ETUDES Sra LES SOURCES DE CONTAMWATION PAR LES BACTEMES
RESPONSABLES DE LA NERVATION NOIRE DU CHOU ET DE LA NECROSE
MARGINALE DE LA LAFTUE
par
Elisabeth Wellman-Desbiens
memoire presente au Departement de biologie en vue de 1'obtention du grade de maitre es sciences (M.SC.)
FACULTE DES SCIENCES
UNIVERSITE DE SHERBROOKE
Sherbrooke, Quebec, Canada, octobre 1998
1*1
National Library of Canada Acquisitions and Bibliographic Services 395 Wellington Street Ottawa ON K1AON4 Canada Bibliotheque nationale du Canada Acquisitions et services bibliographiques 395, rue Wellington Ottawa ON K1AON4 CanadaYour file Votre reference
Our file Notre reference
The author has granted a
non-exclusive licence allowing the
National Library of Canada to
reproduce, loan, distribute or sell
copies of this thesis m microfonn,
paper or electronic formats.
The author retains ownership of the
copyright in this thesis. Neither the
thesis nor substantial extracts from it
may be printed or otherwise
reproduced without the author's
permission.
L'auteur a accorde une licence non
exclusive permettant a la
Bibliotheque nationale du Canada de
reproduire, prefer, distribuer ou
vendre des copies de cette these sous
la fonne de microfiche/fihn, de
reproduction sur papier ou sur format
electronique.
L'auteur conserve la propriete du
droit d'auteur qui protege cette these.
Ni la these ni des extraits substantiels
de celle-ci ne doivent etre imprimes
ou autorement reproduits sans son
autonsation.
0-612-56981-0
Le /Q //2/i/(^f^i /^^ , Ie jury suivant a accepte ce memoire dans sa version finale.
<J date
President-rapporteur: M. Claude Dery
Departement de biologie
Membre: Membre: Membre: M. Ryszard BrzezinskiDepartement de biologie
Mme Carole Beaulieu
Departement de biologie
Mme Odile Carisse
Agriculture Canada
l/Lt^y [^
SOMMAIRE
La nervation noire des cruciferes, endemique au Quebec depuis plusieurs annees, est causee par Xanthomonas campestris pv. campestris. Cette bacterie occasionne des lesions jaunatres en forme de « V » sur les feuilles de chou. Par ailleurs, 1'agent responsable de la necrose marginale de la laitue est Xanthomonas campestris pv. vitians. Cette maladie se manifeste par 1' apparition, en marge des feuilles, de taches d'apparence graisseuse devenant necrotiques et confluentes. Les methodes de repression chimique contre la nervation noire sont plus ou moins efficaces. En ce qui conceme la necrose marginale, aucun produit phytosanitaire n'est homologue puisque cette maladie n'est appame que recemment dans nos regions. Au Quebec, les differentes sources de contamination n'ont jamais ete etudiees ce qui represente un frein dans Ie developpement d'un programme de lutte efficace centre ces bacterioses. L'objectif de cette recherche est d'etudier les sources de contamination et de determiner leur importance quant a la propagation de la nervation noire du chou et de la necrose marginale de la laitue.
La premiere section porte sur la propagation de X. campestris pv. vitians en conditions
controlees et consiste a etablir Ie patron de la distribution spatiale de la bacterie sur les jeunes
plantules de laitue en fonction du temps et de la frequence des deplacements de la rampe d'irrigation. II a ete possible de suivre la population epiphyte grace a 1'utiUsation d'une souche bacterienne resistante a la rifampicine. Les resultats demontrent qu'un seul foyer d7 infection representant 0,1% des laitues est suffisant pour entrainer 1'infection de 77 a 99% des quadrats presents sous 1'action de la rampe d'irrigation. De plus, il a ete possible cTajuster une equation de Gaussian pour predire la proportion de quadrats infectes en fonction de la
distance par rapport au foyer d'infection (gradient de dispersion). Ce modele doit cependant
etre valide sous les conditions de production employees chez les serristes.
La seconde section porte sur 1'importance de la survie des agents responsables de la nervation noire du chou et de la necrose marginale de la laitue sous nos conditions hivemales selon Ie type d'enfouissement des debris de culture a Fautomne. Pour cette experience, differents
traitements (debris et sol infectes) ont ete mis en place aux sites selectionnes et Ie suivi de la population bacterienne s'est echelonne de 1'automne jusqu'au debut juin. Les resultats ont revele la capacite de survie de X. campestris pv. campestris et de X. campestris pv. vitians sous nos conditions hivemales a la fois libres dans Ie sol et proteges par les debris de culture.
Les resultats de cette etude pourront servir de base a 1'elaboration d'un programme de lutte contre ces maladies bacteriennes.
REMERCEEMENTS
Mes plus sinceres remerciements aux docteurs Odile Carisse et Carole Beaulieu pour leur expertise, leur confiance, leur support, leurs explications, leur passion, leur devouement et ainsi que leur presence.
Merci beaucoup a Daniel, Vicky, Vincent, Any, Julie, Mohammed, Annie et Melanie pour Ie support technique, 1'ecoute, leur disponibilite, leur bonne humeur, leur patience, pour les moments de confidence, de delire et pour 1'esprit d'equipe.
Toutes mes pensees a mon pere, a ma mere toujours presente dans mon coeur, a Benoit, a Muscade et a mes amis pour leur amour, leurs encouragements, les discussions, les moments de repos et les episodes de folie.
TABLE DES MATEERES
SOMMAIRE...ii
REMERCIEMENTS ... iv
TABLE DES MATffiRES... v
LISTE DES ABREVIATIONS ...viii
LISTE DES TABLEAUX...ix
LISTE DES FIGURES...x
INTRODUCTION...!
CHAPITRE1
ETUDE SUR LA DISPERSION DE XANTHOMONAS CAMPESTRIS PV. VITIANS
EN CONDITIONS CONTROLEES ... 161.1 Introduction ... 16
1.2 Materiel et methodes ... 17
1.2.1 Selection de la souche bacterienne mutante naturellement resistante a un antibiotique ... 17
1.2.2 Comparaison du pouvoir d'adaptation des souches bacteriennes sauvages et des souches mutantes... 17
1.2.3 Etude de la dispersion de X. campestris pv. vitians en conditions controlees
22
1.2.3.1 Mise en place du dispositif experimental en serre ... 22
1.2.3.2 Production du foyer d'infection ... 23
1.2.3.3 Introduction du foyer d'infection ... 24
1.2.3.4 Echantillonnage ... 25
1.2.3.5 Analyses statistiques ... 28
l.SResultats ... 29
1.3.1 Selection de la souche bacterienne mutante resistante a la rifampicine
29
.3.2 Etude de la dispersion de X. campestris pv. vitians en conditions controlees •"» •-» .. JJ 1.4 Discussion ...58CHAPITRE II
ETUDE SUR LA SURVffi DANS LE SOL DES BACTERffiS RESPONSABLES DE
LA NERVATION NOIRE DU CHOU ET DE LA NECROSE MARGINALE DE LA
LAITUE AUX CONDITIONS fflVERNALES...642.1 Introduction ... 64
2.2 Materiel et methodes ... 65
2.2.1 Selection des sites experimentaux... 65
2.2.2 Dispositif experimental...65
2.2.3 Preparation des traitements ...68
2.2.4 Mise en place des parcelles experimentales ... 70
