Caractérisation d’anticorps anti-ectonucléotidases par
immunohistochimie et immunolocalisation de ces
enzymes dans le cordon ombilical et dans le rein
humain
Mémoire
Cosme Jacques Hervé Mahoulome Agonsanou
Maîtrise en microbiologie-immunologie
Maître ès sciences
(M. Sc.)Québec, Canada
Caractérisation d’anticorps anti-ectonucléotidases par
immunohistochimie et immunolocalisation de ces
enzymes dans le cordon ombilical et dans le rein
humain
Mémoire
Cosme Jacques Hervé Mahoulome Agonsanou
Sous la direction de :
iii
RÉSUMÉ
Les ectonucléotidases sont des familles d’enzymes présentes au niveau de la membrane cellulaire et ayant pour rôle d’hydrolyser les nucléotides en nucléosides. L’objectif principal de cette étude était d’abord de caractériser des anticorps produits dans le laboratoire contre les ectonucléotidases afin de les utiliser pour définir l’expression de ces enzymes dans le cordon ombilical et dans le rein humain par immunohistochimie. Les ectonucléotidases qui nous ont intéressées dans ce travail sont les Nucléosides Triphosphates Diphosphohydrolases (NTPDases) de la membrane cytoplasmique, soient les NTPDase1, -2, -3, -8, ainsi que l’ecto-5'-Nucléotidase (CD73). Nous avons aussi localisé ces enzymes dans ces tissus par histochimie enzymatique pour vérifier l’immunolocalisation obtenue et aussi pour vérifier si ces enzymes sont bel et bien actives dans les structures détectées par immunohistochimie.
Nos résultats suggèrent que, dans le cordon ombilical, les NTPDase1, -2, -3, et l’ecto-5'-Nucleotidase sont localisées au niveau de l’endothélium des artères et de la veine et au niveau de la gelée de Wharton. Au niveau du rein, la NTPDase1 est localisée dans les vaisseaux sanguins, la NTPDase2 est localisée sur la face apicale des tubules rénaux, la NTPDase3 ainsi que la NTPDase8 sont localisées dans les tubules rénaux.
L'identification de la localisation cellulaire de ces enzymes dans le cordon ombilical et dans le rein humain pourra nous aider à définir le rôle de ces enzymes dans ces organes et par extension pourra nous informer sur le rôle potentiel des nucléotides extracellulaires dans ces tissus.
iv
ABSTRACT
Ectonucleotidases are families of enzymes present in the cell membrane and whose role is to hydrolyse the nucleotides into nucleosides. The main objective of this study was to characterize antibodies produced in the laboratory against ectonucleotidases and then to evaluate the expression of these enzymes in the umbilical cord and in the kidney by immunohistochemistry. The ectonucleotidases that we have selected for this work were the Nucleosides Triphosphates Diphosphohydrolases (NTPDases) of the cytoplasmic membrane, namely NTPDase1, -2, -3, -8, as well as ecto-5'-Nucleotidase (CD73). In addition, we have performed enzymatic histochemistry to verify the immunolocalization obtained and also to verify if these enzymes are active in the structures detected by immunohistochemistry.
Our results suggest that, in the umbilical cord, NTPDase1, -2, -3, and ecto-5'-Nucleotidase are located at the endothelium of the arteries and vein and in Wharton frost. In the kidney, NTPDase1 is located in the blood vessels, NTPDase2 is located on the apical surface of the renal tubules, NTPDase3 as well as NTPDase8 are located in the renal tubules. The current identification of these enzymes at the protein and activity level in the umbilical cord and in the kidney is a prerequisite with the goal to define the role of these enzymes and in extension of extracellular nucleotides in these tissues.
v
TABLE DES MATIERES
RÉSUMÉ ... iii
ABSTRACT ... iv
LISTE DES TABLEAUX ... vi
LISTE DES FIGURES ... vii
REMERCIEMENTS ... x
CHAPITRE I- INTRODUCTION... 1
I- LE CORDON OMBILICAL ... 1
1- DÉFINITION ... 1
2- COMPOSITION DU CORDON OMBILICAL ... 2
II- LES REINS ... 6
1- DÉFINITION DES REINS ... 6
2- STRUCTURE ET FONCTIONS DES REINS... 6
III- LES NUCLÉOTIDES EXTRACELLULAIRES ... 9
1- LES SOURCES ET MÉCANISMES DE RELÂCHEMENT DES NUCLÉOTIDES EXTRACELLULAIRES .... 10
2- HYDROLYSE DES NUCLÉOTIDES EXTRACELLULAIRES ... 11
CHAPITRE II- HYPOTHÈSES-OBJECTIFS ET METHODOLOGIE DU PROJET ... 17
IV- HYPOTHÈSES ET OBJECTIFS ... 17
V- MÉTHODOLOGIE DU PROJET ... 20
1- PRÉLÈVEMENT DES TISSUS ... 20
2- IMMUNOLOCALISATION ... 20
3- HISTOCHIMIE ENZYMATIQUE ... 21
CHAPITRE III- RESULTATS ET DISCUSSION ... 24
VI- RÉSULTATS... 24
PARTIE I- CARACTÉRISATION DES ANTICORPS ANTI-ECTONUCLÉOTIDASES PAR IMMUNOHISTOCHIMIE ... 24
PARTIE II- LOCALISATION DES ECTONUCLÉOTIDASES DANS LE CORDON OMBILICAL HUMAIN ... 30
PARTIE III- LOCALISATION DES ECTONUCLÉOTIDASES DANS LE REIN HUMAIN ... 36
PARTIE IV- ÉVALUATION DE L’ACTIVITÉ ENZYMATIQUE DES ECTONUCLÉOTIDASES DANS LE CORDON OMBILICAL HUMAIN ET DANS LE REIN HUMAIN PAR HISTOCHIMIE ENZYMATIQUE ... 40
VII- DISCUSSION ... 45
CHAPITRE IV- CONCLUSION ET PERSPECTIVES ... 52
1- CONCLUSION ... 52
2- PERSPECTIVES ... 52
vi
LISTE DES TABLEAUX
vii
LISTE DES FIGURES
Figure 1: Image d’un fœtus et son cordon ombilical au sein de l’organisme maternel ... 1
Figure 2: Vue du cordon ombilical ... 3
Figure 3: L’enroulement des artères ombilicales autour de la veine ombilicale ... 3
Figure 4: Structure du cordon ombilical ... 4
Figure 5: Les reins dans le système urinaire ... 6
Figure 6: Structure du rein ... 7
Figure 7: Circulation du sang dans les reins ... 9
Figure 8: Sources des nucléotides extracellulaires 22 ...11
Figure 9: Structures des NTPDases (Adapté de Zimmermann et al.2002) ...13
Figure 10: Immunolocalisation de la NTPDase1 dans le colon humain sur des coupes en paraffine et en cryosection avec l’anticorps hN1-8LI5 ...25
Figure 11: Immunolocalisation de la NTPDase2 en paraffine et en cryosection avec l’anticorps hN2-H9S ...26
Figure 12: Immunolocalisation de la NTPDase3 en paraffine et en cryosection avec l’anticorps hN3-2CI4 ...27
Figure 13: Immunolocalisation de la NTPDase1 dans l’artère du cordon ombilical humain avec l’anticorps hN1-9LI5 ...30
Figure 14: Localisation de la NTPDase2 dans la veine du cordon ombilical humain avec l’anticorps hN2-2LI5 ...31
Figure 15: Localisation de la NTPDase3 au niveau des artères dans le cordon ombilical humain avec l’anticorps hN3-1CI4 ...32
Figure 16: Localisation de la NTPDase8 au niveau des artères et la veine dans le cordon ombilical humain avec l’anticorps hN8-D7AS...33
Figure 17: Localisation de l’ecto-5’-nucléotidase au niveau des artères dans le cordon ombilical humain avec l’anticorps h5NT-2LI5 ...34
Figure 18: Localisation de l’ecto-5’-nucléotidase au niveau de la veine dans le cordon ombilical humain avec l’anticorps h5NT-2LI5 ...35
Figure 19: Localisation des ectonucléotidases dans le rein humain ...36
Figure 20: Localisation des ectonucléotidases dans le muscle lisse du rein humain ...37
Figure 21: Localisation de la NTPDase1 au niveau des capillaires entre les tubules rénaux ...38
Figure 22: Localisation de l’ecto-5’-nucléotidase dans le rein humain ...39
Figure 23: Hydrolyse de l’ATP, l’ADP et l’AMP dans le cordon ombilical ...40
Figure 24: Hydrolyse de l’ATP dans le rein humain ...41
Figure 25: Hydrolyse de l’ADP dans le rein humain ...42
viii
LISTE DES ABRÉVIATIONS ET DES SIGLES
Symboles Définition
ADN Acide Désoxyribonucléique ADP Adénosine 5'-Diphosphate AK Adénylate Kinase
AMP Adénosine 5'-Monophosphate
AR Antigen Retrieval ou Techniques de démasquage d’épitopes antigéniques ARN Acide Ribonucléique
ATP Adénosine 5'-Triphosphate BSA Albumine du Sérum de Bovin
NPP Nucléotide Pyrophosphatases/Phosphodiestérases NTPDases Nucléosides Triphosphates Diphosphohydrolases PAL Phosphatase alcaline
UDP Uridine 5'-Diphosphate UTP Uridine 5'-Triphosphate
ix Je remercie l’Éternel Dieu Tout Puissant de m’avoir accordé le souffle de vie pour faire ce
x
REMERCIEMENTS
Je tiens d’abord à remercier de tout mon cœur le Professeur Jean Sévigny de m’avoir accepté dans son équipe de recherche et de m'avoir permis de poursuivre mes études graduées dans son laboratoire. Avec vous j’ai eu une forte passion pour la recherche biomédicale. J'ai appris beaucoup de choses avec vous qui sont très nombreuses à citer. Merci infiniment pour vos disponibilités, vos conseils très précieux, votre amour pour les autres. Votre qualité de vous intéresser à la vie privée des autres fait que vous êtes pas seulement un simple directeur de recherche pour les étudiants, mais aussi un "Père". Tous ce qui passeront par votre main, verront pleines de choses qui seront comblées dans leur vie bien sur le plan académique, que privé. Que l’Éternel vous accorde une large longévité afin que vous voyiez et apprécier le fruit de tout ce que vous aviez semé en nous comme baguages académiques. Encore infiniment merci Professeur Jean Sévigny.
