Comment un herbivore surabondant bénéficie des
ressources dépendantes, indépendantes ou découplées
de la pression de broutement
Thèse
Marie-Andrée Giroux
Doctorat en biologie
Philosophiae doctor (Ph.D.)
Québec, Canada
© Marie-Andrée Giroux, 2014
Résumé
La pression de broutement des herbivores peut induire des modifications majeures dans les communautés végétales qui peuvent réduire l’abondance des ressources alimentaires des herbivores et éventuellement mener à une rétroaction négative entre la taille des populations d’herbivores et leur taux de croissance. Une telle boucle de rétroaction peut s’opérer via une réduction de la masse corporelle à haute densité, suivie d’une réduction de la survie et de la reproduction. Malgré la détérioration persistante des communautés végétales de l’île d’Anticosti (Québec, Canada) suite à l’introduction d’une population de cerf de Virginie en absence de prédation naturelle, la taille de cette population est demeurée élevée durant les dernières décennies. Un des mécanismes suggérés pour expliquer cette situation est l’utilisation de ressources alimentaires qui seraient indépendantes ou découplées temporellement de la pression de broutement par le cerf. Deux des ressources consommées par le cerf à l’île d’Anticosti sont considérées indépendantes et découplées temporellement du broutement par le cerf, soit les algues échouées et le sapin baumier. Nous avons déterminé comment la masse corporelle du cerf à l’île d’Anticosti est influencée par les ressources alimentaires dépendantes, indépendantes et découplées temporellement du broutement. Premièrement, nous avons démontré expérimentalement que la densité de cerfs affectait négativement l’abondance estivale des herbacées et arbustes préférés, qui, en retour, affectait positivement la masse corporelle automnale. Deuxièmement, nous avons démontré que la contribution des algues au régime alimentaire en dehors de la saison hivernale était inférieure à 23% et qu’elle n’expliquait pas les variations de la masse corporelle automnale. La masse corporelle automnale augmentait plutôt avec la contribution de plantes d’habitat ouvert dans le régime alimentaire. Troisièmement, nous avons démontré que la perte de masse hivernale diminuait avec l’utilisation des sapinières durant un hiver rigoureux. Nos travaux permettent de mieux comprendre comment les herbivores surabondants peuvent bénéficier des ressources alimentaires affectées à divers degrés par le broutement. La prochaine étape serait de déterminer l’impact de ces différentes ressources sur le taux de croissance des populations surabondantes d’herbivores.
Abstract
Herbivores can induce major modifications to plant communities that can reduce forage abundance and lead to a retroaction between the size of herbivore populations and their growth rates. Such a feedback loop can be generated by a reduction in body mass at high herbivore density, followed by a reduction in survival and reproduction. Despite the lasting deterioration of plant communities on Anticosti Island (Québec, Canada) following the introduction of a white-tailed deer population in absence of natural predation, the size of this population has remained high during the last decades. One of the mechanisms suggested to explain this situation is the use of sources of forage independent or temporally uncoupled from deer browsing pressure. Two types of forage used by deer on Anticosti Island are considered independent or temporally uncoupled from deer browsing, namely drifted seaweed and balsam fir. We determined how body mass of deer on Anticosti Island is influenced by sources of forage that are dependent, independent and temporally uncoupled from deer browsing. First, we demonstrated experimentally that deer density had a negative effect on the summer abundance of preferred forbs and shrubs, which in turn had a positive effect on body mass in the fall. Second, we demonstrated that the contribution of seaweed to diet outside winter was smaller than 23% and did not explain variations in body mass in the fall. Fall body mass rather increased with the contribution of plants from open habitat to diet. Third, we demonstrated that overwinter mass loss decreased with the use of balsam fir stands during a harsh winter. Our findings provide a better understanding of how overabundant herbivores can benefit from sources of forage influenced to varying degrees by browsing. The next step would be to determine the impact of these different resources on the growth rate of overabundant herbivore populations.