2.2.5 Traitement des echantillons ...71
2.2.6 Tests pour identifier les souches bacteriennes inconnues ... 75
2.2.6.1 Tests de Gram ...75
2.2.6.3 Pouvoirs pathogenes ...76
2.2.7 Analyse des resultats ...78
2.3 Resultats...79
2.4 Discussion ...91
CONCLUSION... 96
LISTE DES ABREVIATIONS
l^g :
nl :
ATCC:
cm :cm
D.O. :FS :
g :
1 :
LSD :
M :
m : m2 •. mg : ml :NB :
Nm :ppm :
SAS :
SM :
ssv ;
sv :
TSA :
UFC :
uv :
microgrammemicrolitre
'American Type Culture Collection' centimetre centimetre cube
densite optique
milieu Fieldhouse-Sasser grammelitre
Least significant difference
molarite
metre metre carremilligramme
millilitre
bouillon nutritif
nanometrepartie par million
Statistical analysis system
milieu 'amidon-methionine' milieu semi-seiectifvitians milieu vitians
tryptic soy agar
unite formant des colonies rayons ultra-violets
LISTE DES TABLEAUX
Tableau 1. Periodes de croissance utilisees pour etablir les courbes de croissance ... 18 Tableau 2. Legende des cotes utilisees pour revaluation visuelle de la necrose marginale
de la laitue ... 21
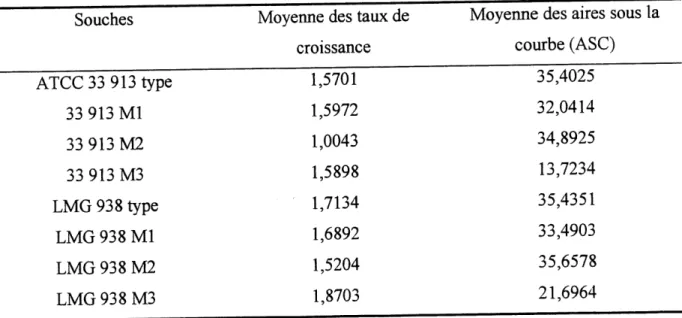
Tableau 3. Resultats de la moyenne des taux de croissance et des aires sous la courbe pour
les souches sauvages et leurs mutants...29
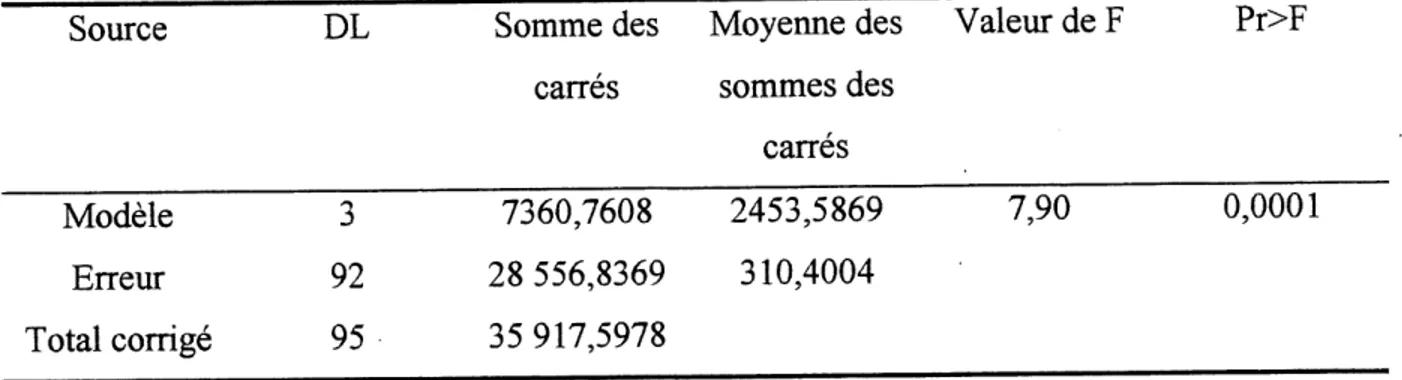
Tableau 4. ANOVA sur les donnees de la severite des symptomes de la nervation noire
.31
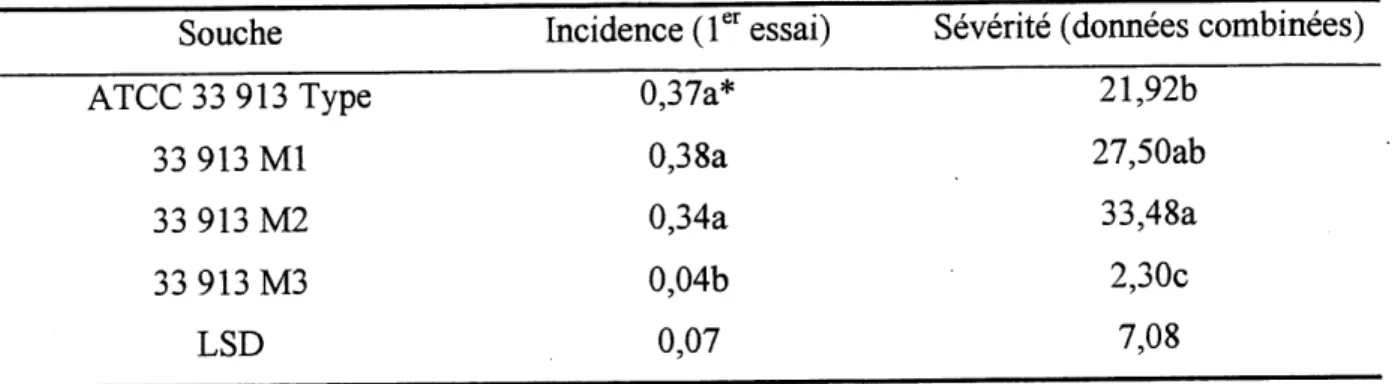
Tableau 5.. Comparaison des pouvoirs pathogenes entre ATCC 33 913 Type (X. campestris
pv. campestris} et les mutants selectiones (33 913 Ml, 33 913 M2 et 33 913
M3)...32
Tableau 6. ANOVA sur 1'effet de la frequence des arrosages ... 58 Tableau 7. Traitements mis en place a Napierville et a St-Jacques-le-Mineur ... 65
Tableau 8. Traitements mis en place aux fermes experimentales de I'Acadie et de
Ste-Clotilde... 66
Tableau 9. Poids des echantillons de laitues infectees pour les deux essais...69
Tableau 10. Programme du cabinet de croissance avec brumisateur (photoperiode de 18
heures)... 77
Tableau 11. ANOVA sur les donnees de survie a St-Jacques-le-Mineur (16 mai 1997 et 13
mai 1998)...80
Tableau 12. Test de comparaisons multiples entre les traitements appliques a
St-Jacques-le-Mineur ... 83
Tableau 13. ANOVA sur les donnees de survie a Napierville (16 mai 1997 et 13 mai 1998)
.86
Tableau 14. ANOVA sur les donnees de la survie a I'Acadie (28 mai 1997 et 13 mat 1998)
.89 Tableau 15. Temperatures (°C) au cours de I'experience de survie a I'Acadie et a
LISTE DES FIGURES
Figure
Figure
Figure
Figure
Figure
Figure
Figure
Figure
Figure
Figure
Figure
Figure
Figure
Figure
Figure
Figure
Figure
Figure
1.2
3. 4. 5. 6. 7. 8. 9. 10. 11. 12. 13. 14. 15. 16. 17. 18.Symptomes de la nervation noire du chou ...4
Symptomes de la necrose marginale de la laitue ...7
Jeunes plantules de laitues en serre avec la rampe d' irrigation ... 24
Impression des feuilles de laitues sur Ie milieu de reisolement ...25
Colonies de X. campestris pv. vitians sur Ie milieu de reisolement...27
Disposition des quadrats infectes par X. campestris pv. vitians a chacun des temps d'echantillonnage au cours du 1 essai...35
Disposition des quadrats infectes par X. campestris pv. vitians a chacun des temps d'echantillonnage au cours du 2 essai ...39
Disposition des quadrats infectes par X. campestris pv. vitians compiles de fa9on cumulative au cours du 1 essai ...43
Disposition des quadrats infectes par X. campestris pv. vitians compiles de fa9on cumulative au cours du 2 essai ...47
Pourcentage cumulatif de quadrats infectes en fonction du nombre de jours (ou d'arrosages) apres 1'introduction du foyer d'infection... 51
Graphiques de modelisation presentant la proportion de quadrats infectes en fonction du nombre de jours apres 1'introduction du foyer d'infection ... 52
Pourcentage de quadrats infectes en fonction de la distance par rapport au foyer d'infection lors du 1CT essai. ...55
Pourcentage de quadrats infectes en fonction de la distance par rapport au foyer d'infection lors du 2 essai. ...56
Modele de Gaussian predisant Ie gradient de disperion (Ie pourcentage de
quadrats infectes en fonction de la distance au foyer d'infection).. ...57Appareil d'acquisition des donnees meteorologiques de type CR10... 67
Enfouissement d'un chou dans un sac d'oignon a 1'automne ...71
Recolte des echantillons au printemps ... 72
Figure 19. Survie de X. campestris pv. campestris en sol mineral a St-Jacques-le-Mineur .81
Figure 20. Survie de X. campestris pv. campestris en sol mineral a Napierville...84 Figure 21. Survie de X. campestris pv. campestris en sol mineral a 1'Acadie ...87 Figure 22. Sunde de Z campestris pv. vitians en sol organique a Ste-Clotilde... 90
INTRODUCTION
La carotte, la laitue. Ie mai's sucre, 1'oignon et Ie chou sont les principales productions maraicheres au Quebec (Bureau de la statistique du Quebec, 1995). En 1994, la valeur de ces cultures englobait pres de 70% de toute la valeur de la production legumiere du Quebec. Avec une consommation annuelle de 34 000 tonnes de choux et de 70 000 tonnes de laitues en 1993, Ie Quebec est en voie d'etre autosuffisant en ce qui conceme ces productions. De fait, dans nos regions, la culture de la laitue augmente considerablement et represente 81% de la production canadienne alors que celle du chou represente 48% de ce meme marche
(Statistiques Canada, 1998).
Depuis de nombreuses annees, ces deux cultures sont sujettes a differentes maladies dont certaines bacterioses. En effet, la nervation noire du chou et la necrose marginale de la laitue prennent de 1'ampleur et causent des pertes importantes. Les plants infectes sont souvent invendables et tres sensibles aux pourritures molles causees par d'autres agents pathogenes soit en champ, soit au cours d'un entreposage. Par contre, lors d'infection legere, les feuilles atteintes sont eliminees a la recolte. Au Quebec, les epidemies sont sporadiques et la gravite de ces infections est liee aux conditions meteorologiques. Ie temps chaud et pluvieux favorisant Ie developpement de ces maladies.
La nervation noire du chou et la necrose marginale de la laitue sont causees par des bacteries du genre Xanthomonas. Ces bacteries, a gamme cThote limitee, possedent la propriete d'infecter une ou plusieurs espece de plantes. On classe les souches d'une meme espece en differents pathovars selon leur pouvoir pathogene sur differentes especes vegetales. Le pathovar donne generalement Ie nom de la plante d'origine (Rat et Chauveau, 1985). Selon leurs genes d'avimlence, les souches pathogenes ne causent la maladie que sur certains cultivars de plantes. On divise alors les souches d'un pathovar en races selon leur capacite a infecter certains cultivars d'une espece vegetale (Minsavage et al., 1990).