Je tiens à remercier tous mes Parents, surtout mon Papa pour votre soutien malgré la distance. Merci beaucoup pour les discussions sur la vie privée et des choses que vous m’appreniez aussi sur le plan académique, professionnel et spirituel. Ceci m’a toujours permis de surmonter des moments difficiles dans la vie. Que le Seigneur vous bénisse encore et vous accorde une large longévité. Bonne retraite à toi Papa Chéri!
Je tiens à remercier Madame Julie Pelletier. Merci beaucoup à vous pour votre disponibilité et tous vos multiples aides, conseils et suggestions pour les travaux de recherche. Je vous en suis vraiment reconnaissant.
Je tiens à remercier les collègues de notre équipe de recherche, Mabrouka, et Romuald pour l’harmonie, les discussions et les blagues au travail. Merci beaucoup.
xi Je tiens à remercier le Ministère de la Santé de la République du Bénin, pour m’avoir accordé la bourse pour mon séjour d’étude ici au Canada.
Je remercie beaucoup ma jumelle Antoinette Agonsanou, pour ton cœur chaleureux. Merci beaucoup.
Je tiens à remercier de tout mon cœur, tous mes amis, vous qui m’aviez aidé d’une manière ou d’une autre malgré vos multiples occupations. Que le Seigneur vous bénisse abondamment.
Je tiens à remercier tous mes Frères et sœurs pour vos différents soutiens moraux. Sans oublier l’épouse de mon Papa. Que le Seigneur vous bénisse abondamment.
Je tiens à remercier tous les Professeurs et collègues du Centre de Recherche du Centre Hospitalier Universitaire de l’Université Laval, et ceux rencontrés dans le cadre des activités académiques pour les discussions qui ont participé d’une manière à une autre, à ma culture académique.
1
CHAPITRE I- INTRODUCTION
I-
LE CORDON OMBILICAL
1- DÉFINITION
Le cordon ombilical est un organe présent chez l’organisme maternel et qui joint le fœtus ou l'embryon à la mère1. L'embryon et le fœtus sont nourris durant la gestation via les vaisseaux sanguins du cordon ombilical. Le cordon ombilical est composé de trois structures essentielles : La gelée de Wharton dans laquelle est noyée une veine et deux artères2. Le rôle principal de ses trois structures est la circulation entre le fœtus/embryon et le placenta au cours de la grossesse, assurant des fonctions vitales pour le fœtus/embryon.
Figure 1: Image d’un fœtus et son cordon ombilical au sein de l’organisme maternel
Image du catalogue de Médecine. (Lien : linternaute.com/science/biologie/diaporamas/06/odyssee-de-la-vie/10.shtml, consulté en avril 2017)
2
2- COMPOSITION DU CORDON OMBILICAL
A- LA GELÉE DE WHARTON
La gelée de Wharton identifiée par Thomas Wharton en 1656 est une structure décrite comme étant «protecteur des vaisseaux contre la compression2, 3, 4, 5. La gelée de Wharton renferme beaucoup de polysaccharides et très riche de collagène. Elle est riche en myofibroblastes4, 5. Le cordon ombilical est un organe important en source de cellules souches mésenchymateuses4. D’autres travaux effectués à ce sujet, décrivent la gelée de Wharton comme une structure renfermant beaucoup de protéines telle que l’actine α, la desmine, la fibronectin1, beaucoup de facteurs de transcriptions de la famille des GATAs, et de plusieurs autres protéines3.
B- LA VEINE OMBILICALE
La veine conduit le sang veineux du placenta vers le fœtus4, 5, 6
. La veine et les artères établissent une connexion entre l’organisme de l’embryon et le placenta maternel.
C- LES ARTÈRES OMBILICALES
Les deux artères ombilicales sont plus petites que la veine. Elles véhiculent le sang pauvre en oxygène et rempli de déchets du fœtus vers le placenta4, 5, 6. Les déchets issus du fœtus/embryon sortent par les artères et sont expulsés par le placenta maternel. Ceci est très important pour maintenir un environnement vital et pour le développement du fœtus/embryon.
3
Figure 2: Vue du cordon ombilical
Image du catalogue de Médecine. (Lien : linternaute.com/science/biologie/diaporamas/06/odyssee-de-la-vie/10.shtml, consulté en mars 2017)
Elles s’enroulent en forme spirale autour de la veine.
Figure 3: L’enroulement des artères ombilicales autour de la veine ombilicale
4 Les artères (coloration bleu) sont enroulées autour de la veine ombilicale (coloration rouge). Ses structures précédentes sont noyées dans la gelée de Wharton (coloration rose). La gelée de Wharton
et la veine ombilicale contiennent respectivement des cellules mésenchymateuses et des cellules souches de sang placentaires
De nombreux travaux suggèrent que les artères sont très pauvres en collagène comparativement à la gelée de Wharton qui est riche en collagène6
Figure 4: Structure du cordon ombilical
(1- Les deux artères ombilicales; 2- La veine ombilicale; 3- La gelée de Wharton) Santé gynécologique sur http://sweetpics.site/u/umbilical-cord-cross-section. Consulté en avril
2017)
D- ASPECT MÉDICAL DU CORDON
OMBILICAL
1- SECTION DU CORDON OMBILICAL
Dès la naissance du bébé, le cordon ombilical n'a plus d'utilité, car le bébé a désormais les potentialités de s’adapter à son nouvel environnement. Ainsi, il est nécessaire de faire le clampage du cordon ombilical. Le clampage est le fait de couper le cordon à la
1
1
2
3
5 naissance du bébé. En effet, on note deux types de clampages : Le clampage tardif et le clampage précoce. L'Organisation Mondiale de la Santé (OMS) dans son rapport de recommandation sur le clampage, préconise le clampage tardif (entre 1 et 5 min et après l'arrêt des pulsations du cordon) plutôt que précoce (dès le dégagement), à réserver dans des cas plus graves7. Selon l’OMS, le clampage tardif du cordon ombilical permet une circulation continue du sang du placenta vers le bébé pour une période estimé à trois minutes supplémentaires après la naissance contrairement à ce qui est observé au niveau du clampage légèrement plus tardif du cordon qui entraîne une augmentation de 50 % des réserves en fer à six mois chez le nourrisson. Chez toutes les femmes y compris les femmes séropositives au VIH et chez celles dont on ne connaît pas de façon précise le statut VIH, l’OMS recommande le clampage tardif du cordon ombilical7
.
