t13-c15-18:3) improved glucose and lipid metabolism in
29
neonatal pigs. Methods: Since mono-CLNA isomers combine one conjugated two-double bond system with an n-3
30
polyunsaturated fatty acid (PUFA) structure, the experimental protocol was designed to isolate the dietary structural
31
characteristics of the molecules by comparing a CLNA diet with three other dietary fats: 1) conjugated linoleic acid
(c9-t11-32
18:2 + t10-c12-18:2; CLA), 2) non-conjugated n-3 PUFA and 3) n-6 PUFA. Thirty-two piglets weaned at 3 weeks of age
33
were distributed into the four dietary groups. Diets were isoenergetic and food intake was controlled by a gastric tube. After
34
2 weeks of supplementation, gastro-enteral (OGTT) and parenteral (IVGTT) glucose tolerance tests were conducted.
35
Results: Dietary supplementation with mono-CLNA did not modify body weight/fat or blood lipid profiles (p>0.82 and
36
p>0.57, respectively) compared with other dietary groups. Plasma glucose, insulin and C-peptide responses to OGTT and
37
IVGTT in the CLNA group was not different from the three other dietary groups (p>0.18 and p>0.15, respectively).
38
Compared to the non-conjugated n-3 PUFA diet, CLNA-fed animals had decreased liver composition in three n-3 fatty
39
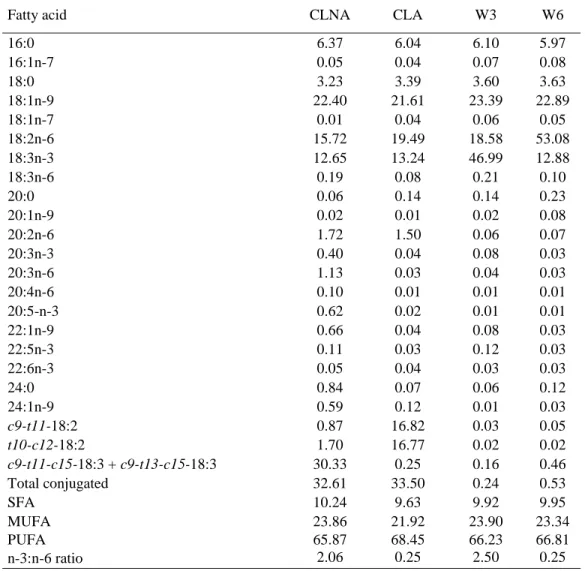
acids (18:3n-3; 20:3n-3; 22:5n-3) (p<0.001). Conclusions: These results suggest that providing 1% mono-CLNA is not
40
effective in improving insulin sensitivity in neonatal pigs.
41
42
Key words: conjugated linolenic acid: n-3 fatty acid: insulin resistance: pig
3
Introduction
44
45
Conjugated fatty acids refer to a set of positional and geometric isomers of polyunsaturated fatty acids (PUFA) with
46
conjugated double bonds. Conjugated linoleic acids (CLA) were first identified in the late eighties by Pariza et al. [1]. Since
47
then, consumption of CLA was associated to weight loss [2] and improving of insulin sensitivity [3]. Another group of
48
conjugated fatty acids recently received more attention because it combined an n-3 and a conjugated double bond:
49
conjugated α-linolenic acids (CLNA) [4]. CLNA isomers are naturally present in plant seeds (di-CLNA) and in dairy
50
products (mono-CLNA). Mono- and di-CLNA differ by their conjugated double-bond system: mono-CLNA have a single
51
conjugated double-bond system at the n-5 or n-7 carbon, i.e rumelenic acid, c9-t11-c15-18:3, whereas di-CLNA have a
52
double conjugated double-bond system at the n-5/n-7 or n-8/n-10 carbons, i.e α-eleostaric acid c9-t11-t13-18:3.
Mono-53
CLNA isomers are produced by biohydrogenation of α-linolenic acid by rumen bacteria [5, 6].
54
Because of the worldwide problem of obesity in children and the related health problems such as hypertension and diabetes
55
mellitus [7], there is an increased interest in the development of preventive and therapeutic strategies for improving insulin
56
resistance [8]. Indeed, using antidiabetic drugs is not appropriate for treating diabetes in children unless there is severe
57
glucose intolerance, thereby finding natural strategies such as CLNA isomers is an attractive approach which deserves to be
58
studied. Di-CLNA are able to decrease body weight [9] and fat[10, 11], as well as increase insulin sensitivity in rodents [9,
59
12, 13]. Some studies also reported that either CLA [14] or n-3 PUFA [15] or the combination of the two [16] could
60
improve glucose tolerance. Since a mono-CLNA such as c9-t11-c15-18:3 isomer combines the conjugated double bond
61
system of CLA and the n-3 double bond of α-linolenic acid, it is reasonable to speculate that this original fatty acid structure
62
may provide similar or even enhanced glucose tolerance than the conjugated or n-3 double bond structure.
63
Bioavailability of mono-CLNA was reported to be high in rodents. The metabolism of mono-CLNA has already been
64
studied in different animal models excluding pigs [5, 6, 17].
65
The objective of the present study was to investigate whether dietary supplementation with mono-CLNA (c9-t11-c15-18:3 +
66
c9-t13-c15-18:3) improves glucose metabolism in neonatal piglets. In order to isolate the role of the conjugated
double-67
bond in combination with the n-3 PUFA structure, mono-CLNA group will be compared with three other dietary treatments:
68
CLA isomers, non-conjugated n-3 and non-conjugated n-6 PUFAs.
