Mécanismes de transport, d'agrégation et de production
du krill (Thysanoessa raschii et Meganyctiphanes
norvegica) dans l'estuaire et le golfe du Saint-Laurent
Thèse
Deborah Benkort
Doctorat interuniversitaire en océanographie
Philosophiæ doctor (Ph. D.)
MECANISMES DE TRANSPORT, D'AGREGATION ET
DE PRODUCTION DU KRILL (Thysanoessa raschii ET
Meganyctiphanes norvegica) DANS L'ESTUAIRE ET LE
GOLFE DU SAINT-LAURENT
Thèse
DÉBORAH BENKORT
Sous la direction de :
Frédéric MAPS, directeur de recherche Stéphane PLOURDE, codirecteur de recherche
Résumé
Le krill est un crustacé macrozooplanctonique qui joue un rôle clé dans l'écosystème marin pélagique. Il représente une plateforme trophique entre les producteurs primaires planctoniques et les niveaux supérieurs de la chaine alimentaire. Dans un contexte de changements environnementaux, la compréhension des impacts de la variabilité naturelle du forçage physique sur la physiologie du krill et la dynamique des populations apparait essentielle pour mieux appréhender la dynamique, l’évolution et la gestion des écosystèmes marins subarctiques. Dans l'estuaire et le golfe du Saint-Laurent (EGSL), les communautés de krill sont dominées par deux espèces sympatriques, Meganyctiphanes norvegica et
Thysanoessa raschii.
L'objectif principal de ce projet de recherche était d'étudier, à travers le développement de modèles biophysiques, les effets de la variabilité environnementale sur la distribution, la reproduction et la croissance de ces deux espèces. Le projet nous a permis i) de construire un modèle physiologique spécifique à l'espèce (IBM) pour le stade adulte des deux espèces de krill et de le valider avec les données disponibles ; ii) d'identifier spatio-temporellement les zones de croissance et de reproduction potentielles de l'EGSL pour les deux espèces iii) d'étudier le rôle de la variabilité interindividuelle sur la dynamique de production de T. raschii face à la variabilité environnementale.
Dans le premier chapitre, nous avons construit un modèle physiologique basé sur l'individu et spécifique à chaque espèce représentant de manière réaliste le cycle de vie annuel des deux espèces à la station Rimouski située dans l'estuaire du Saint-Laurent. Nous avons montré que la dynamique de production des individus de M. norvegica et de T. raschii était fortement liée à l'environnement alimentaire. Cependant, les deux espèces ont montré des trajectoires de croissance et de reproduction bien différentes, T. raschii montrant une saison de production estivale, alors que M. norvegica montrait une saison de production plutôt automnale. Le modèle a permis de mettre en évidence comment la compréhension et la mise en œuvre des processus d'alimentation étaient essentielles pour une représentation précise de la dynamique de croissance de chacune des espèces.
Dans le deuxième chapitre, nous avons couplé le modèle physiologique développé au chapitre 2 à un modèle de circulation générale 3D et à un modèle biogéochimique de type NPZD, afin d'étudier la dynamique de production à l'échelle de l'EGSL dans son ensemble. Les résultats ont montré une forte saisonnalité du potentiel de croissance et de reproduction des deux espèces, étroitement liée à la dynamique du phytoplancton et du zooplancton, représentant leur champ de proies. Bien que les connaissances soient limitées au nord-est du golfe, les résultats de notre modèle ont montré l'émergence de zones de forte production, dans l'estuaire, dans la région du détroit de Belle Isle, le long de la côte ouest de Terre-Neuve et le long de la côte nord. Notre étude a également mis en évidence l'existence d'un circuit cyclonique de production du krill longeant les rives de l'ensemble de l'EGSL, de la péninsule d'Avalon, au sud de Terre-Neuve, à la rive nord du Québec en traversant le détroit de Belle Isle, durant la saison de production (avril à octobre).
Dans le troisième chapitre, nous nous sommes intéressés à quantifier la contribution relative de la variabilité environnementale interannuelle (2003 et 2006) et celle de la variabilité interindividuelle sur la dynamique de production de T. raschii dans le système EGSL. Nos résultats ont montré qu'environ 15% de la variabilité totale des variables d'état du modèle IBM pouvait être expliquée par la variabilité interindividuelle simulée ajoutée au modèle IBM. Les résultats montrent également que l'inclusion de la variabilité interindividuelle n'a pas modifié significativement la réponse de la population en réponse à l’environnement : les mesures de centralité des variables d'état obtenues entre 2003 et 2006 sont restées similaires dans les simulations avec et sans variabilité interindividuelle. Cependant, il en est résulté un élargissement des distributions de fréquence des variables d'état. De plus, une augmentation des réponses positives des variables d'états lors des simulations avec variabilité interindividuelle a également émergé des résultats du modèle pour l'année 2003, évoquant une résistance de la population face à la variabilité de l'environnement.
Les résultats de cette thèse fournissent des données importantes pouvant être intégrées dans un plan de gestion écosystémique et constituent une base pour l'étude de la variabilité interannuelle et la compréhension de l'évolution future de la dynamique de production pour les deux espèces.
Abstract
Krill are macrozooplanktonic crustaceans play a key role in the pelagic marine ecosystem. They represent a crucial trophic platform between planktonic primary producers and the upper food-web levels. In a context of environmental changes, understanding the impacts of the natural variability of physical forcing on krill physiology and population dynamics appears essential to better apprehend the dynamics, evolution and management of subarctic marine ecosystems. In the Estuary and Gulf of St. Lawrence (EGSL), krill communities are dominated by two sympatric species, names Meganyctiphanes norvegica and Thysanoessa raschii.
The main objective of this research project was to study, through the development of biophysical models, the effects of environmental variability on the distribution, reproduction and growth of these two species. The project allowed us i) to build a species-specific physiological based model (IBM) for adult stage of both krill species and to validate it with available data; ii) to identify spatio-temporally the potential growth and reproduction areas in the EGSL for both species iii) to study the role of the intraspecific variability on T. raschii population dynamics facing the environmental variability.
In the first chapter we built a physiological species-specific based model representing realistically the annual adult life cycle for both dominant species at the Rimouski station located in the St. Lawrence River Estuary. We showed that the production dynamics of M. norvegica and T. raschii individuals were strongly linked to the feeding environment. However, both species exhibited well different growth and reproductive trajectories, T. raschii showing a summer production season, while M. norvegica an autumnal production season. The model highlighted how understanding and implementing feeding processes was essential for accurate representation of the growth dynamics of each species.
In the second chapter, we coupled the physiological model developed in Chapter 2 with a 3D general circulation model and a biogeochemical model of the NPZD type, in order to study the production dynamics at the EGSL scale. The results showed a strong seasonality of the growth and reproduction potential of both species, closely related to the dynamics of phytoplankton and zooplankton, representing their prey field. Although knowledge was
limited in the northeastern Gulf, model results showed the emergence of high production areas in the Estuary, in the Strait of Belle Isle, along the western coast of the Newfoundland and along the North Shore. Our study also highlighted the existence of a larger cyclonic circuit of krill production and transport that runs along the shores of the whole EGSL, from the Avalon Peninsula in the south of Newfoundland to the North Shore of Québec across the Strait of Belle Isle, and back to the Estuary upstream during the production season (April to October).
In the third chapter, we are interested in quantifying the relative contribution of interannual environmental variability (2003 and 2006) and inter-individual variability on the production dynamics of T. raschii in the EGSL system. Our results showed that about 15% of the variability of IBM model state variables could be explained by the level of simulated inter-individual variability. Results also showed that inclusion of inter-individual variability did not significantly modify the system's response at the population level in response to the environment: the centrality measures of the state variables obtained between 2003 and 2006 remained similar in the simulations with and without inter-individual variability. However, this has resulted in an expansion of the frequency distributions of the state variables. Moreover, an increase in the positive responses of the state variables during simulations with inter-individual variability also emerged from the results of the model for the year 2003, evoking a resistance of the population to the variability of the environment.
These results provide important data to integrate in fully ecosystem-based management plan. They represent a baseline to the study of the inter-annual variability and the understanding of the future production dynamics evolution for the both species.