2- UTILISATION DU CORDON OMBILICAL EN RECHERCHE
Le sang de cordon ombilical est une source énorme de cellules souches hématopoïétiques utilisées conjointement pour des transplantations dans le but de traiter des affections malignes et non malignes chez les enfants et les adultes8, 9, 10, 11. Les cellules souches dérivées du sang du cordon ombilical possèdent une capacité accrue pour la prolifération des cellules progénitrices et leur auto-renouvellement in vitro11. Le sang du cordon ombilical a longtemps été considéré comme un déchet biologique, mais aujourd’hui, le sang contenu dans le cordon ombilical est reconnu pour sa richesse en cellules souches selon Héma-Québec (Première et seule Banque publique de sang de cordon ombilical au Québec). Les cellules souches issues du sang de cordon ombilical sont utilisées pour remédier à plusieurs pathologies et permettent aussi d’améliorer les traitements offerts aux malades souffrant d’un mauvais fonctionnement de la moelle osseuse et des cellules souches impliquées dans la fabrication des globules rouges, des globules blancs et des plaquettes. Elles permettent aussi de traiter les maladies sanguines, malignes comme la leucémie, l’anémie et l’aplasie médullaire (Pathologie qui consiste à une absence de fonction de la moelle osseuse) 8, 9, 10, 11. Le sang de cordon ombilical est une option thérapeutique avantageuse pour les personnes pesant moins de 50 kg, nécessitant une greffe de cellules souches10, 11. En cas des transplantations urgentes, le sang du cordon
6 ombilical sert de transplantation allogénique chez les adolescents et les jeunes adultes atteints de malignité hématologique et qui n’ont pas de donneur de moelle osseuse9
.
II-
LES REINS
1- DÉFINITION DES REINS
Les reins sont deux organes qui sont situés au niveau de la colonne vertébrale, présentant une forme de haricot et qui permettent la synthèse de l’urine à partir de la filtration et de la purification des déchets du sang12, 13. Nous distinguons deux reins : le rein droit étant situé légèrement plus bas que le rein gauche12. Un rein humain mesure 12 centimètres sur 6 et fait 3 centimètres d'épaisseur. Dépendamment de l’âge de l’individu, cette taille varie d’un individu à un autre12
. Des pathologies touchant le rein telles que les insuffisances rénales chroniques peuvent entrainer une diminution de la taille du rein12.
Figure 5: Les reins dans le système urinaire
Institut National des cancer/anatomie du rein, consulté sur http://www.e-cancer.fr/Patients-et-proches/Les-cancers/Cancer-du-rein/Anatomie-du-rein en avril 2017
2- STRUCTURE ET FONCTIONS DES REINS
Le rein est un organe complexe doté de plusieurs fonctions de régulation. L’étude de la structure du rein nous permet de voir que le néphron à lui seul joue un rôle très important
7 au sein de cet organe. Le néphron est l'unité structurelle et fonctionnelle du rein14, 15. Chaque rein humain peut compter environ un million de néphrons constitués d'un glomérule qui permet la formation de l'urine, d'un tubule qui s’ouvre au niveau des calices. Le rein est composé d’une enveloppe externe que l’on nomme capsule qui protège le rein, les néphrons, les calices, le parenchyme rénal et le bassinet dans lequel est secrétée l'urine synthétisée12, 14, 15. Une fois synthétisée au niveau des néphrons, elle est acheminée dans les calices où elle va s'écouler dans le bassinet puis enfin dans l'uretère. Lorsque la vessie est pleine, le besoin d’uriner se fait sentir14, 15, 16.
Figure 6: Structure du rein
Institut National des cancer/anatomie du rein, consulté sur http://www.e-cancer.fr/Patients-et-proches/Les-cancers/Cancer-du-rein/Anatomie-du-rein en avril 2017
La multiplicité des fonctions exercées par le rein confère à celui-ci « une véritable usine d’épuration ». Les glomérules du rein ont des rôles très précis, telle est l’élimination exclusive des déchets véhiculés par le sang17. Ces déchets proviennent en majeure partie
8 des aliments que nous mangeons. Lorsque ces déchets sont accumulés dans le sang, cela deviennent plus souvent toxiques et par conséquent entrainent des pathologies rénales. Il est identifié que la vasopressine favorise une diminution de la réabsorption de l'eau rénale, suivie d’une osmolarité sérique plus faible17, 18
. Un dysfonctionnement des reins peut entrainer l’accumulation de ces déchets dans le sang. Notons cependant que ces déchets se présentent sous forme d’azotes et sont éliminés par les reins sous forme d’urée, l’acide urique obtenu après la dégradation ARN et ADN et enfin la créatinine qui sont les déchets provenant de l’action musculaire.
Les reins interviennent aussi dans la régulation de la pression artérielle en synthétisant des hormones comme la rénine et l’angiotensine qui joue un très grand rôle dans l’homéostasie cellulaire18, 19
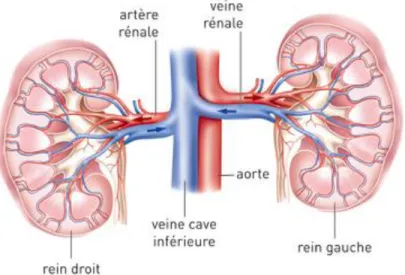
. Cela joue un rôle principal dans la dilatation des vaisseaux, dans la contraction des vaisseaux et dans la régulation de la pression artérielle. On note deux types d’angiotensines : l’angiotensine I et II. L’angiotensine I stimule la vasoconstriction des artérioles conduisant ainsi à l’élévation de la pression artérielle et le second rôle est la stimulation de la synthèse d'aldostérone intervenant dans l'élimination du sodium par le rein19. Le rein admet aussi un rôle central dans la circulation sanguine et lymphatique. En effet via les vaisseaux du rein, le sang arrive dans les artères, repart par la veine rénale qui est reliée à la veine cave inférieure.
9
Figure 7: Circulation du sang dans les reins
Institut National des cancer/anatomie du rein, consulté sur http://www.e-cancer.fr/Patients-et-proches/Les-cancers/Cancer-du-rein/Anatomie-du-rein en avril 2017
III- LES NUCLÉOTIDES EXTRACELLULAIRES
Les nucléotides comme l’ATP sont les sources de la génération de l’énergie cellulaire. Ils fournissent l'énergie nécessaire aux réactions chimiques du métabolisme, au potentiel électrochimique des molécules à travers la membrane, à la division cellulaire et au transport actif de molécules chimiques impliquées dans le maintien de l’architecture de la cellule en général et de la membrane cellulaire en particulier20, 21. En considérant leur rôle précieux dans le métabolisme cellulaire, de par leur rôle central dans le métabolisme intracellulaire, ils sont des molécules ubiquitaires20, 21. On sait maintenant que les nucléotides sont aussi des molécules de signalisation extracellulaire qui activent les récepteurs P222. Les récepteurs P2Y sont des récepteurs couplés aux protéines G que stimulent les nucléotides extracellulaires tels que l’ATP, l’ADP, l’UDP, l’UTP et l’UDP-glucose.
10
1- LES SOURCES ET MÉCANISMES DE RELÂCHEMENT
DES NUCLÉOTIDES EXTRACELLULAIRES
Dans le cytoplasme de la plupart des cellules, les concentrations de nucléotides mesurées sont de l'ordre du millimolaire (mM) 20, 21. Les nucléotides sont présents en grande quantité dans les fluides extracellulaires22. On note trois mécanismes à l’origine de la libération des nucléotides à l’extérieure des cellules : la nécrose cellulaire, l’exocytose de granules sécrétoires (vésicules synaptiques, granules denses des plaquettes), le flux sortant via des transporteurs membranaires22, 23. Il faut dire que la libération de ces nucléotides comme l’ATP, l’UTP, etc., à la surface de la cellule amène à une concentration suffisamment élevée dans le milieu qui entraine l’activation des récepteurs P222, 23. L’exocytose des nucléotides extracellulaires se déroule par exemple au cours des phénomènes de la transmission synaptique ou l’ATP colocalise dans les vésicules synaptiques avec d’autres neurotransmetteurs tels que l’acétylcholine et la noradrénaline qui sont libérés en même temps. Les granules des plaquettes qui fusionnent à la membrane plasmique et se déversent lors de l’agrégation plaquettaire en représente un autre exemple22, 23. Une stimulation des plaquettes par la thrombine, entraine des concentrations d’ADP et d’ATP de 100 μM à la surface des plaquettes22, 23
. Les nucléotides libérés via les transporteurs membranaires est une réponse à différentes stimulations mécaniques telles que : la force de cisaillement exercée par le flux sanguin sur l’endothélium vasculaire, le gonflement osmotique des cellules marquées par l’entrée importante des molécules telles que le Na+ le Ca 2+ dans la cellule, l’ouverture de la polarité membranaire favorisant une migration importante des ions de l’épithélium ciliaire et l’étirement des cellules épithéliales urinaires lors des mécanismes de la distension de la vessie22, 23.