69
70
Methods and materials
71
72
Animals and diets
73
Thirty two Yorkshire × Landrace × Duroc piglets (females and castrated males) weaned at 3 weeks of age were separated
74
into eight groups of four animals. To control for food intake, an oesophageal gastric tube was installed into all animals as
75
previously described by Cortamira et al. [18]. Individual adjoining metabolism cages with plastic floors allowed free
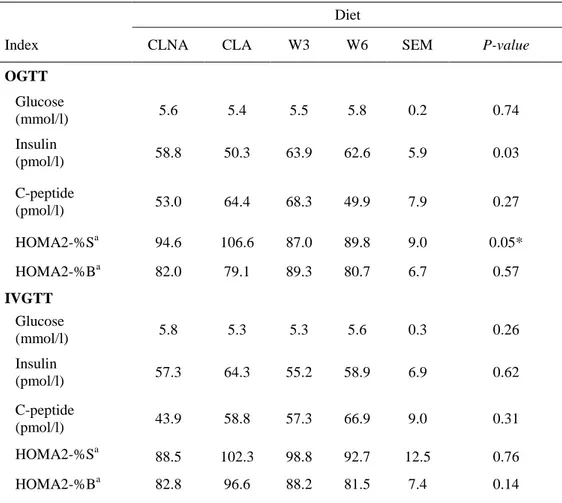
76
movement and room temperature was kept at 27°C. At baseline, average body weight was 7.6 ± 0.4 kg. Within each group
77
the piglets (from the same litter) were assigned to one of four dietary treatment groups, fed entirely with a commercial diet
78
(barley (25%), maize (20%), dried whey (20%), soybean meal (10%), extruded soybean (8%) and plasma protein (5%);
79
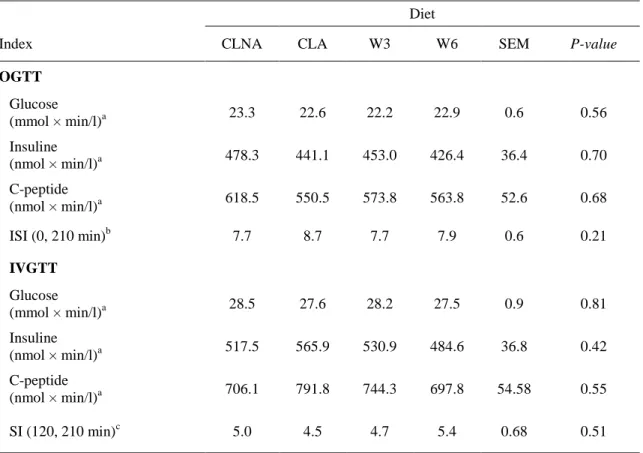
Table 1) plus either 1% of the caloric intake of the basal diet in the form of one the following lipid emulsion of specific fatty
4
acids: (1) synthetic mixture of two mono-conjugated α-linolenic acids (c9-t11-c15-18:3 + c9-t13-c15-18:3; CLNA); (2)
81
mixture of two conjugated linoleic acids (c9-t11-18:2 + t10-c12-18:2; CLA); (3) n-3 fatty acids (W3); or (4) n-6 fatty acids
82
(W6).
83
Mono-CLNA isomers were synthesized by alkali isomerization of α-linolenic acid. Thereafter, CLNA isomers were purified
84
by preparative chromatography using a reverse phase column, as previously described by Trottier [19]. The fatty acid
85
profile of the four lipid emulsions is shown in Table 2.
86
All diets were isoenergetic. The feeding regime was based on daily increment of 7.1 g feed per kg0.75 body weight (g/kg0.75)
87
up to the target maximum daily value of 56 g/kg0.75. The daily intake was adjusted three times per week according to
88
changes in body weight in order to maintain the weight stable.Basal diet was mixed with water (1:2) and infused with a
89
syringe into the stomach via the gastric tube. Daily meals were given at 08:00, 11:30 and 16:00 hours; each representing
90
45%, 20% and 35% of the diet caloric intake, respectively. Dietary treatments were given during the morning meal. Before
91
the morning meal on day 0 (before attribution of treatments) and on day 15 post-weaning, body weight was measured and
92
blood samples were collected via jugular venipuncture as previously described by Matte et al. [20].
93
On day 9 of the experimental protocol, a jugular catheter was installed by a non-surgical technique described by Matte et al.
94
[21]. All animals were tested for insulin resistance by two glucose tolerance tests: a gastro-enteral (OGTT) and a parenteral
95
(IVGTT) test. Briefly, after fasting for 18h, piglets (n=32) were given either an oral (OGTT) or an IV (IVGTT) dose of
96
glucose (1.0g/kg BW) over a period of 120 min. Blood samples were collected every 30 min for 240 min (0, 30, 60, 90, 120,
97
150, 180, 210 and 240 min) starting after the initial glucose infusion. Blood samples were centrifuged at 3000 rpm for 10
98
min at 4°C and plasma was stored at -20°C until glucose, insulin and C-peptide analyses were performed. Two days after
99
the first glucose test, the protocol was repeated with the other glucose test using the same animal. All animals were
100
sacrificed on day 17. The liver was removed, weighed and samples were stored at -20°C for further analysis. The digestive
101
tract, brain, lungs and heart were removed and stored at –20°C until lipid quantification was performed.
102
Throughout the experimental protocol animals were cared for according to the recommended code of practice of Agriculture
103
Canada [22] and the procedure was approved by the local Animal Care Committee following the guidelines of the Canadian
104
Council on Animal Care [23].
105
106
Biochemical analyses
107
Plasma glucose was measured by an enzymatic colorimetric assay (GLU GOD-PAP; Roche Diagnostics, Indianapolis, IN,
108
USA) whereas insulin (Porcine Insulin RIA Kit PI-12K; Linco Research Inc., St Charles, MI USA) and C-peptide (Porcine
109
C-peptide RIA kit PCP-22k; Linco Research Inc.) were assayed by commercial RIA kits. The homeostatic model
110
assessment (HOMA2), described by Levy et al. [24] was used to estimate insulin sensitivity (HOMA2-%S) and secretion
111
(HOMA2-%B) from baseline plasma parameters measured during OGTT and IVGTT. The area under the curve (AUC) of
112
glucose, insulin and C-peptide were calculated using the trapezoidal method [25] between 0 and 210 min. Matsuda’s insulin
113
sensitivity whole-body index (ISI) was also calculated from the data generated during the OGTT [26]. An insulin sensitivity
114
index (SI) derived from the IVGTT was calculated according to a modified method described by Bergman and colleagues
115
[27, 28].