Résumé ... iii
Abstract ... v
Liste des figures ... x
Liste des tableaux ... xv
Liste des sigles ... xvi
Remerciements ... xviii
Avant-propos ... xx
Introduction générale ... 1
I Chapitre 1 - Individual-based modelling explains the contrasted seasonality in size, growth and reproduction of the sympatric Arctic (Thysanoessa raschii) and Nordic krill (Meganyctiphanes norvegica) in the St. Lawrence estuary, eastern Canada. ... 14
I.1 Résumé ... 14
I.2 Abstract ... 15
I.3 Introduction ... 16
I.4 Methods ... 18
I.4.1 Study area and environmental data sampling ... 18
I.4.2 Environmental forcing climatologies ... 20
I.4.3 Krill sampling and derived data ... 23
I.4.4 Modelling physiological processes ... 24
I.4.4.1 Moulting ... 24
I.4.4.2 Growth ... 26
I.4.4.3 Reproduction ... 28
I.4.5 Model parameterization, calibration and validation ... 33
I.4.6 Simulations and sensitivity analyses ... 34
I.5 Results ... 34
I.5.1 Environmental forcing at Rimouski station ... 34
I.5.2 Growth and body size dynamics ... 35
I.5.3 Reproduction dynamics ... 42
I.5.4 Model verification and validation ... 43
I.5.5 Sensitivity analysis ... 46
I.6 Discussion ... 48
I.6.1 Adult krill production dynamics ... 48
I.6.3 Recommendations and future direction ... 52
II Chapitre 2 - Arctic krill (Thysanoessa raschii) and Nordic krill (Meganyctiphanes norvegica) circuits of production revealed by the interactions between their physiology, swimming behaviour and the circulation in the subarctic Gulf of St. Lawrence, Canada ... 54
II.1 Résumé ... 54
II.2 Abstract ... 55
II.3 Introduction ... 56
II.4 Methods ... 59
II.4.1 General circulation model ... 59
II.4.2 Biogeochemical model ... 60
II.4.3 Lagrangian transport ... 61
II.4.4 Individual based physiological model ... 62
II.4.5 Numerical experiments ... 65
II.4.6 Cluster analysis ... 66
II.5 Results ... 66
II.5.1 Environmental forcing in the EGSL ... 66
II.5.2 Spatiotemporal dynamic of the "Static" simulations ... 68
II.5.2.1 Patterns of growth potential and egg production ... 68
II.5.2.2 Physiological clusters ... 73
II.5.3 Spatiotemporal dynamic of the "Transport" simulations ... 76
II.5.3.1 Physiological clusters ... 76
II.5.3.2 Circuits of krill production ... 76
II.6 Discussion ... 85
II.6.1 M. norvegica and T. raschii production dynamics ... 85
II.6.2 Krill aggregations: production sources or sinks ... 88
II.6.3 Ecological and management implications ... 90
II.7 Conclusion ... 93
III Chapitre 3 - Relative contribution of interannual environmental variability and inter-individual variability to the production dynamics of Arctic krill (Tysanoessa raschii) in the sub-Arctic Gulf of St. Lawrence, eastern Canada: insights from a numerical modelling experiment. 94 III.1 Résumé ... 94
III.2 Abstract ... 95
III.3 Introduction ... 96
III.4.1 General circulation model ... 99
III.4.2 Biogeochemical model ... 99
III.4.3 Lagrangian transport ... 100
III.4.4 Physiological model ... 100
III.4.5 Numerical experiments ... 101
III.4.6 Inter-individual "paramosome" generation ... 102
III.4.7 Cluster analysis ... 105
III.4.8 Canonical Correlation Analysis (CCorA) ... 105
III.5 Results ... 106
III.5.1 Interannual forcing variability in the EGSL ... 106
III.5.2 Effects of interannual variability on T. raschii production ... 108
III.5.3 Effects of inter-individual variability on T. raschii production ... 109
III.6 Discussion ... 116
III.6.1 Plastic phenotypic responses of T. raschii to environmental variability .... 117
III.6.2 Effects of genotypic diversity on T. raschii production dynamics ... 118
III.6.3 Relationships between genotypic diversity and phenotypic plasticity of T. raschii 120 III.6.4 Limitations and future directions ... 121
Conclusion générale ... 124
Liste des figures
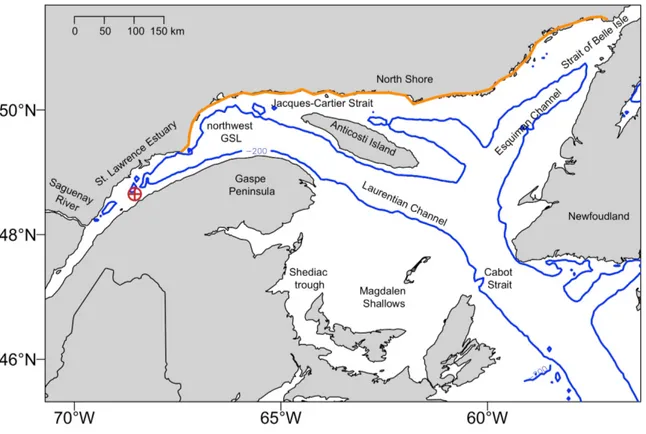
Figure 0.1 Carte du domaine d'étude, le golfe et l'estuaire du Saint-Laurent, montrant l'isobathe 200 m (ligne bleue), les principaux chenaux (Laurentien, Esquiman) et détroit (Cabot, Belle Isle, Jacques-Cartier). La croix cerclée rouge représente la station Rimouski et la ligne orange représente la délimitation de la Côte-Nord (de Pointe-des-Monts au sud à Blanc-Sabon au nord). ... 4 Figure 0.2 Meganyctiphanes norvegica adulte (a) et Thysanoessa raschii adulte (b). Les dessins
représentatifs sont tirés de Marine Species Identification Portal ( http://species-identification.org/index.php). ... 6 Figure 0.3 Représentation schématique du réseau trophique de l’estuaire et du golfe du Saint-Laurent
centré sur le krill (adapté de Harvey et al. 2003). ... 7 Figure 0.4 Schéma conceptuel de la structure de la thèse. Intégration des trois chapitres en réponse à
la problématique générale dans un contexte spatiotemporel. Les couleurs intègrent le niveau organisationnel de la population du jaune représentant l'individu au rose représentant la population. ... 13 Figure I.1 Bathymetric map of the Estuary and Gulf of St. Lawrence showing the 300 m (thick line)
and 100 m isobaths (dashed line), as well as the Rimouski station (red target). NF means Newfoundland. ... 20 Figure I.2 Environmental forcing fields. (a) Temperature amplitude (°C) experienced by M. norvegica
during its diel vertical migrations (gray shaded area) between the mixed surface layer (0 – 25 m) at night and its refuge depth during the day (average of 145 m). The average of the temperature envelope, weighted by the photoperiod shown in (c), is represented by the thick gray line. (b) idem for T. raschii whose daytime depth was 115 m in average. (c) Photoperiod defined as the nighttime (thick black line) or daytime (thick dashed black line) proportion over 24 h. (d) Food conditions for individuals of both species (µg C.L-1): phytoplankton biomass within the mixed surface layer (0 – 25 m) (thick black line), mesozooplankton biomass above 100 m (dashed-dotted line) and below 100 m (dashed line). ... 22 Figure I.3 Schematic of the krill moulting process implementation. N: no; Y: yes. ... 25 Figure I.4 Schematic of the krill reproduction implementation for M. norvegica. The only conceptual
difference for T. raschii is that one breeding cycle requires only one moulting cycle, instead of two. ... 31 Figure I.5 Metabolic balance (red/blue shaded area) between the assimilated fraction of the individual
ingestion rates (red lines) and the individual respiration rates (blue lines), both in mg C.h -1.ind-1, for all simulated individuals of M. norvegica during (a) five years and (b) the last year of simulation. Respiration rates are presented on a negative axis to represent the loss of biomass. Dark grey shaded area: positive energetic balance; light grey shaded area: negative energetic balance. Time-evolution of simulated M. norvegica individuals’ body mass in mg C (black lines) and body length in mm (grey lines) during (c) five years and (d) the last year of simulation and corresponding observations from individuals sampled in the Estuary and Gulf of St. Lawrence system. Black circles: observed body mass (mg C); grey crosses:
observed body length (mm); horizontal dashed black line: body mass target for the calibration procedure (average of all observations, see text); vertical dashed lines on the panel (c): January 1st. ... 37 Figure I.6 (a) Metabolic balance between (red/blue shaded area) the assimilated fraction of the
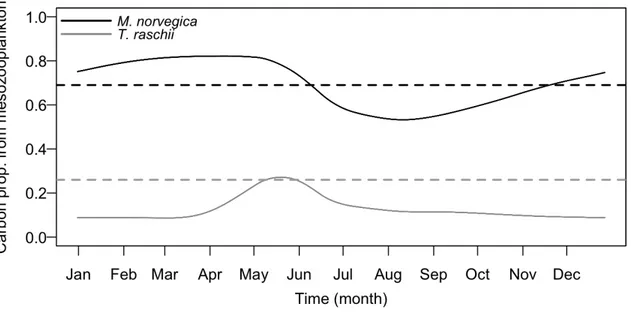
individual ingestion rates (red lines) and the individual respiration rates (blue lines), both in mg C.h-1.ind-1, for all simulated individuals of T. raschii during the last year of simulation. Dark grey shaded area: positive energetic balance; light grey shaded area: negative energetic balance. (b) Time-evolution of simulated T. raschii individuals’ body mass in mg C (black lines) and body length in mm (grey lines) and corresponding observations from individuals sampled in the Estuary and Gulf of St. Lawrence system. Black circles: observed body mass (mg C); grey crosses: observed body length (mm); horizontal dashed black line: body mass target for the calibration procedure (average of all observations, see text). ... 39 Figure I.7 Proportion of body carbon derived from mesozooplankton for simulated individuals of M.