11 Figure 8: Sources des nucléotides extracellulaires 22
2- HYDROLYSE DES NUCLÉOTIDES EXTRACELLULAIRES
Les nucléotides extracellulaires sont transformés rapidement par des enzymes présentes à la surface de nombreux types cellulaires et nommées ectonucléotidases22, 23, 24, 25, 26. Les ectonucléotidases régulent donc la concentration des nucléotides extracellulaires en les hydrolysant. Fait important, ces molécules sont aussi les agonistes des récepteurs P223, 24, 25, 26
. Les mécanismes de relâchement, d’interconversion des nucléotides à la surface des cellules, ainsi que le catabolisme permettent de contrôler les concentrations extracellulaires de l’ATP, l’ADP, l’UTP et l’UDP23, 24, 25, 26.
Les ectonucléotidases se subdivisent en 4 groupes et ayant chacun différents rôles22, 23. On a les nucléosides triphosphates diphosphohydrolases (NTPDases), les ectonucléotides pyophosphatases/phosphodiestérases (NPP), les phosphatases alcalines (PAL) et l’ecto-5'-nucléotidase22, 23.
12
A- LES NUCLÉOSIDES TRIPHOSPHATES
DIPHOSPHOHYDROLASES (NTPDases)
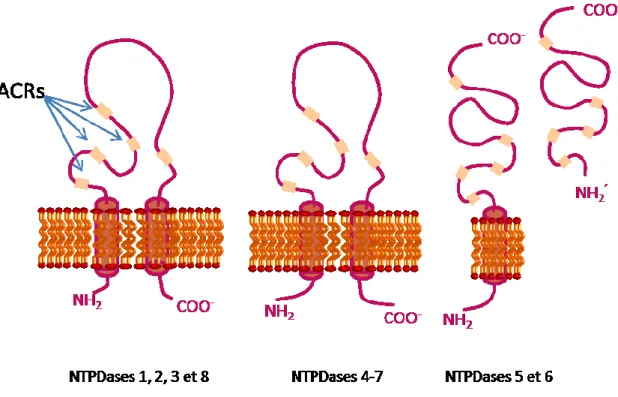
Les NTPDases sont responsables majoritairement de l'hydrolyse des nucléotides extracellulaires présents à la surface des cellules à pH physiologique22, 25,27. On distingue huit membres de cette famille qui sont regroupés en deux catégories selon leur localisation membranaire. La première catégorie regroupe les NTPDase1, -2, -3 -et 8 ancrées à la membrane plasmique, avec leur site actif faisant face au milieu extracellulaire22, 23, 24, 25, 26. Le deuxième groupe comprend les NTPDase4-7 et qui sont ancrées à la membrane des organelles cellulaires (lysosomes, appareil de Golgi et réticulum endoplasmique), avec leur site catalytique faisant face vers l’intérieur des organelles22, 23, 24, 25, 26.
Parmi les NTPDases, nous avons les NTPDase5 et -6, qui en plus de leur encrage à la membrane par un seul domaine transmembranaire, peuvent aussi être sécrétées sous forme soluble dans le milieu extracellulaire suite à un mécanisme de clivage protéolytique à l'extérieur de leur domaine transmembranaire22, 28.
13
Figure 9: Structures des NTPDases (Adapté de Zimmermann et al. 2002)
Légende : ACRs = Apyrase Conserved Regions
Les autres NTPDases (NTPDasel, -2, -3, -4, -7 et -8) ont une structure un peu différente de celle des NTPDase5 et -6. Les NTPDasel, -2, -3, -4, -7 et -8 ont deux domaines transmembranaires avec des extrémités N- et C-terminale intracellulaires22.
B-
LES ECTONUCLÉOTIDES
PYROPHOSPHATASES/PHOSPHODIESTÉRASES (NPP)
Les NPPs sont au nombre de sept membres (NPP1-7) qui sont des protéines à un seul passage transmembranaire et dont le site catalytique fait aussi face au milieu extracellulaire23, 29, 30, 31, 32. Ils ont pour rôles d’hydrolyser les liaisons pyrophosphates
14 et/ou phosphodiesters de différents substrats33, 34. La NPP1 et la NPP3 hydrolysent respectivement l’ATP et l’ADP en AMP32, 33. L’activité enzymatique des NPPs est inhibée par le dithiothréitol, le 2-mercapto-éthanol et le glycosaminoglycanes (GAG)/glycoaminoglycanes34. Aussi, l'activité hydrolytique des NPPs est optimale à pH basique35.
C- LES PHOSPHATASES ALCALINES (PAL)
Les phosphatases alcalines (PAL) sont des hydrolases qui ont pour rôle de cliver une liaison phosphoester par la libération d’un groupement hydroxyle et un ion phosphate36. Présent chez tous les organismes, elles sont capables d’hydrolyser la plupart des composés organiques disposant des groupements phosphorylés comme les nucléosides phosphates comme l’ATP, l’ADP et l’AMP32,36, 37. Elles ont une faible spécificité de substrat et peuvent cliver les groupes phosphate de nombreuses molécules telles que des nucléotides et des protéines. L’activité catalytique de la phosphatase alcaline est optimale en milieu basique (pH > 7)32,36, 37, 38, 39.
D- L’ecto-5'–NUCLÉOTIDASE
En fait nous avions ici deux catégories d’enzymes : Les 5'-nucléotidases cytosoliques et l'ecto-5'-nucléotidase/CD73 qui est membranaire. Ces enzymes hydrolysent spécifiquement les nucléosides monophosphates comme l'AMP en nucléosides et phosphates inorganiques40, 41, 42. L'ecto-5'-nucleotidase est aussi connue sous le nom de LVAP-2 (lymphocyte vascular adhesion protein 2) comme étant un marqueur de maturation des lymphocytes B/T40, 41, 42. De nature ubiquitaire, l'activité biochimique de cette ectoenzyme est optimale à pH neutre et est dépendante des ions divalents comme le Mg2+, le Ca2+, le Zn2+, le Mn2+et le Co2+. L'ecto-5'-nucléotidase possède une plus large spécificité de substrat envers les nucléosides 5'-monophosphate que les 5'-nucléotidases cytosoliques40, 41, 42. Plus récemment, notre laboratoire a démontré dans le foie que les NTPDase1,-2 et -8 co-localisent avec l’ecto-5’-nucléotidase/CD7340, 41, 42. Et par
15 conséquent on peut supposer que le complexe NTPDase-CD73 a un rôle important dans la régulation de la production d’adénosine hépatique40, 41, 42
17
CHAPITRE II- HYPOTH
ÈSES-OBJECTIFS ET METHODOLOGIE
DU PROJET
IV- HYPOTHÈSES ET OBJECTIFS
Le cordon ombilical fait partie des organes qui participent au maintien des fonctions biologiques et vitales du fœtus en général et assure les échanges entre l’organisme maternel et le fœtus en particulier. Il arrive que la femme en période de gestation développe des pathologies qui touchent le cordon ombilical tel que les maladies inflammatoires du cordon ombilical tel que l’inflammation de la gelée de Wharton qui est une cause des lésions inflammatoires aiguës du placenta comme la Chorioamniotite43, 1, 44, 45, 46
. En plus de nos jours, plus de 3 millions des nouveau-nés meurent chaque année dans le monde de maladies affectant le cordon ombilical en période néonatale comme le gros cordon, l’artère ombilicale unique, les reliquats embryonnaires, les fistules, les anomalies de paroi (cœlosomie, hernie ombilicale), les tumeurs (essentiellement le granulome), la chute retardée du cordon, et les infections locales (omphalite) ou à distance47.
Mon hypothèse est la suivante : « Les ectonucléotidases seraient présentes dans le cordon ombilical où ils participeraient à la modulation des fonctions du cordon ombilical par l'hydrolyse des nucléotides extracellulaires ».
Pour répondre à une partie de cette hypothèse, je me suis fixé les deux objectifs suivants : (1)- Tester d’abord mes anticorps humains anti-ectonucléotidases, pour connaître les conditions dans lesquelles ils fonctionnent et les utiliser pour répondre à l’objectif (2). (2)- Déterminer la localisation des ectonucléotidases dans le cordon ombilical humain.
Les informations sur l'expression cellulaire des ectoenzymes qui hydrolysent les nucléotides extracellulaires dans le rein sont incomplètes chez les mammifères et ne sont pas connues chez l’humain. En effet, l’équipe du Dr. Sévigny en collaboration avec le Dr. Kishore a localisé en 2005 la NTPDase1 et la NTPDase2 dans le rein chez la souris et ils ont identifié que la NTPDase1 est exprimée par les cellules musculaires lisses vasculaires
18 et l'endothélium dans les artères interlobulaires, les artérioles glomérulaires afférentes et les capillaires péritubulaires tandis que la NTPDase2 est exprimée dans la capsule de Bowman, les artérioles glomérulaires, l'adventice des vaisseaux sanguins et la paroi pelvienne.