Table des matières
RÉSUMÉ ... III ABSTRACT ... V TABLE DES MATIÈRES ... VII LISTE DES TABLEAUX ... XI LISTE DES FIGURES ... XIII LISTE DES ANNEXES ... XV REMERCIEMENTS ... XIX AVANT-PROPOS ... XXIII
CHAPITRE 1 ... 1
INTRODUCTION GÉNÉRALE ... 1
INTERACTIONS PLANTES-HERBIVORES ... 2
FACTEURS POUVANT INTERFÉRER DANS LA BOUCLE DE RÉTROACTION PLANTES-HERBIVORES ... 3
Délai dans la réponse dépendante de la densité chez les herbivores ... 3
Stochasticité environnementale ... 3
Utilisation de subsides écologiques ... 4
COMPOSANTES BIODÉMOGRAPHIQUES DES GRANDS HERBIVORES ... 5
Influence relative des composantes biodémographiques sur le taux de croissance ... 5
Survie juvénile et fertilité ... 6
Masse corporelle et contraintes saisonnières ... 7
PROBLÉMATIQUE À L’ÉTUDE ... 8
OBJECTIFS ET ORGANISATION DE LA THÈSE ... 10
CHAPITRE 2 ... 13
FORAGE-MEDIATED DENSITY AND CLIMATE EFFECTS ON BODY MASS IN A TEMPERATE HERBIVORE: A MECHANISTIC APPROACH ... 13
RÉSUMÉ ... 14 ABSTRACT ... 15 INTRODUCTION ... 16 METHODS ... 17 Study area ... 17 Experimental design ... 18 Winter climate ... 19 Statistical analyses ... 19 RESULTS ... 22 Experimental yearlings ... 22
Experimental vs. reference yearlings... 24
DISCUSSION ... 24
Density-dependence in life-history traits: a mechanistic approach ... 24
Carry-over effects of spring body mass ... 28
ACKNOWLEDGEMENTS ... 29
CHAPITRE 3 ... 31
CAN WE USE THE ISOTOPIC DIFFERENCE BETWEEN OPEN AND CLOSED HABITATS TO STUDY THE FORAGING BEHAVIOUR OF LARGE HERBIVORES? NEW EVIDENCE FROM A BOREAL STUDY SYSTEM ... 31
RÉSUMÉ ... 32
ABSTRACT ... 33
INTRODUCTION ... 34
MATERIAL AND METHODS ... 36
Study area ... 36
Study design ... 37
Food sources sampling ... 37
Deer sampling ... 38
Analyses ... 38
RESULTS ... 42
Objective 1: Isotopic difference between habitats ... 42
Objective 2: Isotopic difference in a single species between open and closed habitats ... 42
Objective 3: Variations in habitat-related deer diet ... 43
DISCUSSION ... 45
Canopy effect ... 45
Variations in the long term diet of deer ... 46
CONCLUSION ... 47
ACKNOWLEDGMENTS ... 47
CHAPITRE 4 ... 49
SHOULD A TEMPERATE UNGULATE LIVING IN A COASTAL ENVIRONMENT FORAGE IN OPEN, CLOSED OR LITTORAL HABITAT TO INCREASE ITS BODY MASS? NEW INSIGHTS FROM STABLE ISOTOPE ANALYSIS ... 49
RÉSUMÉ ... 50
ABSTRACT ... 51
INTRODUCTION ... 52
MATERIALS AND METHODS ... 54
Study area ... 54
Study design ... 55
Sampling of forage resources ... 56
Deer sampling ... 57
Deer density ... 57
Analyses ... 57
RESULTS ... 59
DISCUSSION ... 62
Habitat-specific use of forage resources ... 62
Increasing body mass by foraging more in open habitat... 63
ACKNOWLEDGMENTS ... 65
CHAPITRE 5 ... 67
A NEW WAY OF ASSESSING FORAGING BEHAVIOUR AT THE INDIVIDUAL LEVEL USING FAECES MARKING AND SATELLITE TELEMETRY ... 67
RÉSUMÉ ... 68 ABSTRACT ... 69 INTRODUCTION ... 70 METHODS ... 71 Ethics statement ... 71 Methodological approach ... 71 RESULTS ... 77 DISCUSSION ... 77
Linking foraging behaviour with life-history traits ... 78
Add-ons to existing methods for sampling foraging behaviour ... 79
Guidelines on the use of this technique in other study systems ... 80
ACKNOWLEDGMENTS ... 83
CHAPITRE 6 ... 85
WINTER SEVERITY MODIFIES THE RELATIONSHIPS BETWEEN HABITAT USE, DIET, AND WINTER MASS LOSS IN A TEMPERATE LARGE HERBIVORE ... 85
RÉSUMÉ ... 86
ABSTRACT ... 87
INTRODUCTION ... 88
METHODS ... 90
Study area ... 90
Climatic data in reference stations... 92
Capturing, marking and euthanizing individuals ... 93
Monitoring diet and habitat use ... 93
Statistical analyses ... 95
RESULTS ... 98
Comparison of climatic conditions among habitats and winters ... 98
Comparison of diet between winters... 100
Determinants of variations in the rate of body mass loss ... 102
DISCUSSION ... 103
Foraging behavior between and within years ... 103