Nervation noire du chou
La ner^ation noire des cruciferes est causee par la bacterie, Xanthomonas campestris pv. campestris (Pammel) Dowson. Cette maladie, de distribution universelle, est consideree
comme la plus destructive pour les cruciferes apres la hemie (Schaad et White, 1974a; Schultz
et Gabrielson, 1986; Schaad et Dianese, 1981; Kocks et Zadoks, 1996; Williams, 1980;
Ruissen et al., 1989, Hunter et al, 1975; Dzhalilov et Tiwari, 1995). En 1893, Pammel, aux Etats-Unis, a ete Ie premier a decrire les symptomes de cette bacteriose sur Ie rutabaga (Brass ica campestre). II isola des lesions une bacterie mobile de couleur jaune qu'il baptisa Bacterium campestre (Meier, 1934).
Cette bacterie, colonisatrice de type vasculaire, progresse de la marge des feuilles vers 1'interieur de ces demieres. Les phytobacteriologistes sont generalement en accord pour affirmer que les bacteries pathogenes des plantes ne possedent pas de structures de penetration comme celles retrouvees chez les champignons (Huang, 1986). La bacterie penetre done par les ouvertures naturelles telles les stomates et les hydatodes. Les hydatodes sont des structures semblables aux stomates et situes a Fextremite des nervures en marge des feuilles. Elles assurent Ie phenomene de la guttation resultant d'une forte pression racinaire combinee a un faible niveau d'evapotranspiration (Ruissen et Gielink, 1992). Elle peut aussi entrer par une blessure au niveau des feuilles. Cependant, les hydatodes sont la porte d'entree par excellence pour ces bacteries (Williams, 1980; Ruissen et al., 1989). Toutefois, un niveau d'humidite tres eleve est requis. A ce moment, lorsque la plante gutte, de multiples gouttelettes suintent en marge des feuilles. Les bacteries incorporees dans ces gouttes sont attirees dans les feuilles lorsque les stomates s'ouvrent et que la transpiration s'accelere. Selon les resultats d'une experience effectuee en conditions controlees, les cultivars de choux different dans leur habilete a gutter. En effet, les cultivars sensibles a la nervation noire possedent une capacite
de guttation superieure a celle des cultivars plus resistants (Ruissen et Gielink, 1992). Cette
bacterie produit un polysaccharide extracellulaire, Ie xanthane. Leben (1965) a indique que
certaines bacteries produiraient des substances mucoides protectrices les sauvegardant de la dessication ou restreignant Ie delogement des cellules lors des lavages. De ce fait, des etudes
au microscope electronique revelent la presence de materiel amorphe emergeant entre les cellules bacteriennes sur les feuilles (Mariano et McCarter, 1993; Timmer et al., 1987). Cette
matrice composee de differents types de polysaccharides extracellulaires (PSE) favorise
1'ancrage des cellules au niveau de la surface des feuilles, previent la dessication et modifie Fenvironnement physique et chimique pour Ie rendre favorable a la croissance et a la survie bacterienne (Denny, 1995). Les Xanthomonas sont des producteurs de xanthane (Denny, 1995). Ce compose se revele etre 1'assemblage d'un squelette de cellulose avec des chaines de trisaccharides meublant les cotes. La production de ce compose en combinaison avec les amas de bacteries obstment partiellement Ie xyleme ce qui a comme consequence de restreindre Ie flot de 1'eau et des elements nutritifs. Les premiers symptomes visibles de cette desorganisation se traduisent par 1'apparition du noircissement des canules. Par la suite. Ie manque d'eau a la marge de la feuille induit Ie developpement d'une chlorose. II en decoule des lesions caracteristiques en forme de « V » et de couleur jaunatre a 1'extremite des feuilles (Figure 1). Le noircissement des veines, consequence de la destruction partielle ou complete des vaisseaux, est observable lorsque les lesions prennent de 1'expansion (Rat et Chauveau, 1985). Dans des cas extremes, il n'est pas rare d'observer des poumtures molles causees directement par Fagent pathogene ou par d'autres parasites secondaires comme Erwinia carotovora ou Pseudomonas marginalis (Rat et Chauveau, 1985). L'agent responsable de la nervation noire entrame egalement la formation de symptomes atypiques, confondues avec plusieurs autres desordres, lors d'une infection par une blessure (Rat et Chauveau, 1985). Un simple halo de couleur jaune entoure alors 1'abrasion. Dans un tel cas, la colonisation des vaisseaux est plutot rare.
Figure 1. Symptomes de la nervation noire du chou.
Plusieurs experiences montrent que les souches bacteriennes deficientes dans la production de xanthane sont moins aggressives. Par exemple, 1'infiltration, dans une feuille de navet, d'une suspension bacterienne (10 ufc/ml) mutante dans la production de xanthane cause des symptomes plus discrets que la souche sauvage (Denny, 1995).
Avant la penetration de 1'agent responsable de la nervation noire dans Ie systeme vasculaire des cruciferes, la periode de quiescence correspond a la vie epiphyte de la bacterie a la surface des feuilles de choux. Sa presence dans les gouttes au niveau des hydathodes marque la fin de
la periode epiphyte et Ie debut de la phase infectieuse (Ruissen et al., 1989). Cette periode de
quiescence est variable selon Ie niveau d'humidite. Une experience effectuee par Ruissen et
al. (1989) revele qu'elle peut etre de 6 jours dans un environnement sature en humidite (100%)
et elle se prolonge jusqu'a 13 jours avec un niveau d'humidite de 75% Ie jour et de 100% la
detection de X. campestris pv. campestris dans les fluides au niveau des hydatodes. En fait, peu d'attention a ete portee au phenomene de la guttation comme mode de contamination.
Une temperature entre 25 et 28°C, un niveau d'humidite eleve et des conditons pluvieuses favorisent Ie developpement de la nervation noire du chou entrainant des pertes economiques importantes (Rat et Chauveau, 1985). Par ailleurs, la penetration et 1'evolution de la bacterie est egalement possible a des temperatures se situant entre 16 et 18°C (Hunter et al., 1975).
Necrose marginale de la laitue
Presente au Quebec depuis 1994, la necrose marginale de la laitue est une nouvelle maladie dans la region du sud de Montreal. En 1996, au cours d'un ete chaud et pluvieux, elle a cause des pertes importantes chez les producteurs de la region (Sauriol). D'abondants degats, resultant parfois en une perte totale au moment de la recolte, ont deja ete observes dans plusieurs regions du monde dont 1'Italie, la Califomie, 1'Ohio, la Floride, 1'etat de New York,
1'Afrique du Sud et la Nouvelle-Zelande (Boesewinkel, 1977; Schroth et al., 1964; Wallis et
Joubert, 1972; Zoina et Volpe, 1992; Sahin et Miller, 1997; Burkholder, 1954; Pemezny et al.,
1995).
La necrose marginale de la laitue a ete raportee pour la premiere fois en 1918 par N. A. Brown
(Brown, 1918; Sahin et Miller, 1997). En 1951, Elliot enumerait trois differents types de
bacteries pouvant causer la necrose marginale de la laitue: X. vitians, X. lactucea et X. lactucea scariolae. Une etude ulterieure a permis de classer ces trois types de bacteries comme etant des souches de X. vitians. Dans une premiere reclassiflcation du genre, 1'espece X. vitians a ete renommee X. campestris pv. vitians incluant Ie type A et B (Dye et al., 1980). Dans une etude recente, Vauterin et al. (1995) reclassifiaient Ie genre Xanthomonas sur la base de Fhybridation ADN-ADN. Les deux types, A et B, de X. campestris pv. vitians ont alors ete separes en deux especes differentes : X. axonopodis pv. vitians et X. hortorum pv. vitiuns.
Pour Ie moment, on peut egalement envisager que les deux especes de Xanthomonas soient presentes au Quebec. Pour les besoins de ces travaux et des connaissances presentes, nous referons a X. campestris pv. vitians comme agent responsable de la necrose marginale de la laitue.
Comme pour 1'agent responsable de la nervation noire, X. campestris pv. vitians penetre par les ouvertures naturelles (stomates ou hydatodes) ou encore par une blessure sur les feuilles (Patterson et al., 1986). La necrose marginale de la laitue se manifeste par 1'apparition de petites taches d'aspect graisseux devenant necrotiques et confluentes (Figure 2). On peut egalement observer des taches bmnes circulaires reparties aleatoirement sur la feuille. Les plants infectes sont invendables et tres sensibles aux poumtures molles causees par d'autres agents pathogenes. Dans les cas d'infections legeres, il est possible d'enlever les feuilles atteintes au moment de la recolte. Cependant, les infections plus graves occasionnent la perte totale du plant. Par temps chaud, cette infection evolue souvent vers une pourriture molle generalisee. Le developpement de la maladie est favorise par des periodes de grande humidite, de rosee ou de pluie a une temperature optimale entre 26 et 28°C. II existe, cependant, une possibilite de developpement de la maladie a des temperatures plus basses sur des plantes sensibilisees par Ie gel (Messiaen et al., 1991).
Sources de contamination
L'etude des differentes sources de contamination est essentielle a 1'elaboration de programmes de lutte efficace contre les bacterioses. Par la suite, 11 devient plus facile de faire un choix eclaire quant aux differentes actions a prendre en vue de prevenir et de reprimer ces maladies dans nos regions.