Ceci nous amène à formuler l’hypothèse suivante : « Les ectonucléotidases seraient présentes dans le rein humain où ils participeraient à la modulation de ses fonctions par l'hydrolyse des nucléotides extracellulaires ».
Pour répondre à une partie de cette hypothèse, je me suis fixé l’objectif de déterminer la localisation des ectonucléotidases dans le rein chez l’homme.
20
V-
MÉTHODOLOGIE DU PROJET
1- PRÉLÈVEMENT DES TISSUS
Des biopsies de colon humain de trois (3) patients obtenues après chirurgie intestinale, de cordons ombilicaux de trois (3) femmes obtenues après accouchement et de rein humain obtenues de deux (2) patients ayant subi une chirurgie rénale ont été obtenues au CHU de Québec-Université Laval. Les tissus de colon humain sont congelés dans de l’OCT (Optimum Cutting Temperature) pour utilisation en cryosection et fixés dans du paraformaldehyde pour l’enrobage en paraffine. Les tissus du cordon ombilical et du rein sont congelés dans de l’OCT, c’est-à-dire mise en cryosection. Les coupes en cryosection sont conservées à -80oC et les coupes en paraffine sont conservés à 4oC jusqu’à leur utilisation pour l’immunohistologie. Pour chaque tissus, les tests sont répétés trois (3) fois sans compter les tests d’optimisation du signale des anticorps dans les tissus qui ont été fait à plusieurs reprises.
2- IMMUNOLOCALISATION
Les tissus en paraffine ont été sortis et couper au microtome et ceux en cryosection au cryostat. Des coupes histologiques de l’ordre de 6 microns (6 µm) ont été faites pour chaque tissu.
Les lames de cryosection ont été transférées dans une cuve contenant une solution de formaline-acétone (Seulement pour les coupes en cryosection) pour la fixation des antigènes pendant 2 minutes à 4oC et ensuite bloquées dans une solution de blocage, 5% BSA (Albumine du Sérum de Bovin), suivie d’une incubation avec l’anticorps primaire (1/500 pour les anticorps polyclonaux et 1 ug/mL pour les anticorps monoclonaux). Les anticorps ont été dilués avec une solution de 1% de BSA. Ensuite les lames sont incubées dans de H202 30% diluée à 3% dans du PBS 1X, pendant 10 minutes à température pièce, suivie de deux lavages successifs dans du PBS-T (5 minutes maximum par lavage). Ensuite le kit Avidin/Biotin est utilisé pour bloquer la biotine endogène. Après cette étape, les lames ont été incubées avec l’anticorps secondaire biotinylé. Après avoir passé à l’incubation avec le système avidine-biotine-peroxydase, la coloration des noyaux à
21 l’hématoxyline ont suivi la révélation à la Diaminobenzidine (DAB). La coloration des noyaux à l’hématoxyline permettant une détermination topographique du marquage. Enfin, les lames ont été visualisées au microscope pour l’analyse des résultats. Chacune des étapes de cette technique a été suivie de deux lavages successifs au PBS-T.
A l’exception de la cryosection, la fixation des antigènes dans les coupes de tissus incluses en paraffine n’est pas faite directement comme l’on fait sur les coupes de tissus en cryosection avec la solution d’acétone formaline. Les coupes en paraffine ont été d’abord déparaffinées par une solution de toluène suivie de l’incubation à des degrés décroissants d’éthanol (100o
C, 90oC, 70oC, 0oC). Cette étape a été suivie de l’utilisation des techniques de démasquage d’épitopes antigéniques. En effet, au niveau des coupes incluses en paraffine, les dérivés d’aldéhyde tel que la paraformaldehyde forment des liaisons protéiques qui masquent les sites antigéniques dans ces tissus, ce qui bloque ou rend faible la détection immunohistochimique de certaines protéines. Les techniques de démasquage d’épitopes antigéniques ont pour rôle de briser ces liaisons croisées des protéines, démasquant les épitopes des antigènes dans ces coupes de tissus et améliore l'intensité de coloration des anticorps48, 49.
Ainsi, pour les coupes en paraffine, les épitopes ont été démasqués par plusieurs techniques appelées «antigen retrieval» ou techniques de démasquage d’épitopes antigéniques. Ces techniques font appel à l’usage de certaines solutions dans le but de démasquer les sites antigéniques des antigènes. Les anticorps en paraffine ont été testés par plusieurs de ces solutions dont le Tampon Sodium Citrate (PH : 6), la Protéinase K et le Tampon TE (PH : 8). La Protéinase K a été diluée dans le Tampon TE (20 ug/ml). J’ai utilisé ses trois techniques pour tester nos anticorps en paraffine.
3- HISTOCHIMIE ENZYMATIQUE
Les coupes initialement conservées à -80oC, ont été transférées dans une cuve pour la fixation contenant une solution de formaline-acétone suivie d’une pré-incubation, avec une solution de Maléate Sucrose Buffer. Ensuite, on a effectué une incubation à PH = 7,4 avec les substrats ATP, ADP, et AMP suivie d’une révélation avec le sulfure d'ammonium. Les lames ont été ensuite visualisées au microscope pour l’analyse des
22 résultats. Pour évaluer l’activité enzymatique de la NTPDase1, -3, -8 j’ai utilisé les substrats ATP et ADP car la NTPDase1, -3, -8 hydrolyse l’ATP et l’ADP. Pour évaluer l’activité enzymatique de la NTPDase2, j’ai utilisé l’ATP. La NTPDase2 hydrolyse préférentiellement l’ATP. Pour évaluer l’activité enzymatique de l’ecto-5’-nucléotidase, j’ai utilisé l’AMP. Le lévamisole a été utilisé pour bloquer la phosphatase alcaline. Ainsi les substrats ne permettent pas de distinguer avec certitude l’identité de la NTPDase responsable de cette activité mais confirme la présence de ces enzymes dans les tissus.
24
CHAPITRE III- RESULTATS ET DISCUSSION
VI- RÉSULTATS
PARTIE I- CARACTÉRISATION DES ANTICORPS
ANTI-ECTONUCLÉOTIDASES PAR IMMUNOHISTOCHIMIE
Nous avons d’abord vérifié que les anticorps étaient spécifiques et qu’ils marquaient les bonnes structures en les utilisant par immunohistochimie dans un tissu où nous connaissons déjà la localisation des NTPDase1, -2, -3, -8 et de l’ecto-5’-Nucléotidase. Tous les anticorps que je vais présenter dans ce mémoire ont détecté les structures attendues dans le colon humain. En effet, dans ce tissu nous savons que la NTPDase1 est exprimée par les cellules musculaires lisses tant vasculaires que non-vasculaire ainsi que dans les cellules endothéliales des vaisseaux sanguins50, 51. La NTPDase2 est exprimée par les fibroblastes autour des vaisseaux sanguins et par les cellules gliales autour des neurones52, 53. La NTPDase3 est localisée dans les neurones54. La NTPDase8 est localisée au niveau des canalicules biliaires60 et au niveau de l’épithélium intestinal (donnés non encore publiés)60 et finalement l’ecto-5’-nucléotidase est localisée dans les muscles lisses et dans les vaisseaux sanguins55, 56, 57.
Les tests des anticorps anti-ectonucléotidases ont été effectués sur des coupes de colon humain en cryosection et en paraffine. Les prochaines figures montrent quelques-uns de ces résultats. J’ai testé ses anticorps dans plusieurs techniques de démasquage d’épitopes antigéniques. Les figures 10, 11 et 12 montrent respectivement un signal en brun correspondant au marquage par immunohistochimie avec des anticorps dirigés contre la NTPDase1, la NTPDase2, la NTPDase3 en paraffine avec plusieurs techniques de démasquage d’épitopes antigéniques.
25
Figure 10: Immunolocalisation de la NTPDase1 dans le colon humain sur des coupes en paraffine (a et b) et en cryosection (c) avec l’anticorps hN1-8LI5
L’anticorps hN1-8LI5, qui est dirigé contre la NTPDase1 humaine, montre un signal positif tel que l’on peut le visualiser ici par le dépôt brunâtre. En paraffine avec la technique de démasquage d’épitope antigénique Protéinase K, on note un signal au niveau du muscle lisse (longue flèche en a). L’anticorps hN1-8LI5 montre également un signal semblable sur des coupes en paraffine incubées dans le Tampon TE (b) et de la lamina propria (petite flèche en b). L’anticorpshN1-1CI5 montre un signal positif en cryosection et révèle la présence de la NTPDase1 au niveau des vaisseaux sanguins dans le colon humain (courte flèche en c).