norvegica (black) and T. raschii (grey) during the last year of simulation. Dashed lines: targets for the calibration procedure (average of observations). ... 40 Figure I.8 Comparisons between the simulated (lines) and observed (circles) individual condition
indices (ratio of mass to length) for (a) M. norvegica and (b) T. raschii, for the last year of simulation. Black dots and error bars: mean and standard error of the observations. ... 41 Figure I.9 Simulated patterns of (a) M. norvegica and (b) T. raschii individuals for the last year of
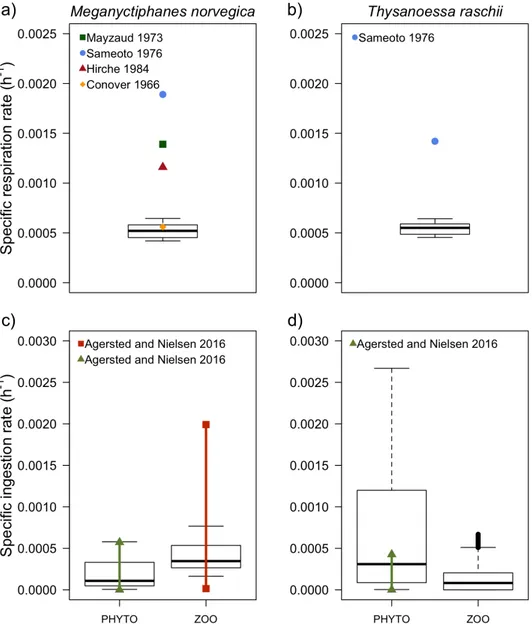
simulation. Red dots: number of eggs per clutch per female (left axis); black lines: gonad body mass proportion (right axis). ... 43 Figure I.10 Upper panels: distribution of simulated mass-specific respiration rates for (a) M.
norvegica and (b) T. raschii individuals compared to values reported in the literature (see legend). Each boxplot includes all the individual values simulated over the last year of simulation. Bottom panels: distribution of simulated mass-specific ingestion rates on phytoplankton and mesozooplankton for (c) M. norvegica and (d) T. raschii individuals compared to values reported in the literature when available (see legend). Again, includes all the values simulated over the last year of simulation. In (c), the thick red line between both squares represents the range of mass-specific ingestion rates observed for M. norvegica, based on a copepodite (late development stages of copepods) diet offered at a concentration of 494 μg C.L-1 and at a temperature of 5 °C (Agersted and Nielsen 2016). In (c) and (d), the green line between the triangles represents the range of mass-specific ingestion rates observed for each species based on a phytoplankton diet offered at concentrations varying between 75 and 225 µg C.L-1 and at 5 °C (Agersted and Nielsen 2016). ... 45 Figure I.11 Sensitivity analysis of the model’s parameters: A is the assimilation efficiency coefficient;
x0,phy andx0,zoo are the clearance rate scaling factor for phytoplankton and mesozooplankton,
respectively; ei is the activation energy for ingestion processes; h0 is the handling time
scaling factor; r0 is the respiration scaling factor; Q is the fraction of growth allocated to the
gonad. Upper panels: sensitivity of M. norvegica model. Mean relative change for the last year of simulation of (a) body mass and (b) egg production. Lower panels: sensitivity of T. raschii model. Mean relative change for the last year of simulation of (c) body mass and (d) egg production. Black bars: responses for a 10% decrease of the parameters relative to the base run; grey bars: responses for a 10% increase of the parameters. Dashed line: null horizon. ... 47
Figure II.1 Canadian east coast locating the Gulf of St. Lawrence (left side) and detail map of Gulf of St. Lawrence with areas of interest (right side). Isobaths 200 m (light blue line) are indicated. The orange line represents the extent of the North Shore (from Pointe-des-Monts in the south
to Blanc-Sablon in the north). ... 62
Figure II.2 Monthly mean forcing for the surface mixed layer (0-30 m) for temperature (a,b,c), phytoplankton biomass (d,e,f) (mg C.m-3) and mesozooplankton biomass (g,h,i) (mg C.m-3) in April (a,d,g), June (b,e,h) and August (c,f,i) in the EGSL. ... 68
Figure II.3 Positive daily growth potential in the percentage of the individual body mass for M. norvegica (upper panels) and T. raschii (bottom panels) during spring (a,d), early summer (b,e) and late summer (c,f) in the "Static" simulations. Positive growth potential ranged from 0 to 0.25% per day. ... 70
Figure II.4 (a) Distribution of the individual monthly daily growth potential expressed in % of body mass change for each simulated month and (b) proportion of GSL area where positive growth occurred for each month. M. norvegica is represented in purple and T. raschii is represented in green. Red dashed line represents the zero horizon. ... 71
Figure II.5 Upper panels: Relative frequency distribution of females according to the number of spawning events during the breeding season throughout the EGSL for M. norvegica (a) and T. raschii (b). Bottom panels: Relative frequency distribution of females producing eggs and the associated mean number of eggs released per female (black dots) according to the month of the year throughout the EGSL for M. norvegica (c) and T. raschii (d). Error bars on egg release are included but are very narrow and basically unnoticeable. ... 72
Figure II.6 Results of the clustering analysis for M. norvegica in the "Static" simulation. (a) Mapping of the physiological clusters. Individual clusters were grouped further according to the overall production patterns as described in the right-hand legend (see text). (b) Distribution of the individual values of cumulative growth (mg C) according to the physiological clusters; dashed line represents the zero horizon. (c) Distribution of the individual values of cumulative egg production according to the physiological clusters. (d) Median mass trajectory (mg C) of each physiological cluster along the period simulated; dashed line represents the initial body mass. ... 74
Figure II.7 Results of the clustering analysis for T. raschii. Same description as Fig. II.6. ... 75
Figure II.8 Results of the clustering analysis for M. norvegica for the month of April in the "Transport" simulation. (a) Mapping of the physiological clusters determined by the analyses. Individual clusters were grouped further according to the overall growth and reproduction patterns as described in the right-hand legend (see text). (b) Frequency distribution of the cumulative mass of all individuals. The dashed line represents the zero threshold. The red line represents the 90th percentile of the distribution, while each individual black dot corresponds to the median value of each cluster; (c) Median mass trajectory (mg C) of each physiological cluster for the month. Dashed line represent the initial body mass. ... 78
Figure II.9 M. norvegica. Same as Fig. II.8 for the month of June. ... 79
Figure II.10 M. norvegica. Same as Fig. II.8 for the month of August. ... 80
Figure II.12 T. raschii. Same as Fig. II.8 for the month of June. ... 82
Figure II.13 T. raschii. Same as Fig. II.8 for the month of August. ... 83
Figure II.14 Synthesis map representing the best physiological trajectories in the EGSL during the productive season (April to August) for Thysanoessa raschii (a) and Meganyctiphanes norvegica (b). Each colour represents monthly trajectories of the physiological clusters that showed positive growth and egg production. Black dots represent dense krill aggregations (biomass > 100 g.m-2) according to acoustics surveys done in August 2005 and 2007 with a multibeam Simrad EK500 echosounder (see text). ... 84
Figure III.1 Map of Estuary and Gulf of St. Lawrence system with areas of interest indicated. 300 m (light-blue line) and 100 m isobaths (dark blue dashed line) shown. ... 98
Figure III.2 Distribution of all parameters used in "paramosome" for the "Individuals" simulation. The red dashed lines represent the mean values used for the "Clones" simulations. ... 103