Les semences infectees sont une voie de transmission frequente des bacteries phytopathogenes
(Blakeman, 1991; Clayton, 1929; Monteith, 1921). En effet, plus de 50 especes bacteriennes
Figure 2. Symptomes de la necrose marginale de la laitue.
plusieurs pathovars de Pseudomonas syrmgae {coronafaciens, glycmea, lachrymans, syringae et phaseolicold) et de X. campestris (manhiotis, phaseoli, vesicatoria et campestris) (Hirano et Upper, 1983). Blakemen (1991) nous revele que les semences hebergeant les bacteries proviennent a la fois de plants malades que de plants d'apparence saine. Elles colonisent a la fois 1'exterieur et 1'interieur des semences. Les bacteries peuvent alors contaminer seulement Ie tegument ou encore coloniser les cotyledons au moment de la germination. A ce stade, il est alors possible d'observer, soit des lesions sur les plantules, soit seulement un envahissement epiphyte discret des jeunes plants jusqu'au stade sensible smvenant plus tardivement (Messiaen et al., 1991). Apres la germination des semences, les quelques plantules infectees peuvent alors presenter des symptomes, rester naines ou mourir precocement. C'est a partir de ces premiers foyers d'infection que la maladie se propage aux plantes voisines sous I7 action des pluies et des vents. En champ, 1'importance des semences contaminees dans Ie developpement de la maladie est fonction de 1'interaction entre les
variables environnementales, la densite bacterienne par graine et leur frequence d'infection
(HenisetBashan,1986).
Les semences sont la principale source de contamination de la nervation noire du chou (Cook
et al, 1952; Schaad et Kendrick, 1975). L'infection prealable des semences est possible par la
contamination des fleurs, par 1'infiltration des bacteries au niveau des lesions et des stomates, par leur presence a la surface des graines et par 1'invasion de 1'agent pathogene dans Ie systeme vasculaire des plantes. Cette demiere voie assure 1'infection des semences par X. campestris pv. campestris (Goto, 1992). Les experiences demontrent que trois a cinq semences contaminees sur dix mille sont suffisantes pour causer une incidence elevee de la maladie (Schaad et al., 1980). De plus, la presence en champ d'un seul plant de chou infecte par la bacterie peut s'averer une source d'inoculum suffisante pour entrainer une epidemic de nervation noire (Schaad et Kendrick, 1975). Par centre, en absence de conditions environnementales propices au developpement de 1'agent pathogene, rneme les semences fortement infectees ne donneront pas naissance a des plantules symptomatiques. Weller et Saettles (Hirano et Upper, 1983) affirment qu'une concentration bacterienne par graine de 10 et 10 ufc ( de X. campestris pvs phaseoli ou phaseoli var. fuscans) est necessaire pour permettre 1'infection d'une plante.
Tout comme pour la nervation noire, la principale source de contamination de la necrose marginale de la laitue est constituee par les semences (Sahin et Miller, 1997; Pennisi et Pane,
1990; Zoina et Volpe, 1992; Patterson et al., 1986). D'ailleurs, Sahin et Miller (1997) et
Umesh et al. (1996) ont reussi a produire des semences de laitues infectees a partir de plants artificiellement inocules par X. campestris pv. vitians. La confirmation de la presence de F agent pathogene sur les semences a ete realisee en inoculant ces graines sur un milieu de croissance approprie. Comme cette bacterie pourrait aussi etre de type vasculaire, 1'hypothese qu'elle se localise dans les semences ne peat etre negligee (Sahin et Miller, 1997). Les essais
de Zoina et Volpe (1992) nous revelent que Ie semis de graines artificiellement infectees
donne naissance a des laitues produisant aussi des semences contaminees par 1'agent pathogene. Cependant, d'autres references nous indiquent que les tentatives pour reisoler X.
campestris pv. vitians a partir de lots de semences commerciaux out echoue (Ohata et al.,
1982; Umesh et al., 1996). Zoina et Volpe (1992) contredisent cette affirmation en concluant
que 36% des lots de semences echantillonnes etaient contamines par X. campestris pv. vitians.
Toutefois, les semences ne constituent pas la seule source de contamination de ces bacterioses. A la suite de la recolte des choux et des laitues, il est frequent de retrouver une grande quantite de debris vegetaux infectes laisses dans les champs. Cette matiere vegetale peut servir de source d'inoculum importante pour les bacteries phytopathogenes; Cependant, malgre tout cet inoculum potentiel, les bacteries possedent une capacite limitee de smvie dans Ie sol (Messiaen et al., 1991). Une experience effectuee en conditions controlees a demontre que Ie developpement des symptomes de la moucheture bacterienne des tomates (dont 1'agent responsable est P. syringae pv. tomato) sur les cotyledons de cultivars sensibles etait possible lorsque 1'on seme les graines dans un sol contamine par des feuilles infectees (Schneider et Grogan, 1977). De plus, 1'ajout de particules abrasives a la terre augmente Ie nombre ainsi que la taille des lesions sur les cotyledons emergeant de ce milieu. Malheureusement, 1' importance relative des debris comme source d'inoculum dans Ie developpement des maladies reste peu connue (Hirano et Upper, 1983).
En ce qui conceme la nervation noire, des etudes effectuees en Georgie aux Etats-Unis et dans la region de Moscou en Russie ont demontre la survie de X. campestris pv. campestris dans Ie sol protege par les debris de culture de chou pendant une periode approximative de deux
annees (Schaad et White, 1974b; Dzhalilov et Tiwari, 1995). Ces memes etudes indiquent que
1'agent pathogene peut egalement survivre librement dans Ie sol durant 40 a 60 jours. Une autre etude effectuee dans 1'etat de Washington, revele la presence de la bacterie dans les debris de chou infectes aussi longtemps que ces demiers ne sont pas decomposes. La survie
maximale observee etait de 506 jours pour des debris enterres (Schultz et Gabrielson, 1986;
Strandberg, 1977). II semble que cette survie soit six fois superieure au cours d'une saison de culture froide et humide comparativement a une saison estivale chaude et seche (Schaad, 1976). De plus, la presence de debris peu decomposes de chou, infectes par X. campestris pv. campestris dans un champ, en pleine production, provoque une infection massive des choux
voisins. A Finverse, la presence de debris deja en decomposition entraine un niveau de la nervation noire nettement inferieur. En effet, la decomposition de la matiere vegetale reduit la
population totale de 1'agent responsable de la nervation noire dans ces piles de debris (Kocks
et Zadoks, 1996). En quelque sorte, la survie de X. campestris pv. campestris dans Ie sol depend de la vitesse de decomposition de la matiere vegetale.
Selon des travaux executes en Italie, 1'agent responsable de la necrose marginale de la laitue, X. campestris pv. vitians, possederait la capacite de survivre jusqu'a 40 semaines dans Ie sol en conditions de laboratoire etjusqu'a sept mois apres 1'enfouissement des debris de laitue en
champ (Zoina et Volpe, 1992).
Les facteurs climatiques, les proprietes physico-chimiques, les cultures, Factivite
microbiologique du sol et la production de polysaccharides extracellulaires par les bacteries
affectent la survie des agents pathogenes dans Ie sol (Alvarez et Cho, 1978; Goto, 1992). La
densite de la population de bacteries phytopathogenes dans Ie sol peut se trouver reduite par Ie
labour des residus acccelerant la decomposition, par une rotation des cultures et par de basses temperatures I'hiver (Fukui et al., 1994).
Finalement, les mauvaises herbes et autres plantes non-hotes constituent une source d'inoculum non-negligeable (Young, 1969). Ces mauvaises herbes servent de support physique aux bacteries, permettent leur multiplication et deviennent une source de contamination continuelle. Les pluies combinees a des vents forts assurent la dissemination des bacteries vers des plants sensibles. Aucune donnee quantitative ne revele 1'importance des mauvaises herbes comme source d'inoculum. Cependant, des recherches indiquent qu'il existe une correlation entre une forte densite bacterienne sur les feuilles des mauvaises herbes et 1'incidence de la maladie observee sur les plants adjacents (Mew et Vera Cruz, 1986).
Par ailleurs, plusieurs mauvaises herbes de la famille des cruciferes sont communes dans les champs de production de choux. On soup9onne ces demieres d'etre porteuses de la bacterie causant la nervation noire. Cette supposition s'est averee juste en Georgie et en Califomie,
aux Etats-Unis, ou des experiences ont ete realisees. Les resultats nous indiquent que X. campestris pv. campestris peut etre dispersee sur une distance de plus de 12 metres d'une mauvaise herbe infectee a un chou et ainsi Ie contaminer si les conditions sont propices au developpement de la maladie (Schaad et Dianese, 1981). Dans ces regions, 1'infection des semences par 1'agent responsable de la nervation noire suggere que la bacterie est maintenant endemique chez les mauvaises herbes de la famille des cmciferes. Comme ces especes de mauvaises herbes sont presentes tout au cours de Fannee dans ces zones temperees, 1'agent pathogene est alors en mesure de se propager continuellement d'une generation d'adventices a une autre generation. II est interessant de mentionner que certaines especes de mauvaises herbes contaminees par X. campestris pv. campestris ne developpent pas de symptomes typiques.a la nervation noire (Schaad et Dianese, 1981). En effet, les lesions en forme de « V » sont remplacees par de simples taches jaunes avec un centre bmn. Parfbis, les mauvaises herbes sont meme asymptomatiques.