Rate of body mass loss between and within winters ... 105
CONCLUSION ... 106
ACKNOWLEDGEMENTS ... 106
CHAPITRE 7 ... 109
CONCLUSION GÉNÉRALE ... 109
MÉCANISMES SOUS-JACENTS À LA BOUCLE DE RÉTROACTION PLANTES-HERBIVORES ... 111
FACTEURS POUVANT INTERFÉRER DANS LA BOUCLE DE RÉTROACTION PLANTES-HERBIVORES ... 112
GESTION DES RESSOURCES DE L’ÎLE D’ANTICOSTI ... 115 PERSPECTIVES DE RECHERCHE ... 117 BIBLIOGRAPHIE ... 119 ANNEXE 2.1 ... 139 ANNEXE 2.2 ... 141 ANNEXE 2.3 ... 161 ANNEXE 3.1 ... 165 ANNEXE 3.2 ... 169 ANNEXE 3.3 ... 179 ANNEXE 3.4 ... 181 ANNEXE 4.1 ... 183 ANNEXE 4.2 ... 185 ANNEXE 4.3 ... 189 ANNEXE 5.1 ... 191 ANNEXE 5.2 ... 193 ANNEXE 5.3 ... 195 ANNEXE 6.1 ... 197 ANNEXE 6.2 ... 223 ANNEXE 6.3 ... 225 ANNEXE 6.4 ... 229
Liste des tableaux
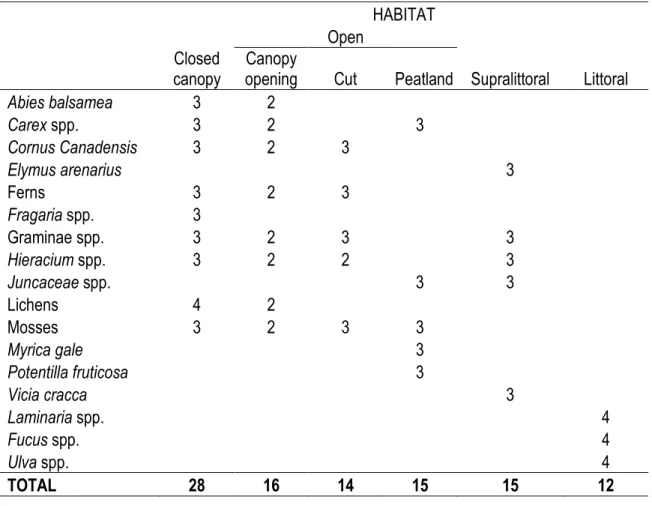
Table 3.1. Number of samples per taxa and habitat used to estimate the isotopic difference between
habitats (objective 1) and to average habitat-specific signatures in isotopic white-tailed deer diet reconstruction (objective 3) on Anticosti Island, Québec ... 39
Table 4.1. Model selection for generalized additive models fitted to determine whether fall dressed mass
(n = 151) varied with the contribution of resources from open, closed, ecotone supralittoral, and littoral habitat to diet, respectively, and taking into account the covariates presence of milk in the udder, index of deer density, age (cubic spline), and date of harvest (cubic spline). We report the value of the relative frequency of selection (π). Models are ranked by their π and the best model is shown in bold. ... 61
Liste des figures
Figure 1.1. Illustration des éléments clés de la problématique sous-jacente à la thèse. Cette
problématique est centrée sur les deux éléments suivants: 1- le rôle joué par les ressources alimentaires indépendantes (algues échouées) et découplées temporellement du broutement (sources de sapin baumier disponibles en hiver) dans le maintien de densités de cerfs élevées à l’île d’Anticosti (flèches noires) et 2- l’impact que l’utilisation de ces ressources alimentaires pourrait avoir sur les ressources alimentaires dépendantes du broutement (herbacées et arbustes préférés) via leur rôle potentiel dans le maintien de densités de cerfs élevées (flèches noires et grises). ... 10
Figure 1.2. Illustration de l’organisation de la thèse, structurée par les trois objectifs spécifiques et les
cinq chapitres rédigés sous forme d’articles scientifiques. Pour chacun des objectifs spécifiques, l’interaction entre les cerfs et les différentes ressources alimentaires est illustrée dans l’encadré. Dans cet encadré, les petites flèches grises indiquent le lien entre la masse ou la perte de masse corporelle et la densité de cerfs, élément qui n’est pas spécifiquement étudié dans le cadre de cette thèse. ... 11
Figure 2.1. Causal models showing the causal relationships explaining variations in the fall mass of
white-tailed deer. For every effect, we indicated the mean of the posterior distribution and its 95% credible interval estimated with untransformed continuous variables under a Bayesian hierarchical approach. The effects included in the two models used for inference are shown by black lines and text (first model), and by thick black lines and bold black text (second model). Dashed line indicates the effect included by default in the two models used for inference, but whose 95% credible intervals overlapped 0 (density); grey lines and text show the effects included in the global model, but not in the two models used for inference. ... 20
Figure 2.2. A- Influence of white-tailed deer density and winter NAO on forage abundance; B- Influence