5
SI was determined from 120 to 210 min of the IVGTT, i.e. after the end of the infusion to assess the deconvolution of
117
glucose with regards to insulin after the glucose peak. Total cholesterol, triglyceride and non-esterified fatty acid (NEFA)
118
concentration in plasma at day 1 and day 15 were measured by the service diagnostic of the Faculty of Veterinary Medicine
119
at the Université de Montréal (Montreal, QC, Canada). The fatty acid composition of the four diets and liver was
120
determined by gas chromatography as previously described by Castellano et al. [29].
121
122
Statistical Analysis
123
According to results obtained from similar porcine studies [15], the calculated sample size per group (n = 8) was sufficient
124
to detect a difference in insulin sensitivity of at least 15% with a power of 80% and a level of significance of 0.05.
125
The data were analyzed by using the MIXED procedure implemented in Statistical Analysis Systems software (version 6.11
126
of SAS, Cary, NC, USA) [30] according to a completely randomised design with four treatments (CLNA, CLA, W3, and
127
W6) as the main factor. The piglet was considered as the experimental unit. The following model was used:
128
Yij = µ + Fi + eij
129
where Yij is the dependent variable, µ is the overall mean, Fi is the treatment effect and eij is the residual error. Comparisons
130
among treatments were done using the following a priori contrasts (CLNA vs. W3 for CLA properties; CLNA vs. W6 for
131
both CLA and n-3 PUFA properties; CLNA vs. CLA for n-3 PUFA properties) using a Dunnett’s correction. All values are
132
presented as mean ± SEM and differences are considered significant at p<0.05.
133
134
Results
135
136
Anthropometry and blood lipid parameters
137
Body weight of piglets pre-treatment was 7.6 ± 0.4 kg. After 2 weeks of supplementation (day 15), there was no difference
138
(p>0.82) between treatments for either body weight (10.2 ± 0.4 kg) or fat content (10.0 ± 0.8 %). Total cholesterol,
139
triglyceride and NEFA concentrations in blood plasma for day 1 vs. day 15 were, 5.9 ± 0.8 vs. 2.0 ± 0.1 mmol/l, 0.7 ± 0.1
140
vs. 0.3 ± 0.1 mmol/l and 708.3 ± 146.7 vs. 769.6 ± 101.8 μmol/l, respectively. There was no difference (p>0.34) according
141
to dietary treatments for these parameters.
142
143
Liver fatty acid profile
144
There was no significant difference (p=0.67) in liver weight between dietary treatments. The overall organ weight was 248
145
± 14 g. Evaluation of liver fatty acid composition was used as an indicator of whole body fatty acid status modification from
146
dietary treatments. Mono-CLNA (c9-t11-c15-18:3 + c9-t13-c15-18:3) liver content, was higher (p<0.001) in the CLNA diet
147
than the other diets (0.25 vs. 0.01 g/100g fatty acids, respectively). With regards to n-3 fatty acids, 18:3n-3, 20:3n-3 and
148
22:5n-3 were 20 to 40% lower (p<0.001) in CLNA diets compared to the W3 diet. Total n-3 PUFA was also significantly
149
lower (p<0.001) in the CLNA diet than the W3 diet (9.27 vs. 10.77 g/100g fatty acids, respectively). In contrast, the
150
proportion of arachidonic acid (20:4n-6) was 8% higher in the CLNA group compared to the W3 group, resulting in a
151
significantly higher total n-6 PUFA level (CLNA vs. W3, 40.61 vs. 39.52 g/100g fatty acids, respectively).
152
153
6
Basal plasma glucose, insulin and C-peptide concentration
154
Baseline plasma glucose, insulin and C-peptide concentrations were evaluated before the OGTT or IVGTT load of glucose
155
(Table 3). A significant treatment effect was detected for fasting insulin concentration on the OGTT day (p=0.03) and on
156
calculated insulin sensitivity HOMA-%S but the specific contrast test did not allow discrimination between the CLNA
157
group and the other dietary groups (p>0.22). There was no other treatment difference for OGTT and IVGTT (p>0.14; Table
158
3).
159
160
Monitoring of glucose, insulin and C-peptide during OGTT and IVGTT
161
AUC for glucose, insulin and C-peptide monitored between 0 and 210 min are reported in Table 4. Among the four dietary
162
groups, there was no significant difference in glucose, insulin and C-peptide monitoring over the OGTT and the IVGTT.
163
Dietary intake did not improve the Matsuda’s ISI (p=0.71) calculated from the OGTT nor the minimal model-derived
164
insulin sensitivity calculated from the IVGTT (SI; p=0.51).
165
166
Discussion
167
168
The present study aimed to investigate whether dietary supplementation with mono-CLNA improves body composition and
169
glucose tolerance in neonatal piglets.
170
This model was chosen because piglets represent 1) an accelerated model of postnatal development to study human neonatal
171
nutrition and development [31], 2) a relevant model for insulin resistance [32] and 3) a suitable model for evaluating
172
nutritional strategies to enhance glucose tolerance and prevent type 2 diabetes and cardiovascular diseases later in life [33].