Figure III.3 Monthly mean conditions in the surface mixed layer (0-30 m) for temperature (a,b), phytoplankton biomass (d,e) (mg C.m-3) and mesozooplankton biomass (g,h) (mg C.m-3) in 2003 (a,d,g) and 2006 (b,e,h) and the anomalies between both years for each forcings (c,f,i) in the EGSL. ... 107
Figure III.4 Comparison of the frequency distributions of individual body masses at the end of the "Clones" and "Individuals" simulations in a) June 2003 and b) June 2006.The same results are compared by year for c) the "Clones" simulations and d) the "Individuals" simulations. ... 110
Figure III.5 Comparison of the frequency distributions of individual cumulative egg production for the "Clones" and "Individuals" simulations in a) June 2003 and b) June 2006. The same results are compared by year for c) the "Clones" simulations and d) the "Individuals" simulations. ... 111
Figure III.6 Results of the clustering analysis for the year 2003 in the "Clones" simulation. (a) Mapping of the physiological clusters determined by the analyses. Individual clusters were grouped further according to the overall production patterns as described in the right-hand legend (see text). (b) Frequency distribution of the cumulative mass of all individuals. The dashed line represents the zero threshold. The red line represents the 90th percentile of the distribution, while each individual colour dot corresponds to the median value of each numbered cluster; (c) Median mass trajectory (mg C) of each physiological cluster for the month. ... 113
Figure III.7 Same as Fig. III.6 for the year 2006. ... 114
Figure III.8 Same as Fig. III.6 for the "Individuals" simulation. ... 115
Figure III.9 Same as Fig. III.7 for the "Individuals" simulation. ... 116 Figure III.10 Results of the Canonical Correlation Analyses (CCorA) for a) the first canonical axes
for 2003 and c) 2006 and for b) the second canonical axes for 2003 and d) 2006. In each panel, the upper barplot shows the loadings of the biological response variables on their respective axis, whereas the lower barplot shows the cross-loadings of the parameters on the
biological variable axis (represents how each parameters’ values influence the biological variables axis). ... 123
Liste des tableaux
Table I-1 Parameters used in the model. The square brackets report the range of values from the literature. ... 32 Table II-1 Parameters used in the model. The square brackets report the range of values from the
literature. ... 64 Table III-1 Parameters used in the model. The square brackets report the range of values used in
"Individuals" simulations ... 104 Table III-2 Mean and coefficient of variation (CV) of individual final mass during June of 2003 and
2006 for both "Clones" and "Individuals" simulations. ... 110 Table III-3 Mean and coefficient of variation (CV) of individual cumulative egg production during
June of 2003 and 2006 for both "Clones" and "Individuals" simulations; computed with egg-producing individuals only. ... 111
Liste des sigles
0-D 3-D AGC CCorA CDOM CI CIL CRSNG CTD DFO dt DVM EGSL FA GAM GAMM GMC-GEM GSL IBM IMP LSLE LHS MPO neGSL NEMO NEMO-OPA NF NPZD nwGSL SLE SST Zero-Dimensional Three-Dimensional Atlantic Goescience CenterCanonical Correspondance Analysis Coloured Dissolved Organic Matter Conditional Indice
Cold Intermediate Layer
Conseil de Recherches en Sciences et en Génie du Canada Conductivity Temperature Depth
Department of Fisheries and Oceans Timestep
Diel Vertical Migration
Estuary and Gulf of St. Lawrence Fatty Acids
General Additive Model General Additive Mixte Model
Canadian Meteorological Center-Global Environmental Multiscale Gulf of St. Lawrence
Individual Based Model Inter-Moult Period
Lower St. Lawrence Estuary Latin Hypercube Sampling Ministère des Pêches et Océans Northeast Gulf of St. Lawrence
Nucleus for European Modeling of the Ocean
Nucleus for European Modeling of the Ocean – Ocean Parallisé configuration NewFoundland
Nutrient-Phytoplankton-Zooplankton-Detritus northwest Gulf of St. Lawrence
St. Lawrence Estuary Sea Surface Temperature
Perseverare – « La persévérance, c'est ce qui rend l'impossible possible, le possible
probable et le probable réalisé »
Léon Trotsky
Amare – « Aimer c'est savoir dire je t'aime sans parler »
Victor Hugo
Talis ipse esse – « Etre différent n'est ni une bonne ni une mauvaise chose. Cela
signifie simplement que vous êtes suffisamment courageux pour être vous même » Albert Camus
Remerciements
Lorsque j’ai commencé mon doctorat un grand nombre de personnes m’a dit que ce serait une véritable épreuve. Mais tant que l'on ne vit pas cette expérience, il est difficile de mesurer cette réalité. Ces quatre années de thèse ont été pour moi une expérience enrichissante dans laquelle j'ai su m'épanouir et me découvrir, bien que tous les jours ne fussent pas faciles. C'est grâce à mon entourage extraordinaire que j'ai eu la chance d'avoir, que j'ai pu surmonter les difficultés et relever les défis tout au long de ma thèse et aujourd'hui je remercie toutes ces personnes.
Je remercie en premier lieu, mon directeur de doctorat Frédéric Maps qui m’a avant tout donné l’opportunité d’entreprendre cette belle aventure et qui a su m'accorder sa confiance pour mener à bien ce projet de recherche. Il a été un guide lors mon apprentissage et m’a soutenue tout au long de mon projet, m’encourageant et m’apprenant ce que je ne connaissais pas encore. Merci aussi pour ta patience Fred dans mes moments de doutes, de panique et de crise existentielle. Merci, car sans toi cela n'aurait pas été possible.
Je remercie mes codirecteurs, Stéphane Plourde et Diane Lavoie, pour leurs conseils et leurs expériences. Ils m'ont apporté un point de vue externe et une mise en perspective de mon projet qui n'a été que bénéfique à ma formation et à ma recherche.
Je remercie les autres membres de mon jury d'évaluation et de mon jury de thèse, pour leur temps consacré à la relecture et aux corrections de mes travaux de recherche, ainsi que pour leurs conseils.
Je remercie tous les membres de mon laboratoire le NEOLAB (Karolane, Christiane, Blanche, Inge, Maxime et Arnaud) avec qui j’ai eu la possibilité d’évoluer, d’apprendre et d’échanger et avec qui nous avons construit ce jeune laboratoire de modélisation à l’Université Laval. Au sein de ces membres, je remercie tout particulièrement Christiane Dufresne qui a été une véritable amie et un modèle pour moi et qui a su m’épauler et me conseiller tout au long de cette thèse.
Un grand merci à Vincent Martel pour son aide et son apprentissage lors de l'adaptation de mon modèle dans ce bon vieux langage archaïque FORTRAN 90. Il lui aura
fallu beaucoup de la patience et d'énergie pour gérer mes incompréhensions parfois tenaces face à la programmation.
Je remercie aussi David Beauschene et Philippe Massicotte pour leur soutien technique avec le logiciel R et lors de mes moments de désespoirs face à mes bugs informatiques.
Merci au corps enseignant, aux étudiants gradués et non gradués, aux professionnels de recherche, à l'équipage du navire de recherche Amundsen 2016, que j'ai côtoyé et avec qui j'ai eu l'occasion de travailler. Un merci à Louis Fortier qui m'a permis d'intégrer son équipe et de partir en mission en Arctique à bord de l'Amundsen. Et merci à Cyril Aubry et Sarah pour leur formation sur l'échantillonnage en mer et cette expérience inoubliable que nous avons vécus sur le bateau.