of forage abundance and sex on fall dressed body mass of experimental yearlings; C- Influence of spring full body mass on fall dressed body mass of experimental yearlings; D- Fall dressed mass of experimental yearlings against experimental levels of the deer density treatment (the middle lines indicates medians, box limits are the first and third quartiles, whiskers indicate most extreme points ≤1.5 times the interquartile range, and dots indicate outliers); E- Influence of winter NAO on fall dressed body mass of experimental and reference yearlings. Symbols indicate raw data and predicted curves are drawn from parameters estimated with Bayesian hierarchical models. ... 24
Figure 3.1. Carbon and nitrogen isotopic signatures (δ13C and δ15N) of cartilage (corrected for
discrimination) in adult female white-tailed deer and of forage from different habitats on Anticosti Island, Canada. Squares represent the average signatures (± SD) of the four habitat types, whereas circles represent the signatures of individual deer. The sub-panel shaded in grey illustrates examples of habitat-specific Euclidean distances between an isotopic signature of deer and of resources from open (thick line) and closed habitats (dashed line). ... 41
Figure 3.2. Negative relationship between the habitat-specific proportion of forage in female white-tailed
deer diet on Anticosti Island (Canada) from 2002 to 2010 (A- open habitats, B- closed habitats) and habitat-specific Euclidean distances between isotopic signatures of deer and resources. Circles correspond to the average proportion of resources in deer diet, and error bars represent the 5–95th
percentiles of the distribution of proportions (estimated with SIAR). Predicted curves are drawn from the estimates of parameters averaged across models ran with each solution of the final set (see Methods). ... 44
Figure 4.1. Average proportions of forage from open, closed, ecotone supralittoral, and littoral habitat,
respectively, in long-term diet of female white-tailed deer on Anticosti Island (Canada) from 2002 to 2010 (n = 151). Dots correspond to the average proportions of the different forage types and error bars represent the range of the 5–95th percentiles of the distribution of proportions across all individuals
Figure 4.2. Positive effect of the proportion of forage from open habitat in the diet on the dressed body
mass of lactating and non-lactating female white-tailed deer on Anticosti Island (Canada) from fall 2002 to 2010 (n = 151). Dots correspond to the average proportion of resources in deer diet for both lactating and non-lactating females, and error bars represent the 5–95th percentiles of the distribution of
proportions per individual (estimated with SIAR). Predicted curves are drawn from the estimates of parameters averaged across the best fit model ran iteratively with each solution of the final set (see Methods). ... 62
Figure 5.1. Schematic representation of the four-step approach to sample foraging behaviour at the
individual level using faeces marking with food dyes and GPS telemetry. ... 72
Figure 5.2. Schematic representation of a trail as defined in our methodological approach. ... 74 Figure 5.3. Decision tree for determining whether the biomarking/telemetry technique could be used in a
given study system and which type of questions could be investigated. ... 81
Figure 6.1. Temporal trends in A- snow depth, and B- snow sinking depth from 1984 to 2010 in a
sampling station located in balsam fir stands on Anticosti Island (Québec, Canada). Each point represents the mean (±SD) of all measurements taken at 2-week intervals from the beginning of January to the end of March. We highlighted the values corresponding to the harsher (2008-09; white dot) and the milder (2009-10; grey dot) winters sampled in our study. ... 92
Figure 6.2. A- Snow depth, B- snow sinking depth, and C- wind chill temperatures for the main habitat