173
174
Body composition and blood lipids
175
Mono-CLNA might combine anti-obesity properties of CLA, along with those of the α-linolenic acid. There is evidence
176
suggesting that CLA decreases body weight, fat accumulation and improves serum lipids in mice [34], rats [35], hamsters
177
[36] and humans [37]. Similarly, α-linolenic acid was reported to improve the same biomarkers in hamsters [38] and
178
humans [39]. Many studies in rodents [10-12, 40-42] showed that dietary di-CLNA supplementation decreases body
179
weight, body fat as well as plasma triglycerides and cholesterol. This study speculates that because mono-CLNA has an
180
original structure compared to di-CLNA, this conjugated fatty acid will have improved or equally effective glucose
181
tolerance than CLA or α-linolenic acid alone. None of them combined an n-3 PUFA and a CLA structure. However, dietary
182
supplementation of piglets with mono-CLNA for 14 days did not improve body weight, body fat nor blood lipid profiles.
183
Our findings extend previous studies in rodents which reported that dietary mono-CLNA did not lower body weight [17,
184
43]. By analogy, dietary di-CLNA isomers did not lower adipose tissue weight as well as total cholesterol and triglycerides
185
in the plasma of animals [12, 17, 42, 44, 45] and humans [46].
186
187
Liver fatty acid composition
188
CLNA and CLA concentrations in liver reflected the dietary intake of these fatty acids. This response suggests that a
2-189
week supplementation was sufficient for stabilization of PUFA status within the piglet’s body. Our results are in the line
7
with Chartrand et al. [47] who reported that dietary fatty acid content consumed for at least 14 days was proportional to
191
plasma fatty acid profiles and remained constant up to study completion at 36 days. Our results also showed that giving
192
mono-CLNA to piglets changed the n-3 and n-6 fatty acid balance. More specifically, compared to the W3 diet, feeding
193
mono-CLNA for 2 weeks decreased the proportions of 18:3n-3, 20:3n-3, 20:5n-3, 22:5n-3 together with an increase in
194
20:4n-6 and total n-6 PUFA content. Since a previous study in mice reported that n-3 PUFA chronic depletion in the liver
195
led to the development of hepatic insulin resistance over a 3 month period [48], further studies need to be carried out for an
196
extended period of time in pigs fed CLNA with additional health indicators (blood biochemical, other tissue lipid profile,
197
etc) in order to better assess the safety aspects of consuming dietary mono-CLNA isomers.
198
199
Glucose tolerance
200
One of our hypotheses was that combining a conjugated and an n-3 PUFA structure in one fatty acid, like mono-CLNA,
201
would improve insulin sensitivity considering that dietary CLA seems to lower insulinemia in rats [49] and α-linolenic acid
202
intake seems to reduce insulin resistance in rats [50] and in humans [51].
203
Our results suggest a significant treatment effect for fasting insulin concentration and insulin sensitivity on the OGTT day
204
(p=0.03). However, we consider that this result is unlikely of biological significance because specific a priori contrasts
205
indicate that there is no treatment effect of mono-CLNA diet compared to the three other dietary groups. Moreover, these
206
differences were not confirmed at the day of IVGTT or during the two glucose tolerance tests. One possible explanation
207
could be a higher insulin secretion generated by environmental/psychological stresses during the first experiment, i.e.
208
human presence, handling, noise, etc [52, 53]. Also, no treatment effect was seen on C-peptide, a good indicator of
209
endogenous insulin secretion [54], either for OGTT or IVGTT.
210
In regards to the different indexes of insulin sensitivity (HOMA2-%S, ISIMatsuda and SI) and insulin secretion (Insulin and
C-211
peptide levels, AUCs and HOMA2-%B), none were improved by ingestion of mono-CLNA for 2 weeks.
212
Results on a closer structural analog to mono-CLNA such as di-CLNA showed some inconsistency. Indeed, although
213
several studies reported that dietary supplementation with di-CLNA isomers can decrease type 2 diabetes risk [12] and
214
improve glucose tolerance [9, 55] in mice, others reported an increase in insulin resistance (HOMA-IR index) in rats [44]
215
similar to what is reported in mice [56, 57], pigs [58], and humans [59]. Moreover, most of the studies using n-3 PUFA
216
supplement in humans failed to improve insulin sensitivity [60, 61].
217
218
Limitations of the present study
219
The present study extends previous findings [9, 40, 44, 55] using a neonatal pig model and randomised experimental design
220
to compare mono-CLNA diet vs. three other dietary treatments (CLA, W3, and W6). Nevertheless, it has some limitations
221
including the duration of the supplementation and the composition of the CLA diet, since we used a mixture of two isomers:
222
c9-t11-18:3 + t10-c12-18:3. Even if most previous studies have used a CLA isomer mixture, recent findings show that
223
purified CLA isomers could have opposite actions on glucose tolerance, with t10-c12-18:3 reducing insulin sensitivity and
224
c9-t11-18:3 enhancing insulin tolerance [3]. Mono-CLNA was also a mixture of two isomers and this is mostly because it is
225
not possible to cost effectively separate the two CLNA isomers for generating high doses of single CLNA isomers for
8
feeding animals. Therefore, a direct comparison between CLA and CLNA diets based only on the chemical structure is
227
limited.228
229
Conclusions230
This study showed that mono-CLNA, combining conjugated and an n-3 double-bound structure, did not provide additive
231
improvement for body composition, glucose tolerance or blood lipid profile in the neonatal piglet model supplemented for a
232
period of 2 weeks. Conversely, mono-CLNA decreased total n-3 PUFA in liver, a finding which merits consideration in
233
regards to neonatal development and safety.
234
235
Acknowledgments
236
The authors are grateful to M. Guillette for her invaluable technical assistance; to Mélanie Turcotte and the animal care
237
team under supervision of D. Morrissette; and to S. Methot for his help with statistical analysis. The authors are also
238
grateful to Fonds u b cois de echerche sur la Nature et les Technologies (FQRNT) and Naturia Inc (Sherbrooke,
239
Canada) for a Ph.D scholarship to M. P at the time where the experiment was carried-out and to Fonds de recherche
240
Québec-santé for a current Junior 1 salary award. Mono-CLNA were generously provided by Naturia Inc. (Sherbrooke, QC,
241
Canada). The financial support was provided by Naturia Inc and the Mathing Investment Initiative of Agriculture and
Agri-242
Food Canada. None of the authors has any financial conflict of interest.