Je remercie également tous mes amis autant au Canada que ceux de l’autre côté de l’Atlantique en France, qui ont su supporter mes railleries incessantes, mes sauts d’humeurs et mes excès d’énergie (surtout les deux premières années) et qui n’ont jamais arrêté d’être présent. Merci à ma p’tite Marie d'avoir su vivre avec moi et me supporter 24h/24h pendant ma dernière année de thèse, qui ne fut pas la plus calme et douce. À toi Fanny, pour ta douceur et ta gentillesse constante. À vous Pauline et Théo, pour ces soirées passionnantes et nos jeux et paris débiles de dés et de cartes. À toi Steph, pour ces moments d'évasion et de calme au Saguenay. Et à toi Rémi pour ces superbes moments passés ensemble.
Un inconditionnel merci à ma famille, mes parents et mes frères, ainsi qu’à ma meilleure amie Cynthia qui n’ont jamais douté de moi et qui m’ont donné tout l’amour qu’ils pouvaient pour que ma vie loin d’eux se passe bien, que la distance me paraisse moins difficile à aborder et que les tumultes émotionnels de ma thèse soient plus faciles à accepter. Je remercie aussi mon petit chat Pepsie, membre à part entière de ma famille, qui m’a suivi pendant 19 longues années, subissant d’innombrables voyages et qui a su me donner toute l’affection dont elle était capable jusqu’à son dernier jour en mai dernier.
Je commence maintenant une nouvelle aventure et espère rencontrer autant de merveilleuses personnes que ce qui fut le cas durant mon doctorat. Merci à tous, car je ressors de cette aventure grandie, satisfaite et heureuse.
Avant-propos
Cette thèse de doctorat comprend une introduction générale, trois articles scientifiques, correspondant aux Chapitres 1, 2 et 3, et une conclusion générale.
Le chapitre 1 a été publié le 14 septembre 2018 dans la revue Limnology and Oceanography. Il s'intitule "Individual-based modelling explains the contrasted seasonality in size, growth and reproduction of the sympatric Arctic (Thysanoessa raschii) and Nordic krill (Meganyctiphanes norvegica) in the St. Lawrence estuary, eastern Canada". Cet article est passé au travers d'un processus de révision suite à une première soumission le 3 mars 2018, et suite à des modifications suggérées celui-ci a été retravaillé et soumis une seconde fois le 26 juin 2018, pour finalement être accepté à la publication le 30 juillet 2018. La version intégrée dans le premier chapitre de cette thèse est identique à la version publiée dans le journal. En tant que première auteur, ma contribution à ce travail fut l'essentiel de la recherche bibliographique, la conception du modèle, le lancement des simulations numériques, le traitement des résultats et la rédaction de l'article. Le professeur Frédéric Maps a contribué au design du modèle et au lancement des simulations numériques. Le Dr. Stéphane Plourde a aidé au design du modèle. La Dr. Gesche Winkler, Dr. Jory Cabrol, Angélique Ollier et Laurie-Emma Cope ont mené à bien les échantillonnages de terrain et les expériences de laboratoire fournissant les données de validation et de calibration du modèle. Finalement tous les coauteurs ont contribué à la rédaction du manuscrit et au processus de révision de manière significative.
Le chapitre 2 a été soumis le 12 décembre 2018 dans le revue Progress in Oceanography pour l'édition spéciale Climate, Zooplankton and Salmon. Cet article est passé au travers d'un processus de révision. Des modifications suggérées, ont été apportées et une nouvelle version a été resoumise le 25 mai 2019.
Le chapitre 3, est en préparation à la publication.
Les résultats de cette thèse ont été présentés à plusieurs reprises, sous la forme de présentations orales et écrites dans les conférences nationales et internationales suivantes :
[1] AGA Québec-Océan, Modèle individuel physiologique pour le krill, Thysanoessa
raschii et Meganyctiphanes norvegica, dans l'estuaire et le golfe du Saint- Laurent, Nov.
8-9, 2016, Rimouski, Canada (Communication orale) * Mention de mérite obtenue
[2] ArcticNet ASM, Understanding the complex relationship between the environmental variability and the physiology of the arctic krill Thysanoessa raschii and the northern krill Meganyctiphanes norvegica, Dec. 5-9, 2016, Winnipeg, Canada (Communication orale)
[3] ArcticNet ASM, Understanding the complex relationship between the environmental variability and the physiology of the Arctic krill Thysanoessa raschii and the northern krill Meganyctiphanes norvegica, Dec. 5-9, 2016, Winnipeg, Canada (Poster)
[4] 8ème édition du Colloque de biologie de l'université Laval, Modèle physiologique individu-centré du krill (Thysanoessa raschii et Meganyctiphanes norvegica) dans l'estuaire et le golfe du Saint- Laurent, Mar. 29-30, 2017, Québec, Canada (Communication orale)
[5] 3rd International Symposium on Krill, Transiting through a variable seascape: impacts on two sympatric krill physiology productivity in the sub-Arctic Gulf of St. Lawrence, Canada, June 12-16, 2017, St. Andrews, UK (Communication orale)
[6] ArcticNet ASM, Productive areas of the Arctic krill (Thysanoessa raschii) and the Nordic krill (Meganyctiphanes norvegica) in the subarctic Gulf of St. Lawrence (Canada) revealed by their physiology and swimming behaviour, Dec. 10-15, 2017, Québec, Canada (Poster)
[7] World Conference on Marine Biodyversity (accepted Abstract), Modelled emergent productivity areas on dominant krill species depending on their physiology and swimming behaviour in Gulf of St. Lawrence, May 13-16, 2018, Montréal, Canada (Communication orale)
Les travaux de cette thèse ont également été mis en lumière par des confrères lors de leur invitation à des conférences à l'international :
[8] Savenkoff C, Lavoie D, Cabrol J, Ollier A, Guilpin M, Benkort D, de Maillard B. Krill, key component of the Estuary and Gulf of St. Lawrence, Canada: from the individual to the ecosystem. June 15, 2016, Université de Toulouse, France
[9] Winkler G, Plourde S, Tremblay T, Nozais C, Maps F, Lesage V, McQuinn I, Starr M, Chabot D, Savenkoff C, Lavoie D, Cabrol J, Ollier A, Guilpin M, Benkort D, de Maillard B. Midway between phytoplankton and whales: Krill of the Estuary and Gulf of St. Lawrence, Canada. November 4, 2016, University of Hamburg, Germany
Introduction générale
Les euphausiacés
Les euphausiacés, plus communément connus sous le nom de krill que leur ont attribué les baleiniers norvégiens, sont des crustacés planctoniques. Le plancton est un groupe hétérogène d’organismes aquatiques dont les individus sont fondamentalement dépendants du mouvement des masses d'eau sur le plan horizontal (Balvay 2006). Le krill est essentiellement composé d’espèces zooplanctoniques dont les stades de développement juvéniles et adultes sont de taille relativement grande, quelques centimètres, ce qui en fait donc une constituante importante du macrozooplancton. Le macrozooplancton se retrouve à la limite entre le plancton (passif) et le necton, constitué d’organismes pouvant effectuer des mouvements autonomes par rapport aux courants de leur milieu. Les individus de krill peuvent s'affranchir facilement des courants verticaux et entreprendre des migrations journalières (nycthémérales) de plusieurs centaines de mètres d'amplitude en quelques heures, entre la sécurité relative de l’obscurité des profondeurs le jour et l’abondance de nourriture à la surface la nuit (Mauchline and Fisher 1969; Kaartvedt 2010). Ces migrations constituent d’ailleurs le plus important déplacement de biomasse sur notre planète. Les vitesses de nage des plus grands individus peuvent même excéder les courants horizontaux les plus faibles (entre 0.03 et 0.15 m.s-1 ; Kils 1982; Hofmann et al. 1998) grâce à leurs appendices natatoires très efficaces (Thomasson et al. 2003) et ainsi maintenir des formations en bancs cohésifs.
On retrouve le krill distribué à travers tous les océans du monde, réparti en 85 espèces connues (Everson 2000). Ces espèces peuvent être herbivores ou omnivores, ce qui fait de ce groupe une plateforme de transfert trophique importante vers de nombreux prédateurs pélagiques (Levasseur 1996; Bamstedt and Karlson 1998) puisqu’il assure le lien trophique entre les producteurs primaires, le zooplancton unicellulaire et métazoaire et les producteurs secondaires supérieurs (Everson 2000). Les euphausiacés sont après les copépodes le groupe de crustacés zooplanctoniques le plus important dans les écosystèmes pélagiques et vivent en bancs pouvant atteindre des tailles et des biomasses considérables (Townsend 2012).