26
Figure 11: Immunolocalisation de la NTPDase2 en paraffine (a) et en cryosection (b) avec l’anticorps hN2-H9S
L’anticorps hN2-H9S donne un signal positif brun avec la technique de démasquage d’épitope antigénique Tampon TE. Cet anticorps montre que la NTPDase2 est localisée au niveau des structures nerveuses (flèches). Ce signal est montré ici dans le colon humain tant dans les coupes en
27
Figure 12: Immunolocalisation de la NTPDase3 en paraffine (a) et en cryosection (b) avec l’anticorps hN3-2CI4
L’anticorps hN3-2CI4 donne un signal positif brun avec la technique de démasquage d’épitope antigénique Tampon Sodium Citrate. Cet anticorps montre que la NTPDase3 est localisée au niveau
des structures nerveuses (flèches) dans le colon humain tant dans les coupes en paraffine (a) qu’en cryosection (b).
Dans le groupe des anticorps anti-ectonucléotidases dirigés contre la NTPDase1, nous avons la hN1-9LI5 qui fonctionne aussi bien en paraffine avec la technique de démasquage d’épitope antigénique Tampon Citrate. Elle marque aussi les mêmes structures comme la hN1-8LI5 en paraffine et en cryosection. En cryosection, nous avons plusieurs anticorps contre la NTPDase1 qui fonctionnent en paraffine, tels que les anticorps polyclonaux hN1-8LI5, hN1-9LI5, hN1-1CI5, hN1-2CI5 et hN1-3CI5. Les deux premiers anticorps hN1-8LI5, hN1-9LI5 ont aussi fonctionné tant en paraffine qu’en cryosection, alors que les autres anticorps hN1-2CI5 et hN1-3CI5 ont fonctionné
28 seulement en cryosection. En ce qui concerne les anticorps anti-ectonucléotidases dirigés contre la NTPDase2 humaine qui sont testés, nous avons d’autres anticorps polyclonaux tels que la hN2-2LI5, hN2-2LI6 et hN2-3LI5 qui fonctionnent également très bien en paraffine et en cryosection. En plus de ces anticorps, nous avons les anticorps monoclonaux hN2-B2S, hN2-D5S et hN2-H10S, qui fonctionnent tous très bien en cryosection mais n’ont pas fonctionné en paraffine.
Dans le groupe des anticorps anti-ectonucléotidases dirigés contre la NTPDase3 humaine, nous avons les polyclonaux hN3-2CI4, hN3-2CI5 qui fonctionnent très bien en paraffine et en cryosection.
Nous n’avons aucun anticorps dirigé contre la NTPDase8 humaine qui fonctionne en paraffine. En cryosection, nous avons les anticorps monoclonaux dirigés contre la NTPDase8 humaine tels que la hN8-D7AS, la hN8-D7S, la hN8-B3S et la hN8-C5S qui fonctionnent très bien en cryosection, (Tableau 1).
Les anticorps dirigés contre l’ecto-5’nucléotidase humaine fonctionnent très bien en cryosection avec les polyclonals 1LI5, 2LI5, 3LI5, hN5NT-1CI5, hN5NT-2CI5, hN5NT-3CI5, (Tableau 1).
29 Légende :
Nom de l’anticorps : hN1-8LI5 (h : humaine; N1 :NTPDase1; 8L : Lapin No 8; I5 : Injection No 5) Nom de l’anticorps : hN1-3CI5 (h : humaine; N1 :NTPDase1; 3C : Cochon d’inde No 8; I5 : Injection
No 5)
Poly : Polyclonal. Mono : Monoclonal. PK : Protéinase K. Tp : Tampon.
Tableau 1: Liste des anticorps anti-ectonucléotidases humaines fonctionnant en paraffine (a) et en cryosection (b)
Le tableau 1 regroupe les anticorps anti-ectonucléotidases qui fonctionnent en paraffine et en cryosection et ayant un très fort signal. Noter qu’en plus des anticorps de la famille de la NTPDase8 et de l’ecto-5’-nucléotidase qui semblent fonctionner en paraffine et dont le signal est en cours d’optimisation, les anticorps ayant un faible signal ne sont pas figurés dans le Tableau 1. Notez que tous ces anticorps ont été produits dans notre laboratoire.
30
PARTIE II- LOCALISATION DES ECTONUCLÉOTIDASES DANS
LE CORDON OMBILICAL HUMAIN
Maintenant que nous connaissons les anticorps et les conditions permettant d’avoir un signal spécifique contre les ectonucléotidases nous avons procédé à la localisation de la NTPDase1, la NTPDase2, la NTPDase3, la NTPDase8, et de l’ecto-5’-nucléotidase dans le cordon ombilical par immunohistochimie. Les prochaines figures montrent l’expression de ces enzymes dans le cordon ombilical humain.
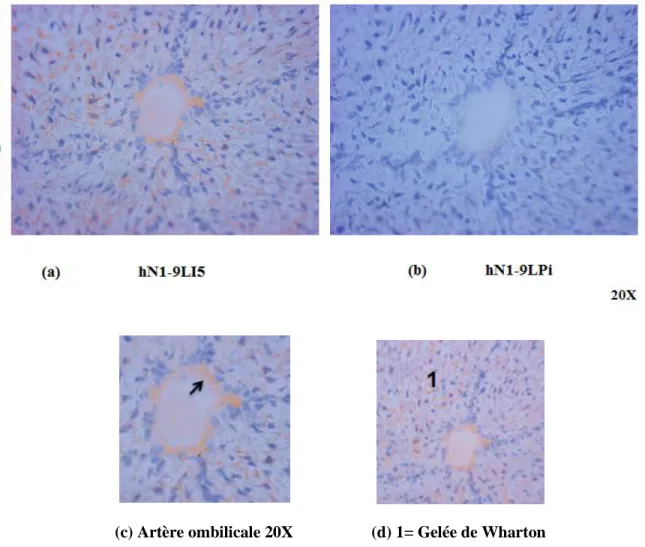
(c) Artère ombilicale 20X (d) 1= Gelée de Wharton
Figure 13: Immunolocalisation de la NTPDase1 dans l’artère du cordon ombilical humain avec l’anticorps hN1-9LI5 en cryosection
Cet anticorps montre un signal positif sur l’endothélium des artères (a, c) et dans la gelée de Wharton (d). (b) Contrôle Pré-immun hN1-9LPi.
31 On constate aussi l’expression de la NTPDase1 autour de la veine, au niveau de l’endothélium et au niveau de la gelée de Wharton dans le cordon ombilical. La NTPDase2 est localisée au niveau du muscle lisse dans le cordon ombilical et au niveau de l’endothélium de la veine (Figure 14).
Figure 14: Localisation de la NTPDase2 dans la veine du cordon ombilical humain avec l’anticorps hN2-2LI5 sur des coupes congelées
Expression de la NTPDase2 dans la veine du cordon ombilical humain avec l’anticorps hN2-2LI5 (a) avec le contrôle Pré-immun hN2-2LPi (b). Cet anticorps révèle que la NTPDase2 est localisée au
niveau de l’endothélium de la veine (flèche en c) et dans le muscle lisse (flèche en d).
La NTPDase3 est exprimée dans le cordon ombilical au niveau de l’endothélium autour des artères et autour de la veine. Elle est également exprimée sur la gelée de Wharton (Figure 15).
32
Figure 15: Localisation de la NTPDase3 au niveau des artères dans le cordon ombilical humain avec l’anticorps hN3-1CI4 sur des coupes congelées
Expression de la NTPDase3 au niveau de l’endothélium et dans la gelée de Wharton dans le cordon ombilical avec l’anticorps hN3-ICI4 (marquage brun au niveau de la figure a et b) avec le contrôle Pré-immun hN3-ICPi (a’, b’). Cet anticorps révèle que la NTPDase3 est localisée au niveau de l’endothélium de la veine (flèche en c) et au niveau de la gelée de Wharton (flèche en d).
La NTPDase8 semble être absente au niveau des artères et son expression est faible au niveau de la veine (Figure 16).
33
Figure 16: Localisation de la NTPDase8 au niveau des artères et la veine dans le cordon ombilical humain avec l’anticorps hN8-D7AS sur des coupes congelées
L’expression de la NTPDase8 est absente au niveau des artères (a, b), l’expression est faible au niveau de l’épithélium de la veine (c, e, f). Un contrôle IgG2b est montré en (d).
L’ecto-5’-nucléotidase est localisée partout dans le cordon ombilical. Elle est exprimée dans le muscle lisse, au niveau de l’endothélium des artères et de la veine. Son expression est forte au niveau de la veine. L’ecto-5’-nucléotidase est aussi exprimée au niveau de la gelée de Wharton. Comme dans d’autres tissus, son expression est à nouveau identifiée de nature ubiquitaire dans le cordon ombilical, (Figure 17 et 18).