243
244
References
245
1. Pariza MW, Ha YL (1990) Conjugated dienoic derivatives of linoleic acid: a new class of anticarcinogens. Med
246
Oncol Tumor Pharmacother 7:169-171.
247
2. Plourde M, Jew S, Cunnane SC, Jones PJ (2008) Conjugated linoleic acids: why the discrepancy between animal and
248
human studies? Nutr Rev 66:415-421. doi:10.1111/j.1753-4887.2008.00051.x
249
3. Bhattacharya A, Banu J, Rahman M, Causey J, Fernandes G (2006) Biological effects of conjugated linoleic acids in
250
health and disease. J Nutr Biochem 17:789-810. doi:10.1093/ilar.47.3.243
251
4. Hennessy AA, Ross RP, Devery R, Stanton C (2011) The health promoting properties of the conjugated isomers of
252
α-linolenic acid. Lipids 46:105-119. doi:10.1007/s11745-011-3636-z
253
5. Plourde M, Destaillats F, Chouinard PY, Angers P (2007) Conjugated α-linolenic acid isomers in bovine milk and
254
muscle. J Dairy Sci 90:5269-5275. doi:10.3168/jds.2007-0157
255
6. Destaillats F, Berdeaux O, Sébédio JL, Juaneda P, Grégoire S, Chardigny JM, Bretillon L, Angers P (2005)
256
Metabolites of conjugated isomers of α-linolenic acid (CLnA) in the rat. Journal of Agricultural and Food Chemistry
257
53:1422-1427. doi:10.1021/jf0481958
9
7. Levy-Marchal C, Arslanian S, Cutfield W, Sinaiko A, Druet C, Marcovecchio ML, Chiarelli F, Amemiyia S,
259
Berenson G, Caprio S, et al. (2010) Insulin resistance in children: Consensus, perspective, and future directions.
260
Journal of Clinical Endocrinology and Metabolism 95:5189-5198. doi:10.1210/jc.2010-1047
261
8. World Health Organization (2000) Obesity: preventing and managing the global epidemic. In: Consultation RoaW
262
(ed) WHO Technical Report Series. World Health Organization, Geneva, p 252.
263
9. Vroegrijk IOCM, van Diepen JA, van den Berg S, Westbroek I, Keizer H, Gambelli L, Hontecillas R,
Bassaganya-264
Riera J, Zondag GCM, Romijn JA, et al. (2011) Pomegranate seed oil, a rich source of punicic acid, prevents
diet-265
induced obesity and insulin resistance in mice. Food Chem Toxicol 49:1426-1430. doi:10.1016/j.fct.2011.03.037
266
10. Koba K, Akahoshi A, Yamasaki M, Tanaka K, Yamada K, Iwata T, Kamegai T, Tsutsumi K, Sugano M (2002)
267
Dietary conjugated linolenic acid in relation to CLA differently modifies body fat mass and serum and liver lipid
268
levels in rats. Lipids 37:343-350.
269
11. Saha SS, Chakraborty A, Ghosh S, Ghosh M (2012) Comparative study of hypocholesterolemic and hypolipidemic
270
effects of conjugated linolenic acid isomers against induced biochemical perturbations and aberration in erythrocyte
271
membrane fluidity. Eur J Nutr 51:483-495. doi:10.1007/s00394-011-0233-0
272
12. McFarlin BK, Strohacker KA, Kueht ML (2009) Pomegranate seed oil consumption during a period of high-fat
273
feeding reduces weight gain and reduces type 2 diabetes risk in CD-1 mice. Br J Nutr 102:54-59.
274
doi:10.1017/S0007114508159001
275
13. Al-Muammar MN, Khan F (2012) Obesity: The preventive role of the pomegranate (Punica granatum). Nutrition
276
28:595-604. doi:10.1016/j.nut.2011.11.013
277
14. Henriksen EJ, Teachey MK, Taylor ZC, Jacob S, Ptock A, Kramer K, Hasselwander O (2003) Isomer-specific
278
actions of conjugated linoleic acid on muscle glucose transport in the obese Zucker rat. Am J Physiol Endocrinol
279
Metab 285:E98-E105. doi:10.1152/ajpendo.00013.2003
280
15. Behme MT (1996) Dietary fish oil enhances insulin sensitivity in miniature pigs. J Nutr 126:1549-1553.
281
16. Winzell MS, Pacini G, Ahrén B (2006) Insulin secretion after dietary supplementation with conjugated linoleic acids
282
and n-3 polyunsaturated fatty acids in normal and insulin-resistant mice. Am J Physiol Endocrinol Metab
290:E347-283
E354. doi:10.1152/ajpendo.00163.2005
284
17. Plourde M, Ledoux M, Grégoire S, Portois L, Fontaine JJ, Carpentier YA, Angers P, Chardigny JM, Sébédio JL
285
(2007) Adverse effects of conjugated alpha-linolenic acids (CLnA) on lipoprotein profile on experimental
286
atherosclerosis in hamsters. Animal 1:905-910. doi:10.1017/S1751731107000079
10
18. Cortamira NO, Seve B, Lebreton Y, Ganier P (1991) Effect of dietary tryptophan on muscle, liver and whole-body
288
protein synthesis in weaned piglets: Relationship to plasma insulin. Br J Nutr 66:423-435.
289
doi:10.1079/BJN19910045
290
19. Trottier JP (2005) Synthèse et purification d'isomères conjugués des acides linoléique et α-linolénique. In: Faculté
291
des sciences de l'argiculture et de l'alimentation. Université Laval, Quebec, p 78.
292
20. Matte J, Girard C, Sève B (1986) Importance of folic acid administred during gestation on haematological status of
293
piglets. Can J Anim Sci 66:523-527.
294
21. Matte JJ (1999) A rapid and non-surgical procedure for jugular catheterization of pigs. Laboratory Animals
33:258-295
264.