Ces grandes agrégations que forment les individus de krill, leur confèrent une place importante d'espèces fourragères, c'est-à-dire qu'il constitue une source importante de nourriture pour un grand nombre d'espèces. Il joue ainsi un rôle écologique important, mais également un rôle dans l'économie liée aux ressources marines renouvelables dans de nombreuses régions côtières en raison de son rôle clé d'espèce fourragère pour plusieurs espèces d'importance économique telles que les poissons et invertébrés épibenthiques (morue, sole, plie, etc.) (Sameoto et al. 1994; Simard and Harvey 2010) et pélagiques (harengs, maquereau, crevettes, etc.). À noter que le krill exerce un rôle tout aussi important pour des espèces non exploitées, mais qui génèrent une activité économique indirecte d'écotourisme et d'observation en milieu naturel, telles que les espèces menacées de grands rorquals, en particulier dans l’Océan Austral (Nicol and Endo 1997; Sourisseau et al. 2006; Simard and Harvey 2010).
Contexte de l'étude
Depuis la fin des années 50, on observe à travers la planète le développement d'une pêcherie ciblant directement le krill, en tant que ressource pour l'alimentation animale et l'aquaculture (Nicol and Endo 1997; Martin 2007). Ces pêcheries ont atteint un pic dans le courant des années 1980 avec un ralentissement durant les années 1990 suite à l’effondrement de l’Union Soviétique (Kawaguchi and Nicol 2007). Cependant, depuis les années 2000, on note un regain d'intérêt pour le krill soutenu par les industries pharmaceutiques, cosmétiques et nutraceutiques en raison du profil de composition lipidique du krill composé d’environ 40% de phospholipides et de 30% d’acides gras oméga-3 (Devaux 2012). Cette composition particulière présente un fort potentiel commercial pour l'utilisation en tant que complément alimentaire et matière première pour les composés biochimiques à forte valeur ajoutée. Les espèces exploitées actuellement par ces pêcheries sont principalement Euphausia superba en Antarctique et Euphausia pacifica au large des côtes japonaises. Cependant dans l’estuaire et le golfe du Saint-Laurent (EGSL), l'augmentation relativement récente des études sur le krill et l'accumulation de nouvelles données de biomasse obtenues in situ par de nouvelles technologies acoustiques ont ranimé un certain intérêt du secteur privé pour une potentielle pêche commerciale. L'obtention de meilleures connaissances biologiques et écologiques sur
les espèces dominantes de cette zone géographique devient d'autant plus nécessaire dans ce contexte, car avant toute éventuelle exploitation d'espèces clés aussi importantes, il est primordial de détenir les connaissances suffisantes pour la mise en place d'une gestion écosystémique éclairée.
De plus, les changements climatiques affectent significativement l'environnement en termes de température, de salinité, de courants, d’oxygène et d'acidité qui peuvent engendrer des changements fondamentaux dans la biologie, l'écologie et la structure des espèces et des écosystèmes (Hoegh-Guldberg and Bruno 2010). Dans un tel contexte, comprendre l'impact de la variabilité naturelle de l'environnement physique sur la physiologie et la dynamique des populations de krill apparaît essentiel pour appréhender la dynamique et l'évolution des populations de krill de l’EGSL, afin de mettre en place des plans de gestion durable.
Site de l’étude : l'estuaire et le golfe du Saint-Laurent
L'EGSL situé sur la côte est du Canada représente un des plus grands écosystèmes estuariens de la planète (Fig. 0.1). En raison de sa position en marge occidentale du bassin de l’Atlantique Nord et de son ouverture au Nord sur le Courant Côtier du Labrador, il représente la mer la plus au Sud présentant des conditions environnementales subarctiques, caractérisées par un couvert de glace saisonnier présent plusieurs mois de l'année. L’EGSL présente une forte variabilité saisonnière et interannuelle de ses conditions océanographiques et climatiques, et un environnement marin particulier où se rencontrent des masses d’eau d’origine Arctique et Atlantique, avec leurs assemblages respectifs d’espèces planctoniques associées.
Figure 0.1 Carte du domaine d'étude, le golfe et l'estuaire du Saint-Laurent, montrant l'isobathe 200 m (ligne bleue), les principaux chenaux (Laurentien, Esquiman) et détroit (Cabot, Belle Isle, Jacques-Cartier). La croix cerclée rouge représente la station Rimouski et la ligne orange représente la délimitation de la Côte-Nord (de Pointe-des-Monts au sud à Blanc-Sabon au nord).
L’EGSL est une mer semi-fermée d’une superficie de 240 000 km² (Benoît et al. 2012). Il est connecté à l'Océan Atlantique au sud via le large et profond détroit de Cabot et à la Mer du Labrador au Nord par l’étroit et peu profond détroit de Belle Isle (Saucier et al. 2003). La circulation générale dans l’EGSL est cyclonique, mais avec une forte empreinte saisonnière. L’EGSL est caractérisé par de forts gradients horizontaux et verticaux des propriétés des masses d’eau et des courants (Lavoie et al. 1996; Saucier et al. 2009). La circulation est principalement influencée par l’apport d’eau douce des rivières et du fleuve Saint-Laurent ainsi que par les marées et les forçages atmosphériques (Saucier et al. 2003; Saucier et al. 2009; Lavoie et al. 2016). Il est alimenté en eau douce par plusieurs tributaires dont les principaux sont bien sûr le fleuve Saint-Laurent, mais aussi les rivières Saguenay, Manicouagan, aux Outardes, du Petit Mécatina et Natashquan, toutes situées sur sa rive Nord (Centre-Saint-Laurent 1996).
Différents régimes de circulation se mettent en place au fil des saisons dans l'EGSL. En été une circulation estuarienne s’organise en trois couches distinctes. Avec l'augmentation de la température de surface et l'augmentation de l'apport d'eau douce suite à la fonte glaciaire, on observe une stratification de la couche d'eau froide qui se divise alors en deux couches. Le système s'organise alors de la manière suivante : une couche profonde salée d’origine Atlantique, une couche intermédiaire froide (CIL) et une mince couche mélangée de surface moins salée et plus chaude dont les températures peuvent atteindre jusqu’à une vingtaine de degré au sud du golfe à la fin de l’été. La CIL s'étend de 10 à 30 m sous la surface jusqu'à environ 100 à 150 de profondeur et est un trait particulier au golfe du Saint-Laurent (Berkes 1973; Gilbert and Pettigrew 1997; Saucier et al. 2003; Cotté and Simard 2005). En hiver, la couche de surface qui a été érodée par le refroidissement et l’action des tempêtes automnales qui l’ont mélangée à la CIL pendant l’automne disparait et on retrouve alors un système à deux couches avec une CIL qui s'étend jusqu'à la surface au-dessus de la couche profonde salée. Ces différentes masses d'eau représentent des habitats préférentiels pour les espèces arctiques et boréales présentes dans ce système unique.
Le krill dans l'estuaire et le golfe du Saint-Laurent
On retrouve dans l’EGSL une communauté de krill composée de quatre espèces (Levasseur 1996) : Meganyctyphanes norvegica (M. Sars, 1857), Thysanoessa raschii (M. Sars, 1864), T. inermis (Kroyer, 1846) et T. longicaudata (Kroyer, 1846). Parmi ces quatre espèces, deux sont dominantes en termes d'abondance, de biomasse et de répartition spatiale et sont donc les espèces cibles de cette étude : le krill arctique T. raschii et le krill nordique
M. norvegica (Fig. 0.2) (Berkes 1973; McQuinn et al. 2015). Ce sont des espèces
sympatriques, c'est-à-dire des espèces possédant des aires géographiques qui se recoupent, partiellement ou entièrement, au moins durant la saison de croissance et de reproduction. Ces espèces se retrouvent en sympatrie dans l’EGSL en raison du régime de circulation océanographique particulier décrit ci-dessus, à savoir le mélange des eaux d'origine arctique du courant côtier du Labrador entrant par le détroit de Belle Isle au nord et de masses d'eaux de l'Atlantique Nord pénétrant par le détroit de Cabot au sud.
a) b)
Figure 0.2 Meganyctiphanes norvegica adulte (a) et Thysanoessa raschii adulte (b). Les dessins représentatifs sont tirés de Marine Species Identification Portal ( http://species-identification.org/index.php).