Une etude menee au Japon a mis en lumiere la capacite de X. campestris pv. vitians a coloniser les mauvaises herbes de fa9on epiphyte (Ohata et al., 1982). De plus, en Califomie, certaines mauvaises herbes appartenant ou non au meme genre que la laitue cultivee (Lactuca scariola, L. canadensis, L. ludoviciana, L. pulchella, Sonchus aleraceus et S. asper) permettent un support physique a X. campestris pv. vitians contribuant a assurer une source de
contamination non negligeable (Schroth et al., 1964). Umesh et al. (1997) out demontre la
presence de la bacterie de fa^on epiphyte a la surface de Matricaria matricarioides indiquant ainsi la possibilite d'une source importante d'inoculum si ces mauvaises herbes sont en
bordure d'un champ de laitues. Sahin et Miller (1998) ont teste la sensibilite de differentes
cultures (tomates, piments, choux, choux fhses, radis, raifort) a la vaporisation de X. campestris pv. vitians en serre. Apres huit jours d'incubation, les plants de tomate et de piment ont montre des symptomes de necrose marginale. Un postulat de Koch a permis de confirmer Ie pouvoir pathogene de 1? agent inocule.
On emploie Ie terme epiphyte pour designer les membres de la microflore qui vivent a la
Hirano et Upper (1983), les surfaces foliaires sont un milieu dynamique ou de multiples
echanges interviennent, selon les conditions environnementales, entre les populations bacteriennes intemes et extemes de la feuille. On considere les bacteries en phase de residence ou epiphyte lorsqu'elles colonisent la surface des plantes hotes ou non-hotes. En effet, les bacteries phytopathogenes et saprophytes partagent ce type de colonisation. Les bacteries pathogenes se multiplient et forment une grande quantite d'inoculum au niveau des parties aeriennes des plantes sans toutefois causer les symptomes de la maladie. Lorsque les conditions environnementales propices a 1'infection sont presentes, il y a explosion des
symptomes par Ie developpement endophyte des agents pathogenes (Hayward, 1974). Ces
microorganismes suivent alors une phase de residence et une phase pathogene. Les bacteries associees aux surfaces aeriennes des plantes sont generalement des batonnets Gram-, ne fbrmant pas de spares et les genres les plus communs sont les Pseudomonas, Xanthomonas et
Envinia (Romantschuk, 1992; Henis et Bashan, 1986).
Comme il a ete mentionne precedemment, les bacteries phytopathogenes sont agglomerees dans une matrice de polysaccharides extracellulaires a la surface des tissus vegetaux (Goto,
1992). Dans ce cas, les cellules bacteriennes ancrees sont difflcilement relachees dans I'air et dispersees vers de nouveaux hotes. La presence d'agents de dispersion tels Ie vent, les pluies, les insectes, la machinerie agricole et les humains est necessaire pour assurer la propagation des bacteries phytopathogenes (Juniper, 1991). Ces agents assurent Ie transfert de 1'inoculum d'une lesion contagieuse vers une nouvelle surface foliaire situee a proximite.
La formation d'aerosols est un mecanisme de dispersion important chez les bacteries
phytopathogenes (Hirano et Upper, 1983). Us sont gener^s par Ie rejaillissement des gouttes
cTeau sur les feuilles. La creation de ces particules balistiques assure Ie transport des bacteries sur une courte distance soit de feuille en feuille d'un meme plant ou d'un plant voisin. En effet, leur cheminement restreint resulte de 1'action des pluies qui engendrent Ie mouvement des bacteries vers Ie sol ainsi que leur redistribution. Cette distance parcoume correspond au gradient de dispersion de 1'inoculum. On definit Ie gradient de dispersion comme etant la decroissance des unites infectieuses en fonction de Feloignement par rapport au foyer
d'infection (Rapilly, 1991). Cette diminution s'explique par la dilution des particules
infectieuses presentes dans 1'air et par leur sedimentation a la surface d'un hote ou au sol. Suite a la vaporisation de plants d'haricot avec une souche de P. syringae pv. syringae
resistante a un antibiotique, la disposition de plats de Petris dans une parcelle experimentale a
permis de retracer la presence des bacteries en suspension au-dessus du couvert vegetal
(Hirano et Upper, 1983). Us ont estime que cent a dix milles bacteries viables etaient alors
deposees passivement sur une feuille de haricot chaque jour et ce, en absence de pluie. La
dispersion et la deposition des bacteries sous des conditions de faible humidite sont des
donnees quantitativement importantes pour I'etude de 1'ecologie des especes bacteriennes.
L'impact des gouttes de pluie sur des feuilles de chou en guttation est un excellent moyen de propagation de X. campestris pv. campestris. En effet, les pluies du matin sont les plus
efficaces puisque la guttation se produit au debut de la joumee (Kocks et Zadoks, 1996).
Alors que X. campestris pv. vitians suinte des lesions, c'est la formation d'aerosols par la combinaison des vents et des pluies qui assure sa dispersion (Patterson et al., 1986). L'irrigation par aspersion ainsi que la pluie sent les meilleurs agents de dispersion de la necrose marginale de la laitue et de la nervation noire du chou (Wallis et Joubert, 1972).
D'ailleurs, la dispersion des bacteries epiphytes de la phyllosphere est consideree importante
dans Ie developpement d'une maladie si, apres attemssage sur une feuille, les colonisateurs survivent, se multiplient et engendrent une population suffisamment elevee pour causer une
infection (Hirano et Upper, 1983).
Moyens de lutte centre ces bacterioses
L'emphase mise sur la repression des maladies fongiques a entrame la decouverte de
nombreux fongicides au detriment de la recherche de bactericides (Swings et Civerolo, 1993).
De plus. Ie developpement de souches bacteriennes resistantes aux produits bactericides rend Ie travail plus difficile (Messiaen et al., 1991). Ce probleme de resistances aux produits est aussi present chez les champignons.
La methode classique de repression contre les maladies bacteriennes est 1'application de produits a base de cuivre (Messiaen et al., 1991). Cependant, ces applications se revelent parfois phytotoxiques pour certaines plantes. Contre les Xanthomonas, V ion zinc s'avere etre un bactericide efficace (Messiaen et al., 1991). La faculte d'adaptation de X. campestris pv. campestris au cuivre pourraient toutefois rendre inutile les interventions chimiques. En effet, en Floride, plusieurs souches de X. campestris pv. vesicatoria sont resistantes au cuivre du a la presence d'un plasmide (Stall et al, 1984). Aucun produit phytosanitaire n'est homologue centre la necrose marginale de la laitue qui est tres recente au Quebec. Dans les deux cas, 1'utilisation d'antibiotiques comme mesure curative est a proscrire puisque ces produits peuvent entrainer Ie developpement de souches bacteriennes resistantes et ainsi nuire a la sante publique.
La prevention, par 1'application de methodes prophylactiques, est la seule alternative possible
(Kocks et Zadoks, 1996; Williams, 1980). Le traitement des semences, 1'elimination des
plants infectes en serre, la rotation des cultures, 1'elimination des debris, la repression des hotes secondaires, 1'usage restreint de 1'irrigation par aspersion ainsi que I'utilisation de cultivars resistants sont des exemples des possibilites a envisager (Hunter et al., 1975). Cependant, 1'identification des principales sources de contamination demeure 1'etape de pour un usage adequat de ces pratiques culturales preventives.
Objectifs
Au Quebec, la presence de ces deux bacterioses limite la production de choux et de laitues. Dans nos regions, les differentes sources de contamination n'ont jamais ete etudiees, ce qui represente un frein dans Ie developpement d'un programme de lutte efficace contre ces maladies. L'objectif de cette recherche est d'etudier les sources de contamination et de determiner leur importance quant a la propagation de la nervation noire du chou et de la necrose marginale de la laitue. Selon la litterature disponible, X. campestris pv. vitians serait viable dans les semences mais difficilement detectable. II nous faut done evaluer les semences
comme source de contamination de la necrose marginale en employant une methode indirecte : la detection sur les jeunes plantules. La premiere section de ce memoire porte sur la propagation de X. campestris pv. vitians en conditions de serre et consiste a quantifier Ie nombre de plants contamines a partir d'un seul foyer de laitues infectees. La seconde section vise a etudier 1'importance de la survie des agents responsables de la nervation noire du chou et de la necrose marginale de la laitue sous nos conditions hivemales selon Ie type d'enfouissement des debris de culture a Fautomne.
CHAPFTRE I
ETUDE SUR LA DISPERSION DE XANTHOMONAS CAMPESTRIS PV. VITIANS EN
CONDFTIONS CONTROLEES
1.1 Introduction.
Selon les resultats d'une recente etude, les semences de laitues produites sur des plants infectes par X. campestris pv. vitians peuvent etre contaminees (Sahin et Miller, 1997). Cependant, les tentatives entreprises pour reisoler la bacterie des lots de semences commerciaux ont echoue (Ohata et al., 1982; Umesh et al., 1996). L'agent responsable de la necrose marginale de la laitue pourrait eft-e viable sur ou dans les semences mais difficilement cultivable sur un milieu de culture. La detection de la bacterie a la surface des jeunes plantules sera la methode employee pour realiser cette experience.
Si les resultats nous indiquent qu'un petit nombre de plants infectes est suffisant pour contaminer une table entiere de production de laitues en serre, alors, il sera pertinent de developper des outils de detection de la bacterie sur les semences ou les plantules. De plus, il sera interessant de mettre 1'emphase sur 1'elaboration de divers traitements a la fois au niveau
des semences et en sen-e.