296
22. Agriculture Canada (1993) Recommanded Code of Practice for Care and Handling of Pigs. In: Publication no.
297
1771E. Agriculture Canada, Ottawa, Ont., Canada
298
23. Canadian Council on Animal Care (1993) Guide to the Care and Use of Experimental Animals. In: Vol. 1. Cavadian
299
Council on Animal Care, Ottawa, Ont., Canada
300
24. Levy JC, Matthews DR, Hermans MP (1998) Correct homeostasis model assessment (HOMA) evaluation uses the
301
computer program. Diabetes Care 21:2191-2192.
302
25. Allison DB, Paultre F, Maggio C, Mezzitis N, Pi-Sunyer FX (1995) The use of areas under in diabetes research.
303
Diabetes Care 18:245-250.
304
26. Matsuda M, DeFronzo RA (1999) Insulin sensitivity indices obtained from oral glucose tolerance testing:
305
Comparison with the euglycemic insulin clamp. Diabetes Care 22:1462-1470.
306
27. Bergman RN, Phillips LS, Cobelli C (1981) Physiologic evaluation of factors controlling glucose tolerance in man.
307
Measurement of insulin sensitivity and β-cell glucose sensitivity from the response to intravenous glucose. J Clin
308
Invest 68:1456-1467.
309
28. Boston RC, Stefanovski D, Moate PJ, Sumner AE, Watanabe RM, Bergman RN (2003) MINMOD Millennium: A
310
Computer Program to Calculate Glucose Effectiveness and Insulin Sensitivity from the Frequently Sampled
311
Intravenous Glucose Tolerance Test. Diabetes Technol Ther 5:1003-1015. doi:10.1089/152091503322641060
312
29. Castellano CA, Audet I, Laforest JP, Chouinard Y, Matte JJ (2010) Fish oil diets do not improve insulin sensitivity
313
and secretion in healthy adult male pigs. Br J Nutr 103:189-196. doi:10.1017/S0007114509991590
314
30. Littell RC, Henry PR, Ammerman CB (1998) Statistical Analysis of Repeated Measures Data Using SAS
315
Procedures. J Anim Sci 76:1216-1231.
11
31. Puiman P, Stoll B (2008) Animal models to study neonatal nutrition in humans. Current Opinion in Clinical
317
Nutrition and Metabolic Care 11:601-606. doi:10.1097/MCO.0b013e32830b5b15
318
32. Christoffersen B, Ribel U, Raun K, Golozoubova V, Pacini G (2009) Evaluation of different methods for assessment
319
of insulin sensitivity in Göttingen minipigs: Introduction of a new, simpler method. American Journal of Physiology
320
- Regulatory Integrative and Comparative Physiology 297:R1195-R1201. doi:10.1152/ajpregu.90851.2008
321
33. Bellinger DA, Merricks EP, Nichols TC (2006) Swine models of type 2 diabetes mellitus: Insulin resistance, glucose
322
tolerance, and cardiovascular complications. ILAR Journal 47:243-258.
323
34. Park Y, Albright KJ, Liu W, Storkson JM, Cook ME, Pariza MW (1997) Effect of conjugated linoleic acid on body
324
composition in mice. Lipids 32:853-858.
325
35. Noguchi R, Yasui Y, Suzuki R, Hosokawa M, Fukunaga K, Miyashita K (2001) Dietary effects of bitter gourd oil on
326
blood and liver lipids of rats. Arch Biochem Biophys 396:207-212. doi:10.1006/abbi.2001.2624
327
36. Gavino VC, Gavino G, Leblanc MJ, Tuchweber B (2000) An isomeric mixture of conjugated linoleic acids but not
328
pure cis- 9,trans-11-octadecadienoic acid affects body weight gain and plasma lipids in hamsters. J Nutr 130:27-29.
329
37. Smedman A, Vessby B (2001) Conjugated linoleic acid supplementation in humans - Metabolic effects. Lipids
330
36:773-781.
331
38. Yang L, Leung KY, Cao Y, Huang Y, atnayake WMN, Chen ZY (2005) α-linolenic acid but not conjugated
332
linolenic acid is hypocholesterolaemic in hamsters. Br J Nutr 93:433-438. doi:10.1079/bjn20041365
333
39. Baxheinrich A, Stratmann B, Lee-Barkey YH, Tschoepe D, Wahrburg U (2012) Effects of a rapeseed oil-enriched
334
hypoenergetic diet with a high content of α-linolenic acid on body weight and cardiovascular risk profile in patients
335
with the metabolic syndrome. Br J Nutr 108:682-691. doi:10.1017/S0007114512002875
336
40. Hontecillas R, Diguardo M, Duran E, Orpi M, Bassaganya-Riera J (2008) Catalpic acid decreases abdominal fat
337
deposition, improves glucose homeostasis and upregulates PPA α expression in adipose tissue. Clin Nutr
27:764-338
772. doi:10.1016/j.clnu.2008.07.007
339
41. Chen PH, Chen GC, Yang MF, Hsieh CH, Chuang SH, Yang HL, Kuo YH, Chyuan JH, Chao PM (2012) Bitter
340
melon seed oil-attenuated body fat accumulation in diet-induced obese mice is associated with cAMP-dependent
341
protein kinase activation and cell death in white adipose tissue. J Nutr 142:1197-1204. doi:10.3945/jn.112.159939
342
42. Arao K, Wang YM, Inoue N, Hirata J, Cha JY, Nagao K, Yanagita T (2004) Dietary effect of pomegranate seed oil
343
rich in 9cis, 11trans, 13cis conjugated linolenic acid on lipid metabolism in obese, hyperlipidemic OLETF Rats.
344
Lipids Health Dis 3 doi:10.1186/1476-511X-3-24
12
43. Plourde M (2006) Étude des effets physiologiques des acides alpha-linoléniques conjugués. In: Sciences de
346
l'alimentation et de la nutrition. Université Laval, Quebec, p 224.