Ces deux espèces jouent un rôle écologique similaire d'espèce fourragère au sein du réseau trophique (Fig. 0.3), bien qu'elles présentent des différences biologiques et physiologiques notables (Berkes 1973; Sameoto 1976; Agersted and Nielsen 2016). En premier lieu, leur enveloppe de préférence thermique les distingue : M. norvegica est une espèce eurytherme dont les populations se retrouvent de la Méditerranée au sud à la Mer de Barents au nord. Cette espèce évolue préférentiellement dans la couche Atlantique profonde du système de masses d'eau de l'EGSL où elle se réfugie le jour lors de ses migrations nycthémérales. T. raschii quant à elle, est une espèce sténotherme présente essentiellement dans les eaux de l'Océan Arctique et de ses mers ancillaires et évolue plutôt dans la CIL de l'EGSL durant le jour (Berkes 1976; Sameoto 1976; Harvey et al. 2009). Ces deux espèces présentent également d'autres différences importantes en termes notamment de reproduction et d’alimentation. On observe des différences pour les saisons de reproduction, au printemps pour T. raschii et à l’été pour M. norvegica (Berkes 1973; Ross and Quetin 2000), ainsi que pour les modes de reproduction, M. norvegica montrant des individus se synchronisant les uns aux autres durant les périodes de ponte (Tarling and Cuzin‐Roudy 2003). Bien que les deux espèces soient généralement qualifiées d’omnivores, la diète de M. norvegica comprend une proportion plus importante de crustacés planctoniques d’ordre de grandeur de quelques mm pour les plus gros, tel que le copépode Calanus finmarchicus très abondant dans l’EGSL et dont les stades de développement avancés atteignent 2 à 3 mm) (Macdonald 1927 ; Guittard and Joly 1999). Cette espèce semble physiologiquement adaptée à ce type d’alimentation,
alors que T. raschii semble privilégier une alimentation à base d’espèces phytoplanctoniques (Macdonald 1927 ; Berkes 1973 ; McClatchie 1985).
Figure 0.3 Représentation schématique du réseau trophique de l’estuaire et du golfe du Saint-Laurent centré sur le krill (adapté de Harvey et al. 2003).
La modélisation comme outil d'exploration
En océanographie, la modélisation numérique est très présente tant au niveau physique (formation et circulation des masses d'eau, couplage atmosphère-océan, etc.) qu'au niveau biologique (croissance et productivité des espèces, dynamique des populations, distribution des populations, etc.) et elle est devenue un outil pluridisciplinaire de choix pour la gestion intégrée des écosystèmes marins. Comme l'a écrit le statisticien Georges Box (Box 1979) « Tous les modèles sont faux, mais certains sont utiles ! ». Un modèle ne permet pas de représenter l'ensemble du système étudié, mais est un outil qui permet par une représentation simplifiée de se focaliser sur un aspect particulier de ce système (Coquillard and Hill 1997). Cependant, l'utilisation de modèles numériques rend possible l'association de processus complexes souvent étudiés séparément lors des expériences in situ et de laboratoire, et ils permettent aussi l'étude des mécanismes à différentes échelles spatio-temporelles et une meilleure compréhension des interactions entre les processus biologiques et leur environnement. Ils permettent de mettre en évidence les connaissances manquantes sur les mécanismes étudiés, de faire émerger des propriétés du système qui ne sont pas identifiables par les systèmes d'échantillonnage actuels et servent à produire de nouvelles pistes de recherches en testant et générant de nouvelles hypothèses falsifiables. C'est donc à l'aide de modèles numériques que les travaux de recherche présentés dans cette thèse ont été abordés, car la modélisation représente l'outil idéal pour comprendre la réponse de la dynamique des populations de krill dans l’EGSL face à la variabilité environnementale.
Objectifs du projet de Doctorat
Ce projet de doctorat s'inscrit au sein d'un Projet Stratégique financé par le CRSNG intitulé "Production et consommation du krill : vers l’évaluation écosystémique de l’état des stocks" dont l'objectif principal est d'évaluer et de comprendre la résilience écologique des populations de M. norvegica et de T. raschii dans l’EGSL, alors qu'aucune exploitation humaine n'en a encore été effectuée. L'objectif principal de ce doctorat est d'étudier et
de la variabilité environnementale sur la distribution, la croissance et la reproduction de ces deux espèces.
Comme toutes les espèces de zooplancton, la distribution et la dynamique spatiale du krill sont principalement contrôlées par les courants et le déplacement des masses d'eau dans lesquelles il vit. Toutefois, dès les stades larvaires, les individus développent des capacités natatoires impressionnantes et ils effectuent notamment des migrations verticales quotidiennes de plusieurs centaines de mètres pour s'alimenter en surface et se protéger des prédateurs en profondeur. Le résultat final des interactions entre les déplacements du krill et les mécanismes de transport dans l’EGSL contribuent de façon importante aux patrons spatiotemporels de répartition des populations de krill observées sur le terrain (Maps et al. 2015 ; Lavoie et al. 2016). Pour une compréhension plus complète et approfondie de la dynamique de production et de transport de ces espèces fourragères clés, il est aussi nécessaire de bien comprendre comment la variabilité de l’environnement à des échelles spatiotemporelles différentes agit sur leur physiologie. En effet, la biologie et la physiologie des individus jouent un rôle important sur deux plans. Premièrement, concernant les propriétés essentielles de dynamique des populations comme le développement et la phénologie d’une part, et le potentiel de croissance et de reproduction d’une autre part. Deuxièmement, l’état physiologique des individus, leur niveau de développement, leur niveau de satiété ou bien leur sexe influencent les comportements de nage (migrations nycthémérales ou comportement grégaire) (Pearre 2003 ; Tarling 2003 ; Sourisseau et al. 2008 ; Piñones et al. 2013). Cet état physiologique intervient donc dans la dynamique spatiale des agrégations et des populations de krill. Le premier chapitre de cette thèse a donc pour objectif de développer un modèle physiologique basé sur l'individu (IBM) pour les
adultes de chaque espèce et d’évaluer les impacts de la variabilité environnementale à une échelle annuelle sur leur potentiel de production dans le système de l’EGSL. Un
modèle bioénergétique de croissance et de reproduction (production d’œufs) basé sur les principes premiers de la biologie tels que la relation à la température d’Arrhenius, la relation allométrique à la masse corporelle et une réponse fonctionnelle de type Holling III a été élaboré pour les deux espèces dominantes de krill de l'EGSL. Le modèle a été confronté aux conditions environnementales retrouvées à la station d'échantillonnage Rimouski de l'estuaire maritime du fleuve Saint-Laurent (Fig. 0.1).
Comme cela a été décrit précédemment, l'EGSL est un environnement complexe dont la variabilité spatiale et temporelle est importante. Les conditions environnementales rencontrées par les espèces planctoniques varient donc selon des dynamiques spatiotemporelles très diverses et les réponses physiologiques des individus seront très contrastées selon l'espace et dans le temps. La dynamique de circulation exerce un contrôle très important sur les réponses des organismes, car les mécanismes de transport des populations zooplanctoniques sont des processus complexes intégrant à la fois des composantes environnementales (courants, topographie, forçages atmosphériques, etc.), physiologiques et comportementales propres à chaque individu (état physiologique, sexe, stade de développement, comportement grégaire, etc.). C’est cependant grâce aux interactions entre ces différentes composantes qu’il est possible d’avoir une vision et une compréhension complète de la dynamique spatiotemporelle de chaque espèce de krill. Les deux espèces M. norvegica et T. raschii montrent des périodes de reproduction différentes, essentiellement au printemps (avril à juin) pour T. raschii et en été (juin à septembre) pour
M. norvegica dans l'estuaire du Saint-Laurent (Berkes 1973, Ross and Quetin 2000, Plourde
et al. 2011) et par conséquent un développement démographique décalé dans le temps. Ainsi, l'influence de la circulation océanique sur les différents stades de développement est probablement différente entre les deux espèces, avec des conséquences encore méconnues sur la répartition et la différentiation des agrégations de M. norvegica et T. raschii observées dans le système. La différence interspécifique dans la période de reproduction illustre une des failles dans l’état actuel de la compréhension des mécanismes biophysiques responsables de la production des patrons spatiotemporels de distribution des populations de krill dans l’EGSL. D’une façon générale, les divers processus physiologiques et démographiques qui sont principalement influencés par la température et la disponibilité en nourriture ne présenteront pas le même fonctionnement pour des espèces différentes. Ainsi, avec des régimes alimentaires differents et des échelles de tolérance thermique différentes, les processus physiologiques de T. raschii et M. norvegica auront une influence différente sur la dynamique de transport de ces espèces.