L'objectifde ce chapitre est d'etudier la distribution spatiale de X. campestris pv. vitians sur les jeunes plants de laitue en fonction du temps et de la frequence des deplacements de la rampe d'irrigation.
1.2 IVIateriel et methodes.
1.2.1 Selection de la souche bacterienne mutante naturellement resistante a un antibiotique.
Pour realiser cette experience, nous avons employe une souche mutante de X. campestris pv. vitians resistante a un antibiotique afin de suivre precisement la bacterie au cours de ses
deplacements et de faciliter sa recuperation. La recherche de mutants naturels a ete effectuee a la fois pour X. campestris pv. campestris et pour Z campestris pv. vitians. L'antibiotique selectionne etait la rifampicine. Apres sa dissolution dans Ie methanol, il a ete incoq)ore aseptiquement a raison de 100 p.g/1, a un milieu du culture TSA 10% prealablement autoclave. Par la suite, ces milieux ont et^ ensemences avec des cultures bacteriennes de reference : ATCC 33 913 pour X. campestris pv. campestris et LMG 938 pour X. campestris pv. vitians. L'inoculum a ete prepare en faisant croitre les souches dans un bouillon NB a 28°C pendant 14 a 16 heures et sous une agitation constante. Apres ajustement de la suspension bacterienne a une D.O. de 0,8 a 550 nm, 100 jjl des suspensions bacteriennes ont ete transferes sur Ie milieu contenant 1'antibiotique. Apres 1'incubation des boites de Petri a 28°C pendant quatre jours, trois colonies bacteriennes resistantes ont ete selectionnees au hasard et repiquees sur
milieu TSA 10% supplemente de 50 |Lig/l de rifampicine pour leur conservation et les tests
futurs.
1.2.2 Comparaison du pouvoir d'adaptation des souches bacteriennes sauvages et des
souches mutantes.
Plusieurs tests ont ete effectues pour detecter les differences comportementales entre les souches bacteriennes mutantes et les souches sauvages.
Pour determiner les courbes de croissance, huit souches bacteriennes ont ete selectionnees:
ATCC 33 913 Type, 33 913 mutant (M) 1, 33 913 M2, 33 913 M3, LMG 938 Type, LMG 938
Ml, LMG 938 M2 et LMG 938 M3. L'inoculum de depart a ete prepare en faisant croitre les
differentes souches dans un bouillon NB a 28°C pendant 14 a 16 heures sous une agitation constante. Les suspensions bacteriennes ont ete ajustees a une D.O. de 0,8 a 550 nm et conservees a 4°C pour s'assurer d'utiliser Ie meme inoculum de depart pour les differentes heures d'ensemencement. L'inoculation des tubes de depart a ete effectuee a raison de 100 jnl par eprouvette contenant 10 ml de bouillon NB. Afin de generer Ie plus de points possibles pour constituer la courbe de croissance, les differents bouillons ont ete inocules, en duplicata, a un intervalle de trois heures soit a lOhOO, a 13h00 et a 16h00. Pour chacun des bouillons, la D.O. a 550 nm a ete relevee a toutes les 12 heures suivant Ie premier ensemencement et s'est poursuivie pendant 48 heures (Tableau 1). L'experience a ete menee trois fois.
Tableau 1. Periodes de croissance utilisees pour etablir les courbes de croissance.
Heures de lecture Heures d'ensemencement et duree des periodes de croissance (heures)
lOhOO 13h00 16h00
9 6
21 18
33 30
45 42
II a ete possible de creer un graphique presentant la D.O. en fonction de la duree d'incubation pour chacune des souches bacteriennes sauvages et mutantes. Le taux de croissance de chacune des souches bacteriennes a ete determine en divisant la valeur de la D.O. maximale par Ie nombre d'heures d'incubation. De plus, Paire sous la courbe a ete calculee pour chacune des courbes avec la fonnule suivante :
Aire sous la courbe : S ((Y^ + Y,+i)/2) * (Xi+i - Xi) (Eq. 1.1)
22h00 (Jour 1)
10hOO(Jour2)
22h00 (Jour 2)
lOhOO (Jour 3)
12
24
36
48
Yi= densite optique au temps X Yi+i= densite optique au temps X+l
Xi+i= nombre d'heures d'incubation au temps X+l Xi= nombre d'heures d'incubation au temps X
Par la suite, une analyse de variance (ANOVA) des aires sous la courbe et du taux de croissance a ete effectuee pour determiner les differences de croissance bacterienne entre les souches testees.
La capacite des bacteries epiphytes a crottre sur Ie milieu semi-selectif a ete evaluee de la fa<?on suivante. Des feuilles de laitue romaine (cultivar Paris Island) et de chou (cultivar Bartollo) provenant de plants cultives en serres ont ete trempees sous une faible agitation dans un becher d'eau distillee sterile pendant une dizaine de minutes. Par la suite, 200 ^1 de cette
eau de lavage a ete prelevee pour inoculer 10 Petris de TSA 10% contenant 50 jig/1 de
rifampicine et de cycloheximide (inhibe la croissance des eucaryotes). Les boites de Petri ont
ete incubees pendant quatre jours a 28°C.
De plus, les mutants out ete selectionnes en fonction de leur capacite a coloniser la surface
foliaire de leur plante hote. La colonisation epiphyte de la phyllosphere a ete determinee par
la technique d'impression des feuilles (Jacques et Morris, 1995) sur Ie milieu selectif. Apres
avoir vaporise cinq plants de chou et de laitue avec les souches mutantes, les plants out ete places en serre pour pemiettre la colonisation des surfaces foliaires par les bacteries mutantes. Apres six heures d'incubation, les feuilles des plants de chou et de laitue etaient bien seches. Une legere pression de la surface superieure et inferieure de deux feuilles par plant a ete realisee sur Ie milieu amende avec la rifampicine a 1'aide d'une tige de bois sterile. Apres Fincubation des boites a 28°C pendant trois a quatre jours, la presence de colonies rondes, bombees, agglomerees et de couleurjaune a ete notee.
En outre, des tests d'hypersensibilite sur des plants de tabac (cultivar Xanthi) ont ete executes selon la methode elaboree par Klement et al. (1964). En effet, ce test permet de s'assurer que les bacteries ont conserve leur pouvoir phytopathogene.
L'etape de dans la selection de la souche bacterienne mutante est la comparaison des pouvoirs pathogenes entre les differents mutants et les souches sauvages. La methode d'inoculation
utilisee a ete la vaporisation des jeunes plants de chou et de laitue au stade de deux feuilles
(Chapitre 2, section 2.6.3). Les souches employees etaient les memes que celles decrites precedemment. Le dispositif experimental consistait en blocs complets aleatoires avec quatre repetitions (blocs) et trois echantillons. L'experience a ete menee deux fois.
A la suite de 1'apparition des premiers symptomes sur les choux, soit apres une periode d'incubation de 14 jours, Fincidence ainsi que la severite de la nervation noire ont ete evaluees. Pour quantifier 1'incidence de la maladie. Ie nombre de feuilles infectees sur Ie nombre total de feuilles du plant a ete determine. La severite de la nervation noire a ete evaluee en detachant et en separant plant par plant les feuilles presentant les symptomes typiques de la maladie. Elles ont ete placees sous une presse pendant 12 heures pour les rendre planes et bien uniformes. La surface foliaire totale de chacune de ces feuilles a ete determinee a 1'aide d'un planimetre. Apres avoir decoupe les portions infectees, la surface de la partie saine etait calculee avec ce meme appareil. La valeur de .la severite a ete calculee en determinant Ie pourcentage des zones infectees par rapport a la surface totale de la feuille pour chacun des plants de chou.
Les tissus vegetaux des laitues ne permettant pas Ie meme genre d'evaluation que les choux, F incidence et la severite de la necrose marginale a ete quantifiiee par une evaluation visuelle des plants. Une cote a ete donnee pour chacune des feuilles des laitues suivant la charte suivante (Tableau 2). Apres cette evaluation visuelle, une cote globale par laitue a ete calculee en multipliant chacune des cotes avec Ie nombre de feuilles infectees associees puis en additionnant ces cinq cotes partielles.
Tableau 2. Legende des cotes utilisees pour revaluation visuelle de la necrose marginale de la laitue.
Cote Description
0 Pas de symptome
1 1-5 lesions en marge des feuilles
2 Plus de 5 lesions en marge des feuilles
3 Lesions coalescentes en marge des feuilles 4 Lesions penetrantes dans les feuilles 5 Feuille senescente
Les resultats obtenus pour les tests du pouvoir pathogene des choux et des laitues ont ete
analyses a Faide du logiciel statistique, SAS (Statistical Analysis System). Un test
d'homogeneite des variances (test de F) a ete effectue afln de verifier si les donnees des deux
essais pouvaient etre combinees. Par la suite, une analyse de variance (ANOVA) a ete effectuee avec les donnees des deux essais combinees. L'analyse de variance permet d'individualiser I'influence de differents facteurs sur un parametre a mesurer (Falissard,
1996). De plus, un test de comparaisons multiples de type LSD (Least Significant Difference)
a permis de regrouper les souches bacteriennes n'etant pas significativement differentes les unes des autres. Ce test est base sur la valeur minimale separant deux moyennes pour qu'on
lesjuge significativement differentes (Falissard, 1996).
Pour s'assurer que les souches bacteriennes mutantes possedaient un metabolisme biochimique semblable a la souche sauvage, 1'identification des souches a ete effectuee par Ie
systeme Biolog GN Microplates (Toussaint).