347
44. Miranda J, Fernández-Quintela A, Macarulla MT, Churruca I, García C, Rodríguez VM, Simón E, Portillo MP
348
(2009) A comparison between CLNA and CLA effects on body fat, serum parameters and liver composition. J
349
Physiol Biochem 65:25-32. doi:10.1007/BF03165966
350
45. Shinohara N, Ito J, Tsuduki T, Honma T, Kijima R, Sugawara S, Arai T, Yamasaki M, Ikezaki A, Yokoyama M, et
351
al. (2012) Jacaric acid, a linolenic acid isomer with a conjugated triene system, reduces stearoyl-CoA desaturase
352
expression in liver of mice. J Oleo Sci 61:433-441. doi:10.5650/jos.61.433
353
46. Yuan G, Sinclair AJ, Xu C, Li D (2009) Incorporation and metabolism of punicic acid in healthy young humans. Mol
354
Nutr Food Res 53:1336-1342. doi:10.1002/mnfr.200800520
355
47. Chartrand R, Matte JJ, Lessard M, Chouinard PY, Giguere A, Laforest JP (2003) Effect of dietary fat sources on
356
systemic and intrauterine synthesis of prostaglandins during early pregnancy in gilts. J Anim Sci 81:726-734.
357
48. Pachikian BD, Essaghir A, Demoulin JB, Neyrinck AM, Catry E, de Backer FC, Dejeans N, Dewulf EM, Sohet FM,
358
Portois L, et al. (2011) Hepatic n-3 polyunsaturated fatty acid depletion promotes steatosis and insulin resistance in
359
mice: Genomic analysis of cellular targets. PLoS ONE 6 doi:10.1371/journal.pone.0023365
360
49. Houseknecht KL, Heuvel JPV, Moya-Camarena SY, Portocarrero CP, Peck LW, Nickel KP, Belury MA (1998)
361
Dietary conjugated linoleic acid normalizes impaired glucose tolerance in the Zucker diabetic fatty fa/fa rat. Biochem
362
Biophys Res Commun 244:678-682. doi:10.1006/bbrc.1998.8303
363
50. Chicco AG, D'Alessandro ME, Hein GJ, Oliva ME, Lombardo YB (2009) Dietary chia seed (Salvia hispanica L.)
364
rich in a-linolenic acid improves adiposity and normalises hypertriacylglycerolaemia and insulin resistance in
365
dyslipaemic rats. Br J Nutr 101:41-50. doi:10.1017/S000711450899053X
366
51. Muramatsu T, Yatsuya H, Toyoshima H, Sasaki S, Li Y, Otsuka R, Wada K, Hotta Y, Mitsuhashi H, Matsushita K,
367
et al. (2010) Higher dietary intake of alpha-linolenic acid is associated with lower insulin resistance in middle-aged
368
Japanese. Prev Med 50:272-276. doi:10.1016/j.ypmed.2010.02.014
369
52. Armario A, Castellanos JM, Balasch J (1985) Chronic noise stress and insulin secretion in male rats. Physiology and
370
Behavior 34:359-361.
371
53. Funderburke DW, Seerley RW (1990) The effects of postweaning stressors on pig weight change, blood, liver and
372
digestive tract characteristics. Journal of Animal Science 68:155-162.
373
54. Faber OK, Binder C (1986) C-peptide: An index of insulin secretion. Diabetes/Metabolism Reviews 2:331-345.
13
55. Hontecillas R, O'Shea M, Einerhand A, Diguardo M, Bassaganya- iera J (2009) Activation of PPA γ and α by
375
punicic acid ameliorates glucose tolerance and suppresses obesity-related inflammation. J Am Coll Nutr 28:184-195.
376
doi:10.1080/07315724.2009.10719770
377
56. Tsuboyama-Kasaoka N, Takahashi M, Tanemura K, Kim HJ, Tange T, Okuyama H, Kasai M, Ikemoto S, Ezaki O
378
(2000) Conjugated linoleic acid supplementation reduces adipose tissue by apoptosis and develops lipodystrophy in
379
mice. Diabetes 49:1534-1542. doi:10.2337/diabetes.49.9.1534
380
57. Kelley DS, Vemuri M, Adkins Y, Gill SHS, Fedor D, Mackey BE (2009) Flaxseed oil prevents trans-10,
cis-12-381
conjugated linoleic acid-induced insulin resistance in mice. Br J Nutr 101:701-708.
382
doi:10.1017/S0007114508027451
383
58. Fernández-Fígares I, Lachica M, Martín A, Nieto R, Gonzáez-Valero L, Rodríguez-López JM, Aguilera JF (2012)
384
Impact of dietary betaine and conjugated linoleic acid on insulin sensitivity, protein and fat metabolism of obese
385
pigs. Animal 6:1058-1067. doi:10.1017/S1751731111002308
386
59. Risérus U, Arner P, Brismar K, Vessby B (2002) Treatment with dietary trans10cis12 conjugated linoleic acid causes
387
isomer-specific insulin resistance in obese men with the metabolic syndrome. Diabetes Care 25:1516-1521.