D’un point de vue quantitatif, une meilleure compréhension des interactions entre les composantes environnementales, physiologiques et comportementales va également permettre d’estimer plus précisément les biomasses et les productions des populations de chacune des deux espèces de krill ainsi que leur évolution dans le temps. Les biomasses de
T. raschii et M. norvegica ont récemment été révisées à la hausse dans l’EGSL grâce à des
moyens d’observation acoustiques plus précis et des algorithmes de traitement innovants (McQuinn et al. 2015). La variabilité temporelle est accompagnée d’une variabilité spatiale. En effet, les populations de T. raschii entrent dans le système de l’EGSL principalement par le détroit de Belle Isle au Nord alors que M. norvegica emprunte le détroit de Cabot au Sud. Toutefois, la zone de Gaspé et le plateau madelinien constituent les zones de plus forte abondance d'œufs et de larves pour les deux espèces (Berkes 1973 ; Plourde et al. 2011). Le deuxième chapitre de cette thèse a donc pour objectif de coupler le modèle physiologique
développé dans le premier chapitre à un modèle de circulation régional, dans le but d’estimer les patrons spatiotemporels et le potentiel de production de M. norvegica et
T. raschii sur l'ensemble de l’EGSL. En associant les modèles de transport et de
physiologie, il sera possible de fournir un estimé quantitatif et les patrons spatiotemporels de la production de chaque espèce de krill dans l’EGSL et de fournir des éléments de réponse concernant la part relative de la production locale à l’échelle régionale par rapport à l’effet mécanique d'agrégation par les structures de courants. Ces estimations plus précises devraient aider à répondre à d’autres importantes questions écologiques telles que les cycles interannuels d’abondance d’œufs de krill dans l’estuaire (Plourde et al. 2011).
L'estuaire et le golfe du Saint-Laurent, en plus de montrer une variabilité saisonnière très contrastée, présentent qui plus est une forte variabilité interannuelle. Certaines années, comme 2003, sont caractérisées comme froides, et présentent des particularités comme un couvert de glace plus important, un retrait de cette glace plus tardif dans la saison, un refroidissement des eaux de surface, etc. (Galbraith et al. 2012 ; Galbraith et al. 2016). À l'inverse des années comme 2006, sont caractérisées comme chaudes, et présentent des particularités opposées au niveau des conditions océanographiques de glace et de température. Ces différences dans les conditions physiques (température et courant) vont affecter les espèces et communautés biologiques qui dépendant à la fois de la circulation des masses d'eau et de leurs caractéristiques physico-chimiques (Thorpe et al. 2004 ; Batchelder et al. 2013). Comme à l'échelle saisonnière, la variabilité environnementale interannuelle du golfe du Saint-Laurent va donc affecter la dynamique de transport et de production du krill. Cependant, il est fascinant de constater que les populations de krill qui prospèrent dans ce système démontrent elles aussi de larges fluctuations de leur biomasse et de leurs patrons spatiaux de distributions d’une année à l’autre. Depuis plusieurs années, une emphase est
faite sur le rôle de la variabilité interindividuelle dans la réponse d'une espèce face à son environnement (Bolnick et al. 2011; Violle et al. 2012 ; Moran et al. 2016). En effet, les études à la fois in situ et les expériences en laboratoire montrent que les individus d’une même espèce présentent des caractéristiques génotypiques différentes, s'exprimant par l'existence de phénotypes variés et permettant aux individus des réponses physiologiques et comportementales différentes face à l'environnement (Sourisseau et al. 2008 ; Ollier et al. 2018). Dans le but d'avoir une représentation plus réaliste et une meilleure compréhension des réponses physiologiques de nos populations de krill face à un environnement spatiotemporel changeant, nous avons implémenté dans le modèle physiologique développé lors du second chapitre une variabilité dans les paramètres des équations physiologiques des différents processus, créant ainsi des individus avec leur propre identité. Le troisième chapitre de cette thèse a donc pour objectif de caractériser la contribution relative de la
variabilité environnementale interannuelle (2003 et 2006) et la variabilité interindividuelle sur la production de T. raschii dans le système EGSL. En comparant
les réponses des individus de krill simulés avec ou sans variabilité interindividuelle face à un environnement variant dans le temps et l'espace, nous serons en mesure de déterminer la part relative de chacun des deux types de variabilité (interindividuelle et environnementale) sur la croissance et reproduction de T. raschii.
Cette thèse de doctorat fourni des avancements majeurs dans la compréhension de la dynamique de transport, d'agrégation et de production des deux espèces dominantes de krill dans l'EGSL en intégrant à la fois des échelles spatiales, temporelles et organisationnelles au travers des trois chapitres qui sont représentés de manière schématique dans la figure 0.4 ci-dessous. Cette thèse fournit des éléments qui seront utiles pour la mise en place d'outils de gestion écosystémiques des stocks des espèces fourragères développés par Pêches et Océans Canada.
Figure 0.4 Schéma conceptuel de la structure de la thèse. Intégration des trois chapitres en réponse à la problématique générale dans un contexte spatiotemporel. Les couleurs intègrent le niveau organisationnel de la population du jaune représentant l'individu au rose représentant la population.
I Chapitre 1 - Individual-based modelling explains the
contrasted seasonality in size, growth and reproduction of the
sympatric Arctic (Thysanoessa raschii) and Nordic krill
(Meganyctiphanes norvegica) in the St. Lawrence estuary,
eastern Canada.
I.1 Résumé
Meganyctiphanes norvegica et Thysanoessa raschii dominent la communauté de krill
dans l'EGSL où ils constituent une espèce fourragère centrale pour son écosystème pélagique. Nous avons développé un modèle individuel physiologique spécifique à l'espèce qui met en œuvre les processus physiologiques critiques de croissance, de mue et de reproduction des adultes en réponse aux forçages environnementaux. Les principales innovations de notre approche étaient le découplage entre la mue et la croissance, ainsi que la prise en compte de deux sources distinctes de proies (phyto- et mésozooplancton). Nos résultats ont révélé que les détails du processus d'alimentation étaient essentiels pour une représentation précise de la dynamique de production des deux espèces. Leurs préférences alimentaires spécifiques ont entrainé des schémas phénologiques spécifiques à chaque espèce. Cette étude souligne l'importance d'une connaissance détaillée du régime et du comportement alimentaire des espèces de krill afin d'améliorer notre compréhension des réactions des populations à un environnement changeant.
Mots-clés : Krill, Thysanoessa raschii, Meganyctiphanes norvegica, Physiologie, Phénologie, Modélisation numérique, Estuaire du Saint-Laurent.
I.2 Abstract
The Nordic krill Meganyctiphanes norvegica and Arctic krill Thysanoessa raschii both dominate the krill community within the estuary and Gulf of St. Lawrence system where they are central forage species for its pelagic ecosystem. We developed a species-specific physiological individual based model that implements the critical physiological processes of growth, moulting and reproduction of adults as responses to environmental forcing. Key innovations of our approach were the decoupling between the moulting schedule and growth, as well as considering two distinct sources of prey (phyto- and mesozooplankton). Our simulation results revealed that the details of the feeding process were critical for an accurate representation of the production dynamics of adult individuals from both species. Their specific feeding preferences on phyto- and mesozooplankton resulted in distinct species-specific phenological patterns that reproduced observations. The present study highlights the importance of detailed knowledge of diet and feeding behaviour of krill species to improve our understanding of population responses to a rapidly changing environment.
Keywords: Krill, Thysanoessa raschii, Meganyctiphanes norvegica, Physiology, Phenology, Numerical modelling, St. Lawrence estuary