34
Figure 17: Localisation de l’ecto-5’-nucléotidase au niveau des artères dans le cordon ombilical humain avec l’anticorps h5NT-2LI5 sur des coupes congelées
Expression de l’ecto-5’-nucléotidase au niveau de l’endothélium et dans la gelée de Wharton dans le cordon ombilical (a, b) avec le contrôle Pré-immun h5NT-2LPi (a’, b’). Cet anticorps révèle que l’ecto-5’-nucléotidase est localisée au niveau de l’endothélium des artères (flèche en c) et au niveau de
35
Figure 18: Localisation de l’ecto-5’-nucléotidase au niveau de la veine dans le cordon ombilical humain avec l’anticorps h5NT-2LI5 sur des coupes congelées
Expression de l’ecto-5’-nucléotidase au niveau de l’endothélium et dans la gelée de Wharton dans le cordon ombilical (a, a’, b) avec le contrôle Pré-immun h5NT-2LPi (b’). Cet anticorps révèle que l’ecto-5’-nucléotidase est localisée au niveau de l’endothélium de la veine (flèche en c) et au niveau de
36
PARTIE III- LOCALISATION DES ECTONUCLÉOTIDASES DANS
LE REIN HUMAIN
Comme nous avons fait précédemment dans le cordon ombilical, les mêmes anticorps ectonucléotidases sont testés dans le rein en plus d’autres anticorps anti-ectonucléotidases. Les figures suivantes montrent l’expression de ces enzymes dans le rein humain.
On note la localisation de la NTPDase1 au niveau des capillaires autour des tubules rénaux (Figure 19) et spécifiquement exprimée dans les muscles lisses, (Figure 20).
Figure 19: Localisation des ectonucléotidases dans le rein humain
L’anticorps hN1-1CI5 dirigé contre la NTPDase1, la hN2-H9S dirigée contre la NTPDase2, la hN3-1CI4 dirigée contre la NTPDase3 et la hN8-D7AS dirigée contre la NTPDase8 ont été utilisés sur des
coupes congelées. La NTPDase1 est localisée au niveau des capillaires autour des tubules rénaux (flèche en (a)). La NTPDase2 est localisée sur la face apicale des tubules rénaux (flèche en (b)). La NTPDase3 est localisée partout dans le rein. La NTPDase8 est localisée dans l’épithélium des tubules
(flèche en (d)).
La NTPDase2 est localisée dans certains tubules (Figure 19) et absente au niveau du muscle lisse dans le rein (Figure 20). Quant à la NTPDase3, elle est exprimée dans toutes les régions du rein (Figure 19) et elle semble être présente au niveau du muscle lisse du
37 rein (Figure 20). La NTPDase8 est exprimée seulement au niveau des tubules rénaux (Figure 19, 20). La NTPDase1 est aussi localisée au niveau des capillaires entre les tubules rénaux (Figure 21).
Figure 20: Localisation des ectonucléotidases dans le muscle lisse du rein humain
L’anticorps hN1-1CI5 dirigé contre la NTPDase1, la hN2-H9S dirigée contre la NTPDase2, la hN3-1CI4 dirigée contre la NTPDase3 et la hN8-D7AS dirigée contre la NTPDase8 ont été utilisés sur des
coupes congelées. La NTPDase1 est localisée au niveau du muscle lisse (flèche) alors que la NTPDase2 et la NTPDase8 sont absentes au niveau de cette structure. La NTPDase3 montre un
signal très faible sinon absent au niveau du muscle lisse du rein.
38
Figure 21: Localisation de la NTPDase1 au niveau des capillaires entre les tubules rénaux
L’anticorps hN1-1CI5 dirigé contre la NTPDase1 a été utilisé sur des coupes congelées avec le contrôle Pré-immun hN1-1CPi. Cet anticorps montre un signal positif au niveau des capillaires entre
les tubules rénaux tubulaires (flèche).
L’expression de l’ecto-5’-nucléotidase est localisée au niveau des capillaires des tubules rénaux et à l’intérieure des tubules rénaux (Figure 22).
39
Figure 22: Localisation de l’ecto-5’-nucléotidase dans le rein humain
L’anticorps h5NT-2LI5 dirigé contre l’ecto-5’-nucléotidase a été utilisé sur des coupes congelées avec le contrôle Pré-immun h5NT-2LPi. Cet anticorps montre un signal positif au niveau des capillaires
40
PARTIE IV- ÉVALUATION DE L’ACTIVITÉ ENZYMATIQUE DES
ECTONUCLÉOTIDASES
DANS
LE
CORDON
OMBILICAL
HUMAIN ET DANS LE REIN HUMAIN PAR HISTOCHIMIE
ENZYMATIQUE
Dans le cordon ombilical, on note une forte hydrolyse de l’ATP, de l’ADP et de l’AMP autour de la veine et des artères. Une forte hydrolyse de ces nucléotides est aussi observée dans la gelée de Wharton (Figure 23).
Figure 23: Hydrolyse de l’ATP, l’ADP et l’AMP dans le cordon ombilical
Activité enzymatique avec les substrats ATP, ADP, et AMPsur des coupes congelées. Dans les artères, on note une forte d’hydrolyse d’ATP (a), d’ADP (b) et d’AMP (c). Le contrôle avec une
41 Dans le rein on trouve une importante hydrolyse de l’ATP entre les tubules et dans/autour (de) certaines parties des glomérules. L’hydrolyse de l’ATP est aussi observée dans le rein humain au niveau des capillaires glomérulaires.(Figure 24). Cette activité est aussi observée au niveau des capillaires entre les tubules rénaux (Figure 24).
Figure 24: Hydrolyse de l’ATP dans le rein humain
Activité enzymatique avec l’ATP sur des coupes congelées. On note une hydrolyse d’ATP au niveau des capillaires entre les tubules rénaux (petite flèche). Cette activité est présente autour de certaines parties des glomérules et au niveau des capillaires entre les tubules rénaux (flèche avec tête pleine).
42 L’hydrolyse de l’ADP et de l’AMP sont aussi observées dans le rein humain (Figure 25 et 26). L’hydrolyse de l’ADP est observée autour des glomérules et des tubules dans certaines régions du rein. Dans d’autres régions, cette activité couvre une partie du pourtour des glomérules et des tubules. L’hydrolyse de l’ADP et de l’AMP sont aussi observées dans le rein humain au niveau des capillaires entre les tubules rénaux.
Figure 25: Hydrolyse de l’ADP dans le rein humain
Activité enzymatique avec l’ADP sur des coupes congelées. On note une hydrolyse d’ADP entre les tubules (flèche). Le contrôle avec une incubation sans nucléotide ne montre aucun marquage.
43
Figure 26: Hydrolyse de l’AMP dans le rein humain
Activité enzymatique avec l’AMP sur des coupes congelées. L’AMP est hydrolysée partout dans le rein humain. L’hydrolyse de l’AMP est présente à l’intérieur des tubules (longue flèche). On note également, une hydrolyse d’AMP au niveau des capillaires entre les tubules rénaux (petite flèche). Le
45
VII- DISCUSSION
La présente étude avait pour but de caractériser les anticorps anti-ectonucléotidases humains, dirigés contre les NTPDase1, NTPDase2, NTPDase3, NTPDase8 et l’ecto-5’-nucléotidase, par immunohistochimie afin de localiser ces enzymes dans le cordon ombilical humain et dans le rein humain. Les résultats de nos travaux montrent que plusieurs de nos anticorps anti-ectonucléotidases fonctionnent en paraffine et en cryosection. Nous avons obtenu des anticorps anti-ectonucléotidases qui fonctionnent en paraffine contre la NTPDase1, la NTPDase2 et la NTPDase3. La hN1-8LI5 est un anticorps polyclonal anti-NTPDase1 qui fonctionne très bien avec la proteinase K et avec le tampon TE. Le tampon TE est une solution avec laquelle on dilue la proteinase K. Ainsi, la proteinase K ne semble pas améliorer la détection pour cet anticorps en paraffine. Cette expérience est faite avec les autres anticorps qui ont été testés. Mais seul la hN1-8LI5, a fonctionné avec ses deux techniques de démasquage d’épitopes antigéniques à la fois avec un fort signal. La hN1-8LI5 marque la lamina propria et le muscle lisse dans le colon humain en paraffine avec les techniques de démasquage d’épitopes antigéniques Proteinase K et Tampon TE (Figure 10). Elle marque aussi les vaisseaux sanguins comme a été déjà démontré les travaux du Dre É. Lavoie et ses collaborateurs en 2010, qui ont localisé la NTPDase1 dans tous les vaisseaux sanguins dans le colon26,50. D’autres travaux ont également démontré la localisation de la NTPDase1 au niveau des vaisseaux52. L’anticorps hN2-H9S dirigé contre la NTPDase2 humaine fonctionne très bien en paraffine. Cet anticorps a récemment fait l’objet d’une publication dans notre laboratoire auquel j’ai participé et qui est actuellement sous presse53. La hN2-H9S marque les structures nerveuses dans le colon en paraffine et en cryosection (figure 11 de ce mémoire et [53]). Des travaux ont démontré la localisation de la NTPDase2 dans les structures nerveuses et leurs rôles importants dans le contrôle de l'activation des neurones périphériques 52, 54. L’expression de la NTPDase3 est localisée au niveau des structures nerveuses dans le colon humain (Figure 12). La NTPDase3 a été localisée dans les neurones54. En plus des polyclonaux hN3-2CI4, hN3-2CI5, nous avons aussi, les polyclonaux hN3-1CI4, hN3-1CI5, hN3-3CI4, hN3-3CI5 et les monoclonaux
46 hN3-B3S et hN3-H10S qui fonctionnent tous très bien en cryosection mais ne fonctionnent pas en paraffine (Tableau 1).