categories used by white-tailed deer females on Anticosti Island (Québec, Canada) during a harsher (2008-09) and a milder (2009-10) winter. For each variable, we show the predicted values (large black dots) and the 95% CIs obtained with linear models, along with the data used in the analyses (small white dots). Data used in analyses correspond either to the raw data collected on habitat-specific sampling stations (snow depth and snow sinking depth) or to the predicted values from the relationships between wind and temperature in habitat-specific sampling stations and at the airport (wind chill) (Annexe 6.1). Identical letters indicate that 95% CIs around estimated differences between habitats overlapped 0... 100
Figure 6.3. Proportion of forage resources in the diet of white-tailed deer females on Anticosti Island
(Québec, Canada) during the harsher (2008-09) and the milder (2009-10) winters. We show the predicted proportions (large black dots) and their 95% CIs obtained using a generalized mixed model (GLMM) with quasibinomial errors, along with the raw data (small white dots). Identical letters indicate that 95% CIs around estimated differences between type of forages overlapped 0. ... 101
Figure 6.4. Decrease in the rate of mass loss by female white-tailed deer on Anticosti Island (Québec,
Canada) with the proportion of locations in balsam fir stands in 2008-09, and with the proportion of herbaceous plants in diet in 2009-10. ... 103
Figure 7.1. A- Synthèse des résultats permettant de mieux comprendre l’influence de ressources
alimentaires ayant différentes dynamiques relatives au broutement et du climat sur le cerf de Virginie à l’île d’Anticosti. B- Approches utilisées pour obtenir ces résultats. ... 111
Liste des annexes
Annexe 2.1. Detailed results of the principal component analysis used to estimate the index of forage
abundance. ... 140
Annexe 2.2. Detailed description and results of path analyses and hierarchical Bayesian analyses. ... 142
Annexe 2.3. Detailed results of the comparison between experimental and reference yearlings (model selection and parameter estimates). ... 162
Annexe 3.1. Additional methodological details of isotopic analyses ... 166
Annexe 3.2. Assessing the habitat-specific isotopic difference between sectors, seasons, and years ... 170
Annexe 3.3. Summer diet of white-tailed deer on Anticosti Island. ... 180
Annexe 3.4. Model selection for linear models fitted to determine if δ13C and δ15N of Cornus canadensis varied between habitat, sector, and their interaction ... 182
Annexe 4.1. Map of the three sectors of the study area on Anticosti Island (Québec, Canada) ... 184
Annexe 4.2. Additional methodological details of isotopic analyses ... 186
Annexe 4.3. Additional results for the determinants of variations in fall dressed mass of deer ... 190
Annexe 5.1. Estimation of collar accuracy. ... 192
Annexe 5.2. Detailed results of model selection for mixed logistic regressions fitted to estimate the probability of sampling only dyed faeces for the different types of tracks (n = 96). ... 194
Annexe 5.3. Probability of recovery of dyed faeces by track type. Open circles represent raw data and closed squares represent predicted values with their 95% confidence intervals. ... 196
Annexe 6.1. Predicting hourly temperature and wind speed, as well as daily snow depth and snow sinking depth per habitat ... 198
Annexe 6.2. Microhistological techniques ... 224
Annexe 6.3. List of candidate models ... 226
« The ideal scientist thinks like a poet and only later works like a bookkeeper. »
Edward O. Wilson
- À William, Nico et mes parents qui remplissent ma vie d’un peu plus de poésie et de
Remerciements