1.2.3 Etude de la dispersion de X. campestris pv. vitians en conditions controlees.
1.2.3.1 Mise en place du dispositif experimental en serre.
Afin de recreer un environnement semblable a celui des serres commerciales. Ie dispositif experimental a ete elabore pour se conformer aux differents parametres etablis par les serriculteurs (dimension des plateaux, semis, terreau, fertilisation, etc).
Le semis des laitues romaines sensibles a la necrose marginale (cultivar Ideal Cos) a ete
realise dans des plateaux de 338 (13 x 26) cellules remplis de Promix®. Le semis du 1 essai
a ete effectue de fa^on manuelle alors que les plateaux du 2 essai out ete semes de fa^on commerciale pour assurer une meilleure uniformite du couvert vegetal au moment de la
germmation.
Pour effectuer 1'experience, quatre tables de serre rassemblees couvraient une superficie de 7,4 m2. Une rampe d'irrigation a ete installee sur rails et 1'usage d'une corde flxee au centre de cette demiere a permis son deplacement lors des arrosages. Au cours du 1 essai, la rampe etait composee de quatre buses a jet de 80 degres distantes de 38 cmet a une hauteur de 56 cm au-dessus du couvert vegetal. En effet, selon cette hauteur theorique, on obtient 200% de recoupement des jets c'est-a-dire qu'un seal jet couvre deux fois la distance entre les buses (Boudreau1). La calibration des buses a ete verifiee a 1'aide d'un chronometre et d'un becher
gradue sur une periode de 60 secondes. Par la suite, il a ete possible de quantifier Ie debit de
chacune des buses (litre/minute) pour s'assurer de leur uniformite. A la suite du 1 essai, nous avons constate que les laitues situees aux extremites de la table avaient manque d'eau. Pour ameliorer la qualite de 1'arrosage, deux nouvelles buses de meme type ont ete ajoutees aux extremites de la rampe a une distance de 38 cm. Ces buses out ete calibrees pour verifier I'uniformite du debit de 1'eau tel que decrit precedemment.
La temperature de la serre a ete fixee a 20°C, cependant, au cours des joumees ensoleillees, elle pouvait atteindre 30°C. Toutefois, une climatisation permettait de maintenir les conditions relativement constantes.
U irrigation des laitues etait effectuee au besoin par Ie deplacement de la rampe permettant 1'arrosage manuel et Ie controle de la vitesse pour une bonne humidification du sol. Apres la germination des laitues, elles ont ete fertilisees a chaque arrosage selon la regie suivante : engrais 15-15-18 a raison de 80 ppm par litre de concentre.
1.2.3.2 Production du foyer d'infection.
Pour simuler une contamination naturelle, Ie foyer a ete produit par 1'infection des semences de laitues avec la souche de X. campestris pv. vitians mutante. La methode employee a ete elaboree par Salcedo et al. (1992). Les semences de laitue ont ete denudees de leur enrobage argileux par un trempage sous agitation dans 1'eau. La suspension visqueuse resultante a ete filtree a 1'aide d'un coton fromage pour recuperer les semences de laitues nues. Par la suite, les graines de laitue ont ete sterilisees, sous une faible agitation, dans one solution d'eau de Javel (0.01%) pendant cinq minutes et rincees successivement dans trois differents bains d'eau distillee. Les graines ont ete placees dans une boite de Petri sterile et sechees sous la hotte a flot laminaire. Par la suite, elles ont ete deposees dans un erlenmeyer sterile avant d'appliquer Ie vide (vacuum) pendant 5 minutes. On a alors ajoute 50 ml de la suspension bacterienne
(LMG 938 M3 prealablement cultivee dans un bouillon de culture NB a 28°C pendant 14 a 16
heures sous une agitation constante). Le vide a ete applique de nouveau pendant 5 minutes. Les graines ont finalement ete rincees vigoureusement a 1'eau distillee sterile et sechees sous la hotte a flot laminaire. On les a conservees dans un tube de type Falcon sterile a 4°C.
Les semences contaminees ont ete semees et maintenues sous les memes conditions que les laitues saines en serre.
1.2.3.3 Introduction du foyer d'infection.
L'experience a debate a 1'emergence des cotyledons de laitues. Le foyer d'infection a ete introduit a la fin du premier tiers de la table de serre. Six laitues saines ont alors ete remplacees par 12 laitues contaminees par X. campestris pv. vitians resistantes a la rifampicine (a raison de deux par cellule). Le foyer d'infection representait 0,1% des laitues presentes dans la serre. A la suite de V introduction du foyer, les laitues ont ete arrosees avec la rampe d'irrigation pour simuler les conditions commerciales (Figure 3).
1.2.3.4 Echantillonnage.
La table de serre a ete divisee en plusieurs petits quadrats de neuf laitues. A chacun des echantillonnages, qui out eu lieu 24 heures apres 1'arrosage, une laitue par quadrat a ete selectionnee au hasard et les feuilles ont ete imprimees sur la gelose de reisolement. Ces impressions ont ete effectuees a 1'aide de batons de bois sterilises et changes a chaque quadrat pour eviter de contaminer artificiellement les autres laitues (Figure 4). A chacun des echantillonnages, 2912 laitues ont ete imprimees, ce qui represente 25% de la totalite des plantules.
Le ler essai a ete mene au cours de I'hiver. A cette epoque, la temperature plus fraiche de la sen-e etait beaucoup plus adaptee a la croissance des laitues et les arrosages etaient fixes a tous les deux jours. Par contre, lors du 2 essai, au printemps suivant, les laitues devaient etre arrosees a tous les jours. Cependant, 1'echantillonnage s'est poursuivi a tous les deux jours.
L'experience s'est deroulee sur une periode de 25 et de 23 jours pour Ie 1CT et Ie 2 essai respectivement et, dans chacun des cas, 12 echantillonnages ont ete faits. Cette periode correspond au temps normal de la production de plants de laitue en serre. Le ler echantillonnage a ete effectue avant 1'introduction du foyer d'infection pour s'assurer que les laitues etaient exemptes de X. campestris pv. vitians resistantes a la rifampicine.
Apres 1'incubation des boites de Petri a 28°C pendant six jours, la presence ou 1'absence de X. campestris pv. vitians a ete notee pour chacun des quadrats (Figure 5).
ISA iu% + 50 mg/1 rifamplcin
LMG 938 M3
Figure 5. Colonies de X. campestris pv. vitians sur Ie milieu de reisolement.
Legende : Sur ce milieu, les colonies de X. campestris pv. vitians sont rondes, mucoi'des, de couleurjaune et souvent en agglomerat. Ces boites de Petri presentent aussi des contaminants.
1.2.3.5 Analyses statistiques.
Les analyses statistiques ont ete faites a 1'aide du logiciel SAS et du logiciel Table Curve.
Elles ont ete effectuees separement pour les deux essais, les variances n'etant pas homogenes. Premierement, les donnees ont ete compilees de fa^on a permettre Ie calcul de la distance de chacun des quadrats par rapport au foyer d'infection. Par la suite, les donnees ont ete exprimees de fa<?on cumulative afin de detemiiner Ie temps requis pour atteindre une incidence de 100%. Alors, on considere qu'un quadrat infecte reste infecte jusqu'a la fin de F experimentation. A I'aide de ces donnees combinees. Ie patron d'incidence de la maladie a ete visualise en effectuant des graphiques representant Ie pourcentage de quadrats infectes en fonction du nombre de jours apres 1'introduction du foyer d'infection, en fonction du nombre d'arrosages et en fonction de la distance par rapport au foyer d'infection pour chacun des temps. D'ailleurs, Ie gradient de dispersion se definit comme etant Ie pourcentage de quadrats infectes en fonction de la distance par rapport au foyer d'infection.
A Fissue de 1'observation des graphiques du gradient de dispersion, une equation a ete ajustee pour predire Ie pourcentage de quadrats infectes en fonction de la distance par rapport au foyer d'infection. Le choix de 1'equation s'est fait selon les criteres suivants : une correspondance avec la fomie de la courbe recherchee, une equation pouvant ajuster les donnees des deux essais. Ie meilleur coefficient de determination possible (R ), Ie nombre de parametres dans
1' equation, la signification biologique du modele ainsi que la normalite de la distribution des
residus.
Ensuite, une equation a ete ajustee sur les donnees des deux essais presentant Ie pourcentage de quadrats infectes en fonction du nombre de jours (ou d'arrosages) apres 1'introduction du foyer d'infection. Cette equation predira la proportion de quadrats infectes en fonction du nombre de jours ou cTarrosages apres 1'introduction du foyer d'infection. Le choix de I'equation a ete effectie selon les criteres cites precedemment.
Finalement, une analyse de variance (ANOVA) a ete effectuee pour determiner 1'effet de la frequence des arrosages sur la proportion de quadrats infectes.
1.3 Resultats.
1.3.1 Selection de la souche bacterienne mutante resistante a la rifampicine.
Les calcul des aires sous la courbe nous ont revele une similitude de la croissance et de 1'allure
des graphiques entre ATCC 33 913 Type, 33 913 Ml et 33 913 M2 (Tableau 3). Cependant,
F observation des resultats du calcul des taux de croissance nous indique une similitude entre
la souche ATCC 33 913 Type, 33 913 Ml et 33 913 M3. En ce qui conceme X. campestris
pv. vitians, seul LMG 938 M3 demontre un plus faible resultat de 1'aire sous la courbe.
Tableau 3. Resultats de la moyenne des taux de croissance et des aires sous la courbe pour les souches sauvages et leurs mutants.