388
doi:10.2337/diacare.25.9.1516
389
60. Vessby B (2000) Dietary fat and insulin action in humans. Br J Nutr 83:S91-S96. doi:10.1017/S000711450000101X
390
61. McAuley K, Mann J (2006) Nutritional determinants of insulin resistance. J Lipid Res 47:1668-1676.
391
doi:10.1194/jlr.R600015-JLR200
392
393
394
14
Table 1: Composition of basal diet
395
Ingredients Calculated concentration
Digestible energy (MJ/kg) 14.98 Total protein (%) 19.94 Crude fibre (%) 1.90 Fat (%) 7.49 Lysine (%) 1.40 Methionine (%) 0.43 Tryptophan (%) 0.24 Calcium (%) 0.80 Phosphorus (%) 0.70
Provided (per kg basal diet): Mn, 40 mg; Zn, 2935 mg; Fe, 299 mg; Cu, 19 mg; I, 2 mg; Se 297 μg; vitamin A, 4.9 mg; vitamin D, 37.5 μg; vitamin E, 66.8 mg; menadione, ; thiamin, 2.7 mg; riboflavin, 8.7 mg; niacin, 31 mg; panthothenic acid 21.2 mg; folic acid, 0.7 mg; pyridoxine, 2.6 mg; biotin, 120 μg; vitamin B12, 25.1 μg; choline, 303 mg
15
Table 2: Analytical fatty acid composition (g/100 g fatty acids) of lipid emulsions added to the dietary treatments
397
Fatty acid CLNA CLA W3 W6
16:0 6.37 6.04 6.10 5.97 16:1n-7 0.05 0.04 0.07 0.08 18:0 3.23 3.39 3.60 3.63 18:1n-9 22.40 21.61 23.39 22.89 18:1n-7 0.01 0.04 0.06 0.05 18:2n-6 15.72 19.49 18.58 53.08 18:3n-3 12.65 13.24 46.99 12.88 18:3n-6 0.19 0.08 0.21 0.10 20:0 0.06 0.14 0.14 0.23 20:1n-9 0.02 0.01 0.02 0.08 20:2n-6 1.72 1.50 0.06 0.07 20:3n-3 0.40 0.04 0.08 0.03 20:3n-6 1.13 0.03 0.04 0.03 20:4n-6 0.10 0.01 0.01 0.01 20:5-n-3 0.62 0.02 0.01 0.01 22:1n-9 0.66 0.04 0.08 0.03 22:5n-3 0.11 0.03 0.12 0.03 22:6n-3 0.05 0.04 0.03 0.03 24:0 0.84 0.07 0.06 0.12 24:1n-9 0.59 0.12 0.01 0.03 c9-t11-18:2 0.87 16.82 0.03 0.05 t10-c12-18:2 1.70 16.77 0.02 0.02 c9-t11-c15-18:3 + c9-t13-c15-18:3 30.33 0.25 0.16 0.46 Total conjugated 32.61 33.50 0.24 0.53 SFA 10.24 9.63 9.92 9.95 MUFA 23.86 21.92 23.90 23.34 PUFA 65.87 68.45 66.23 66.81 n-3:n-6 ratio 2.06 0.25 2.50 0.25
SFA = total saturated fatty acids; MUFA = total monounsaturated fatty acid; PUFA = total polyunsaturated fatty acids; n-3:n-6 ratio = n-3 PUFA to n-6 PUFA ratio
16
Table 3: Basal plasma concentrations of glucose, insulin and C-peptide during OGTT or IVGTT according to the dietary
399
treatments
400
Diet
Index CLNA CLA W3 W6 SEM P-value
OGTT Glucose (mmol/l) 5.6 5.4 5.5 5.8 0.2 0.74 Insulin (pmol/l) 58.8 50.3 63.9 62.6 5.9 0.03 C-peptide (pmol/l) 53.0 64.4 68.3 49.9 7.9 0.27 HOMA2-%Sa 94.6 106.6 87.0 89.8 9.0 0.05* HOMA2-%Ba 82.0 79.1 89.3 80.7 6.7 0.57 IVGTT Glucose (mmol/l) 5.8 5.3 5.3 5.6 0.3 0.26 Insulin (pmol/l) 57.3 64.3 55.2 58.9 6.9 0.62 C-peptide (pmol/l) 43.9 58.8 57.3 66.9 9.0 0.31 HOMA2-%Sa 88.5 102.3 98.8 92.7 12.5 0.76 HOMA2-%Ba 82.8 96.6 88.2 81.5 7.4 0.14
W3 = omega-3 fatty acids diet; W6 = omega-6 fatty acids diet; CLNA = conjugated alpha-linolenic acids diet; CLA = linoleic acids diet; OGTT=oral glucose tolerance test; IVGTT = intravenous glucose tolerance test.
aCalculated insulin sensitivity (HOMA2-%S) and β-cell function (HOMA2-%B) based on homeostatic model assessment [24].
*Specific contrasts p-values for CLNA vs. CLA, CLNA vs. W3 and CLNA vs. W6 were 0.22, 0.53 and 0.81, respectively.
17
Table 4: Plasma glucose, insulin and C-peptide responses in OGTT and IVGTT
402
Diet
Index CLNA CLA W3 W6 SEM P-value
OGTT Glucose (mmol × min/l)a 23.3 22.6 22.2 22.9 0.6 0.56 Insuline (nmol × min/l)a 478.3 441.1 453.0 426.4 36.4 0.70 C-peptide (nmol × min/l)a 618.5 550.5 573.8 563.8 52.6 0.68 ISI (0, 210 min)b 7.7 8.7 7.7 7.9 0.6 0.21 IVGTT Glucose (mmol × min/l)a 28.5 27.6 28.2 27.5 0.9 0.81 Insuline (nmol × min/l)a 517.5 565.9 530.9 484.6 36.8 0.42 C-peptide (nmol × min/l)a 706.1 791.8 744.3 697.8 54.58 0.55 SI (120, 210 min)c 5.0 4.5 4.7 5.4 0.68 0.51
W3 = omega-3 fatty acids diet; W6 = omega-6 fatty acids diet; CLNA = conjugated alpha-linolenic acids diet; CLA = linoleic acids diet; OGTT = oral glucose tolerance test; IVGTT = intravenous glucose tolerance test;
aValues are AUC from 0 to 210 min during OGTT or IVGTT.
b
Insulin sensitivity index (ISI) [26] calculated as follow: ISI= (Glubasal × Insbasal × Glumean × Insmean)0.5. cMinimal model-derived insulin sensitivity index (SI) based on MINMOD Millennium [28].