Mon doctorat fût une aventure scientifique et humaine unique que je dois aux personnes exceptionnelles qui m’ont accompagnée tout au long de ce parcours. Je tiens ici à souligner leur contribution respective à cette aventure, contribution qui restera toujours très chère à mes yeux. En premier lieu, je tiens sincèrement à remercier mon directeur, Steeve Côté. Steeve, ce fût une expérience vraiment enrichissante que de travailler sous ta supervision et je me sens choyée d’avoir été exposée à ta façon rigoureuse et minutieuse de travailler. Je te remercie pour ton incroyable efficacité tout au long de mon doctorat, notamment quand ce fut le temps de réviser mes manuscrits. Je me souviendrai toujours du support unique que tu m’as offert durant cette période critique d’un doctorat. Mes plus sincères remerciements vont également à Jean-Pierre Tremblay, mon co-directeur. Jean-Pierre, tu as une capacité exceptionnelle à communiquer ta vision optimiste et positive de la vie. Cette dernière est contagieuse et je te remercie d’en contaminer tes étudiants! Tout au long de mon doctorat, j’ai eu la chance d’être entourée de plusieurs collaborateurs dont les forces respectives se sont complétées à merveille. Avant de travailler avec vous, je croyais en la force de la collaboration; chacun à votre tour, vous avez contribué à renforcer cette croyance et je vous en remercie. Christian Dussault, mille mercis pour ta contribution au projet d’hiver, ce fût un réel plaisir de travailler avec toi. Anouk Simard, je te remercie d’avoir partagé avec moi le résultat des innombrables heures que tu as passées à peser des cerfs dans les penderies à Anticosti. Éliane Valiquette, merci beaucoup pour ta gentillesse et ton professionnalisme dans le cadre de ton introduction à la recherche. Nicolas Lecomte, merci infiniment de m’avoir remplacé sur le terrain pendant que les soins maternels prenaient tout mon temps. Gilles Yoccoz, je me sens vraiment choyée d’avoir travaillé avec toi. Tu as sans aucun doute le don de transmettre ton savoir aux autres et ce, tout en ayant toujours une histoire intéressante à raconter entre deux équations.
Merci aux membres de l’équipe de la Chaire recherche industrielle CRSNG en aménagement intégré des ressources de l'île d'Anticosti pour votre agréable compagnie, votre dynamisme et votre aide précieuse. Un merci particulier à Sonia de Bellefeuille pour les mille et un services que tu m’as rendues tout au long de mon doctorat avec tant de gentillesse. Je suis également très reconnaissante aux professionnels de recherche, assistants de terrain et de laboratoire, techniciens, étudiants gradués et résidents de l’île d’Anticosti pour leur support et leur précieuse contribution aux
travaux de terrain et de laboratoire: Denis Duteau, Caroline Hins, Julien Hénault-Richard, Alexandre Veillette, Léon L’Italien, Jean-Philippe Harnois, Karim Ladhiri, Jean-François Adam, Marianne Bachand, Julien Arsenault-Hétu, Valérie Bélanger, Gaétan Laprise, Joël Malouin, Danièle Morin, Bianca Chénard, Arzhela Hemery, Marisa Sicilia, Gaston Reuchet, Catherine Bajzak, Sylvain Ménard, Mélanie Veilleux-Nolin, François Lebel, Stéphane Rivard, Bruno Baillargeon, Héli-Boréal. Pour ceux d’entre eux avec qui j’ai eu la chance de travailler sur le terrain, merci pour les bons moments passés avec vous à Anticosti.
Merci à Daniel Fortin, Gilles Gauthier et Stéphane Boudreau pour leur précieux conseils durant les réunions de mon comité de doctorat. I am grateful to Audun Stien, Fanie Pelletier and Stéphane Boudreau for taking time to evaluate my thesis and provide very constructive inputs.
Cassandra Cameron, mille mercis pour ta disponibilité et ton efficacité lors de la mise en page de ma thèse. Merci également à Nicolas Lecomte, Mael Le Corre et Julien Béguin pour avoir relue les parties françaises de ma thèse.