Pour la NTPDase8, nous n’avons pas eu un anticorps qui fonctionne en paraffine avec les techniques de démasquage d’épitopes antigéniques (Tableau1). Mais récemment, nous avons changé ses 3 techniques pour l’EDTA (une autre technique de démasquage d’épitopes antigéniques) et nous avons les anticorps dirigés contre la NTPDase8 humaine et de souris ainsi que les anticorps dirigés contre l’ecto-5’-nucléotidase humaine qui semblent donner un signal en paraffine. Nous sommes en train de travailler dans notre laboratoire pour optimiser le signal de ses anticorps.
Concernant l’expression des ectonucléotidases NTPDase1, -2, -3, -8 et de l’ecto-5’-nucléotidase dans le rein et dans le cordon ombilical, les résultats de nos travaux montrent que les ectonucléotidases sont présentes et sont actives dans le rein humain ainsi que dans le cordon ombilical humain. Ces enzymes exerceraient un rôle dans le maintien des fonctions biologiques de ces organes. Nous avions eu l’hydrolyse de l’ATP et de l’ADP dans le cordon ombilical. Ce qui permet de définir l’activité enzymatique de la NTPDase1, de la NTPDase2, et de la NTPDase3, dont l’expression est observée au niveau des artères ombilicales, de la veine ombilicale et sur la gelée de Wharton dans le cordon ombilical en figure 15 à 20. L’ecto-5’-nucléotidase a hydrolysé l’AMP. Ce qui suggère que cette enzyme catalyse dans le cordon ombilical l’étape d’hydrolyse des nucléosides 5’-monophosphates en nucléosides et phosphate (Pi) précisément au niveau des veines, des artères et au niveau de la gelée de Wharton. Ceci confirme que l’ecto-5’-nucléotidase est active dans ses structures du cordon ombilical humain. L’hydrolyse de l’ATP et de l’ADP observée dans le cordon ombilical en présence d’inhibiteurs de phosphatases alcalines nous permet de déduire que des NTPDases sont présentes au niveau de la veine et des artères ombilicales. Les résultats immunohistochimiques nous montrent la localisation de la NTPDase1, de la NTPDase2 de la NTPDase3 et de l’ecto-5’-nucléotidase au niveau de l’endothélium des artères et des veines. Ce qui nous amène
47 à définir avec l’activité enzymatique obtenue, que la NTPDase1, la NTPDase2, la NTPDase3 et l’ecto-5’-nucléotidase sont tous actives dans le cordon ombilical humain et seraient tous impliquées dans les rôles exercés par les cellules endothéliales au niveau de la veine et des artères. L’ecto-5’-nucléotidase est connue pour être impliquée dans l’hydrolyse de l'AMP en adénosine et régulant ainsi la perméabilité des vaisseaux sanguins55, 56. L’observation de l’expression de l’ecto-5’-nucléotidase dans le cordon ombilical humain, suggère que cet enzyme joue des rôles au sein de cet organe. À ce jour, aucune autre étude n’a vérifié l’activité de ces enzymes, ni de leur expression, ni du rôle de ses enzymes dans le cordon ombilical. Par contre, une libération d’ATP a été observée suite à la stimulation des cellules endothéliales de la veine ombilicale humaine57. Aucune liaison avec les ectonucléotidases n’ont été démontrées pour cette libération de l’ATP des cellules endothéliales de la veine ombilicale humaine. La présente identification des ectonucléotidases dans le cordon ombilical humain dans cette étude permettra de mieux comprendre et de pouvoir anticiper le rôle de ces enzymes au niveau du cordon ombilical humain.
L’hydrolyse de l’ATP, de l’ADP et de l’AMP est observée dans le rein humain. Ce qui nous permet de supposer que ces ectonucléotidases ont des rôles à jouer dans les fonctions rénales. L’hydrolyse de l’ATP que nous avons obtenue dans le rein pourrait nous amener à conclure sur la présence exclusive de la NTPDase2 puis que la NTPDase2 hydrolyse préférentiellement l’ATP et partiellement l’ADP. L’équipe du Dr Sévigny a déjà localisé en 2004, la NTPDase1 et la NTPDase2 dans le rein chez la souris et ils ont identifié que la NTPDase1 est exprimée par les cellules musculaires lisses vasculaires58. L’expression de la NTPDase1 est également observée au niveau de l'endothélium dans les artères interlobulaires, les artérioles glomérulaires afférentes et les capillaires péritubulaires tandis que la NTPDase2 est exprimée sur la capsule de Bowman, les artérioles glomérulaires, l'adventice des vaisseaux sanguins et la paroi pelvienne58. La NTPDase3 est rapportée comme étant localisée dans la plupart des segments de néphron au-delà du tubule proximal59. Aucune information n’est rapportée pour des études antérieures concernant la NTPDase8 qui est connue comme étant responsable de
48 l’hydrolyse des nucléotides extracellulaires dans les canalicules du foie de rat et de la souris60. Du moment où les ectonucléotidases sont impliqués dans plusieurs mécanismes connus tels que les inflammations, alors l’identification de son expression dans le rein, ouvre la voie à beaucoup d’études qui peuvent servir à éclaircir leurs implications physiologiques et physiopathologiques dans les maladies qui touchent cet organe. L’hydrolyse de l’AMP dans les tubules et autour des tubules comme observée avec l’expression de l’ecto-5’-nucléotidase localisée à l’intérieure des tubules et autour des tubules, suggère que l’ecto-5’-nucléotidase a un rôle à jouer dans ces structures. Les travaux de Weiru Zhang et ses collaborateurs en 2013, ont montré chez l’humain, qu’une augmentation de l'adénosine par l'ecto-5'-nucléotidase dans le rein, via les récepteurs A2B, provoque l'hypertension chronique61. Plusieurs rôles de l'ecto-5'-nucléotidase dans le rein ont été, démontrés par Cornelia Blume et son équipe en 2012. Ces travaux ont montré que l’adénosine obtenue suite à l’hydrolyse de l’AMP par l'ecto-5'-nucléotidase dans le rein est impliquée dans la rétroaction tubulo-glomérulaire et qu’elle est un important modulateur immunitaire62. En 2010, Rajakumar et son équipe a montré dans l'ischémie-reperfusion rénale légère que l'AMP peut jouer un rôle protecteur immédiat par l’adénosine qu’elle génère grâce à l’activité hydrolytique de l’ecto-5’-nucléotidase. Ce qui pourrait être utilisée dans le développement de nouvelles cibles thérapeutiques sur l’IRI rénal (kidney ischemia-reperfusion injury)63
. Les travaux de Cornelia Blume montre que chez les souris gloméruliennes cd73-/-, on note un nombre réduit de podocytes (cellules épithéliales différenciées et ramifiées composant le feuillet viscéral de la capsule de Bowman au niveau du glomérule rénal), de fenestrations endothéliales, des capillaires augmentés par glomérules, une endothéliose et une fibrose tubulaire accrue62.
Il faut noter qu’en dehors de notre laboratoire, d’autres laboratoires ont également élaboré des anticorps anti-ectonucléotidases humaines et qui sont utilisés en immunohistologie. Dans le groupe des anticorps contre la NTPDase1 humaine, il y a BU61 qui a été caractérisé par l’équipe du Dr Jean Sévigny en collaboration avec plusieurs équipes en 199664. Cet anticorps est commercialisé par l’entreprise Santa Cruz Biotechnology, inc65. Cet anticorps BU61 fonctionne bien en western blot, en immunoprécipitation, en immunofluorescence65 et en immunohistochimie72.