Souches
ATCC33913type
33 913 Ml
33 913 M2
33 913 M3
LMG 938 type
LMG 938 Ml
LMG 938 M2
LMG938M3
Moyenne des taux de
croissance
1,5701
1,5972
1,0043
1,5898
1,7134
1,6892
1,5204
1,8703
Moyenne des aires sous la
courbe (ASC)
35,4025
32,0414
34,8925
13,7234
35,4351
33,4903
35,6578
21,6964
L'ANOVA sur les donnees du taux de croissance et des resultats du calcul des aires sous la
souches sur les differences de croissances bacteriennes. En effet, les probabilites associees sont respectivement de 0,8177 et de 0,0735. Ces conclusions sont les m^mes pour 1'analyse de variance des souches de X. campestris pv. vitians (P= 0,9758 et 0,3208).
L'ensemencement de 1'eau de lavage des feuilles de chou et de laitue sur un milieu de culture avec rifampicine n'a entraine aucune croissance bacterienne apres 1'incubation des boites de Petri. On peut alors conclure que les bacteries composant la flore normale des feuilles de chou et de laitue ne se developpent pas sur Ie milieu semi-selectifemploye pour recuperer la souche bacterienne mutante lors de 1'experience de dispersion.
Apres la colonisation des feuilles de chou et de laitue par les differents mutants, la technique d'impression des feuilles a permis la recuperation des souches bacteriennes ensemencees.
A la suite de la periode d'incubation des plants de tabac en serre, les plages de necrose obtenues par les souches bacteriennes testees n'indiquent aucune difference entre les souches sauvages et les mutantes.
II a ete possible de combiner les donnees des deux essais portant sur la comparaison de la severite des symptomes de la nervation noire suite a un test d'homogeneite des variances. En
effet. Ie rapport entre la variance de 1'essai un (trois degres de liberte) et de 1'essai deux (trois
degres de liberte) est inferieur a la valeur de F critique a une probabilite de 0,05
(1,9189<9,28). Cependant, suite a ce meme test, il a ete impossible de combiner les donnees d'incidence de la maladie puisque la valeur du rapport des deux essais est superieure a la
valeur de F critique a une probabilite de 0,05 (80,4103>9,28).
L'analyse de variance sur les donnees combinees de la severite des symptomes de la nervation
Tableau 4. ANOVA sur les donnees de la severite des symptomes de la nervation noire.
Source DL Somme des Moyenne des Valeur de F Pr>F carres sommes des
carres
7,90 0,0001
Modele
ErreurTotal corrige
3
92
95
7360,7608
28 556,8369
35 917,5978
2453,5869
310,4004
Les souches mutantes de la nervation noire du chou ont eu un effet significatif sur 1'incidence (P<0,0001). Toutefois, 1'analyse de variance sur les donnees du 2 essai n'a revele aucune difference significative entre la souche sauvage et les mutants au niveau de 1'incidence de la
bacteriose (0,7338>0,05).
Par la suite, un test de comparaison multiple, LSD (voir Tableau 5), a revele que Ie mutant 33 913 M3 est nettement moins agressifque les trois autres souches. L'incidence ainsi que la severite des symptomes causees par cette souche sont significativement inferieures a celles des autres souches. Ce test demontre des differences au niveau de 1'incidence et de la severite de la neryation noire entre la souche 33 913 M3 et les autres souches. De plus, la souche 33 913 M2 semble beaucoup plus agressive que la souche type. Cependant, on ne distingue aucune difference significative au niveau de 1'incidence de la maladie sur les plants de choux pour la
ATCC 33 913 Type
33 913 Ml
33 913 M2
33 913 M3
LSD
0,37a* 0,38a0,34a
0,04b
0,07
Tableau 5. Comparaison des pouvoirs pathogenes entre ATCC 33 913 Type (X. campestris
pv. campestris) et les mutants selectionnes (33 913 Ml, 33 913 M2 et 33 913 M3).
Souche Incidence (1 essai) Severite (donnees combinees)
21,92b
27,50ab
33,48a
2,30c
7,08
*Valeurs avec la meme lettre dans une meme coloime ne sont pas significativement differentes
selon Ie test de LSD (Steel et al., 1980)
II a ete possible de combiner les donnees des deux essais portant sur la comparaison des pouvoirs pathogenes entre X. campestris pv. vitians et ses mutants suite a un test d'homogeneite des variances. En effet, Ie rapport des variances entre 1'essai un (deux degres
de liberte) et I'essai deux (deux degres de liberte) etait inferieur a la valeur de F critique
foumit par la table a une probabilite de 0,05 (2,1079<19,00).
Les resultats de Panalyse de variance portant sur la comparaison des pouvoirs pathogenes
entre LMG 938 Type (X. campestris pv. vitians) et les mutants LMG 938 M2 et LMG 983 M3
n'ont revele aucune difference significative entre ces souches (P=0,l 129).
Les resultats des profils biochimiques (systeme Biolog GN Microplates) nous indiquent que
les mutants font partie de la meme branche phylogenique que leur souche sauvage associee. Us possedent alors un profil nutritionnel tres similaire a celui des souches sauvages
(Toussaint).
1.3.2 Etude de la dispersion de X. campestris pv. vitians en conditions controlees.
L'observation des donnees bmtes des 1 et du 2 essais a chacun des 11 temps d'echantillonnage nous demontre une distribution aleatoire des quadrats infectes autour du
foyer d'inoculum (Figure 6 et 7). En effet, ces graphiques representent la disposition des
quadrats infectes par X. campestris pv. vitians a chacun des jours d'echantillonnage. On observe que 1'envahissement epiphyte de X. campestris pv. vitians sur les laitues est superieur lors du 2 essai (Figure 7). Au cours du ler essai, c'est 1'echantilloimage effectue sept jours apres 1'introduction du foyer d'infection qui revele la plus grande proportion de quadrats infectes representant 20,1% de la totalite des quadrats. On remarque une colonisation massive par X. campestris pv. vitians des Ie 5emejour du 2eme essai. En effet, 57,8% des plants de laitue echantillonnes etaient porteurs de 1'agent responsable de la necrose marginale. Toutefois, c'est au 6 echantillonnage que la proportion des quadrats infectes etait la plus grande (64,4%). II a ete possible de recuperer la bacterie a la surface des laitues composant Ie foyer d'inoculum a partir du 36me et 5emejour apres Ie debut de 1'experience respectivement pour Ie
leretle2emeessai.
Par la suite, les donnees ont ete compilees de fa^on cumulative pour chacun des 11 echantillonnages en considerant qu'une fois un quadrat est colonise, ce demier presente X. campestris pv. vitians]usqu7a. la fin de 1'experimentation. Lors des deux essais, une incidence de 100% n'a pas ete atteinte (Figure 8 et 9). En effet, on remarque une augmentation graduelle de la proportion de quadrats infectes au cours du 1 essai pour atteindre un
maximum de 77,7% Ie 25 jour apres 1'introduction du foyer d'inoculum (Figure 8). Les
donnees du 2eme essai indiquent une augmentation plus rapide de la proportion des quadrats infectes (Figure 9). On denote une augmentation de 59,8% a 99,4% entre Ie 5eme et Ie 23eme jour apres 1'introduction du foyer d'infection.
II est possible de visualiser la progression de 1'incidence dans Ie temps en observant les graphiques presentant Ie pourcentage de quadrats infectes en fonction du nombre de jours ou cTarrosages apres 1'introduction du foyer d'inoculum (Figure 10). Lors du 1 essai, les laitues
ont ete arrosees a tous les deux jours et les jours cTechantillonnage se confondent avec les arrosages. Cependant, les laitues du 2eme essai ont ete irriguees quotidieimement afin d'assurer une bonne croissance des plants. II est possible de tirer les memes conclusions que celles citees precedemment a la suite de 1'observation de ces graphiques. Pour chacun des essais, il a ete possible d'ajuster one equation mathematique decrivant Ie pourcentage de quadrats infectes en fonction du nombre de jours (ou d'arrosages) apres 1'introduction du foyer d'infection. Les modeles retenus selon les criteres selectionnes sont:
Essai 1 : Y= A + BX°'5 (equation de forme lineaire) (Eq. 1.2)
Y= pourcentage de quadrats infectes A= ordonnee a 1'origine
B= parametre associe au taux de progression
X= temps (nombre de jours apres 1'introduction du foyer d5 infection)
Essai 2 : Y= A + B/X0'5 (equation de forme non-lineaire) (Eq. 1.3)
Y= pourcentage de quadrats infectes A= coefficient de regression
B= coefficient de regression
X= temps (nombre de jours apres 1'introduction du foyer d'infection)
Les equations sont:
Essai 1 : Y= -14,9094 + 19,5833 X0'5 (X ^ 0,87) (Eq. 1.2)
Essai 2 : Y= 128,2888 - 145,0233 / X0'5 (X ^ 1,06) (Eq. 1.3)
Ces 2 modeles expliquent respectivement 98,4 et 84,2% des variations pour Ie 1CT et Ie 2 essai. Les graphiques issus de ces equations sont representes a la Figure 11. Pour ces deux modeles, les residus etaient normalement distribues.
^->
U)
^5!55^?^twS5S^I?!3i^IFt^?:iS3iS R!a?i!'3S&.aSSSSS,!SSSKSS??'.";?!?l?!;55SESS3S
m
I
0 <u '0 D 3 bo^
-36-UJ