Merci aux équipes de recherche qui m’ont chaleureusement accueillie comme étudiante invitée durant mon doctorat. I am grateful to Gilles Yoccoz, Rolf Ims and the members of their research group for hosting me during my research internship in Tromsø. It was lovely to work in your research environment, and I really appreciated discussing science, skiing, and kids with you. Merci à Dominique Berteaux et Joël Bêty de m’avoir accueillie durant mon court passage à l’UQAR. Ce fût un réel plaisir de revenir passer du temps au sein de votre équipe de recherche. I am grateful to the team of the Wildlife Research Section of the Government of Nunavut for hosting me during my stay in Igloolik. Finalement, merci au groupe de recherche du Département de Biologie de l’Université de Moncton.
Je suis profondément reconnaissante à Nicolas Lecomte, William Lecomte et mes parents pour leur support et leur amour inconditionnel. Je suis tellement choyée d’avoir des proches aussi généreux. Nicolas, merci pour ton humilité, ton humanité et ta grande force de caractère qui ont été des sources d’inspiration au quotidien tout au long de mon doctorat. William, ta vivacité, ta curiosité, ta sérénité dans le mouvement, ton humour et ton désir profond d’aller de l’avant me touchent. Dolorès et Robert, mes parents, merci de m’avoir inculqué le sens du travail bien fait et la persévérance. Je vous remercie milles fois d’être si généreux de votre temps avec vos proches.
Je tiens également à souligner la contribution financière et logistique des organismes suivants: Conseil de Recherche en Sciences Naturelles et en Génie du Canada (subvention à S. D. Côté, bourses d’étude à M.-A. Giroux), Produits forestiers Anticosti, Université Laval, Ministère des Forêts, de la Faune et des Parcs, Société des Établissements de Plein Air du Québec, Centre d’Études Nordiques (bourses d’étude à M.-A. Giroux), et le Norwegian Research Council (bourses d’étude à M.-A. Giroux).
Avant-propos
La lecture de ma thèse vous offrira l’opportunité de mieux comprendre les interactions entre la population surabondante de cerfs de Virginie à l’île d’Anticosti et ses différentes ressources alimentaires, tout en vous faisant découvrir comment l’étude de cette population m’a permis de répondre à des questions fondamentales en écologie. Ma thèse débute par un chapitre d’introduction générale rédigé en français qui situe la problématique à l’étude par rapport aux connaissances scientifiques relatives aux interactions plantes-herbivores. Cette introduction présente également la structure de ma thèse, dont le cœur est formé de 5 chapitres rédigés en anglais sous forme d’articles scientifiques. Ces chapitres sont suivis d’une conclusion générale rédigée en français qui vise à résumer la contribution de ma thèse aux connaissances scientifiques et suggérer des pistes de recherche. L’ensemble des chapitres forme un tout, bien que les 5 chapitres rédigés sous forme d’articles scientifiques puissent être lus indépendamment.
Je suis l’auteure principale de tous les chapitres de ma thèse, mais j’ai eu la chance de bénéficier de la contribution de mon directeur Dr Steeve D. Côté (professeur titulaire au Département de biologie de l’Université Laval) et de mon co-directeur Dr Jean-Pierre Tremblay (professeur agrégé au Département de biologie de l’Université Laval) pour le financement du projet, l’élaboration des objectifs, la planification du projet, la rédaction et la révision des différents chapitres. J’ai également bénéficié de l’expertise de cinq collaborateurs, dont la contribution respective sera résumée ci-bas. Le chapitre 2 teste l’hypothèse que l’abondance des ressources alimentaires préférées du cerf de Virginie est le médiateur de l’effet de la densité et du climat sur la masse corporelle des cerfs. J’ai rédigé ce chapitre en collaboration avec Dr Jean-Pierre Tremblay, Dr Marie Anouk Simard (alors étudiante au doctorat en biologie à l’Université Laval), Dr Nigel Gilles Yoccoz (professeur titulaire au Département de biologie arctique et marine de l’Université de Tromsø) et Dr Steeve D. Côté. Ma contribution à cette publication parue dans Ecology en 2014 inclut la participation aux travaux de terrain en tant qu’assistante de recherche de premier cycle et d’étudiante au doctorat, la revue de littérature, la réalisation des analyses statistiques et la rédaction.
Le chapitre 3 permet de déterminer si la contribution des ressources alimentaires provenant d’habitats ouverts et fermés au régime alimentaire des herbivores en régions tempérées pouvait être quantifiée via l’analyse d’isotopes stables. Le chapitre 4 utilise l’approche décrite dans le chapitre 3
pour déterminer si la consommation d’algues par les cerfs de Virginie explique mieux les variations dans la masse corporelle automnale que la consommation de ressource alimentaires disponibles dans les habitats ouverts et fermés. J’ai rédigé le chapitre 3, qui sera soumis à PLOS ONE, en collaboration avec Éliane Valiquette (alors étudiante de 1er cycle en biologie à l’Université Laval), Dr
Jean-Pierre Tremblay et Dr Steeve D. Côté. J’ai rédigé le chapitre 4, qui sera soumis à Journal of
Mammalogy, en collaboration avec Dr Jean-Pierre Tremblay, Dr Marie Anouk Simard et Dr Steeve D.
Côté. Mon rôle couvre la planification de l’échantillonnage et la récolte des échantillons végétaux sur le terrain, l’encadrement d’Éliane Valiquette pour une partie des travaux de laboratoire et les analyses préliminaires dans le cadre du cours « Initiation à la recherche », les travaux en laboratoire, les analyses statistiques et la rédaction.
Le chapitre 5 vise à décrire le développement d’une technique d’échantillonnage du régime alimentaire individuel adaptée aux espèces évasives. Le chapitre 6 utilise cette technique pour déterminer l’influence de l’utilisation hivernale de sapin baumier sur la perte de masse corporelle hivernale des cerfs durant deux hivers aux conditions climatiques très contrastées. J’ai rédigé le chapitre 5, paru dans PLOS ONE en 2012, en collaboration avec Dr Christian Dussault (coordonnateur de la recherche sur la faune terrestre et l’avifaune au Ministère des Forêts, de la Faune et des Parcs du Québec), Dr Nicolas Lecomte (alors chercheur postdoctoral au Département de biologie marine et arctique à l’Université de Tromsø), Dr Jean-Pierre Tremblay et Dr Steeve D. Côté. J’ai rédigé le chapitre 6, qui sera soumis à Oecologia, avec les mêmes auteurs que le chapitre 5 à l’exception du Dr Nicolas Lecomte. Mon rôle couvre la planification de l’échantillonnage, la récolte des données sur le terrain, les travaux en laboratoire, les analyses statistiques et la rédaction.
Finalement, en plus des chapitres inclus dans ma thèse, je voudrais souligner l’existence d’un autre article et d’un autre manuscrit en lien direct avec ce projet. Premièrement, j’ai collaboré avec Pierre-Olivier Jean (étudiant au doctorat en biologie à l’Université de Sherbrooke), Dr Robert L. Bradley (professeur titulaire au Département de biologie de l’Université de Sherbrooke), Dr Jean-Pierre Tremblay et Dr Steeve D. Côté aux différentes étapes qui ont mené à la publication de l’article intitulé « Near infrared spectroscopy and fecal chemistry as predictors of the diet composition of white-tailed deer » paru dans Rangeland Ecology & Management en 2014. Deuxièmement, je collabore avec Dr Nicolas Courbin (chercheur postdoctoral en biologie à l’Université Laval), Dr Christian Dussault et Dr
Steeve D. Côté à la préparation d’un manuscrit visant à comparer la sélection hiérarchique de l’habitat par le cerf de Virginie durant deux hivers aux conditions climatiques très contrastées.
CHAPITRE 1
I
NTRODUCTION GÉNÉRALE
Mieux comprendre les patrons d’abondance des organismes vivants et les processus générant ces patrons représente une question centrale en écologie. Pour alimenter cette question, il est souvent nécessaire de prendre en compte les interactions trophiques entre l’organisme à l’étude et ses ressources alimentaires, prédateurs, parasites, et/ou compétiteurs. Dans les systèmes d’études simples, il peut être suffisant de se concentrer sur l’interaction trophique qui domine le réseau ou la chaîne trophique. Par exemple, dans le cas des populations d’herbivores sans prédateur, l’étude de l’interaction plantes-herbivores serait suffisante pour assurer une meilleure compréhension des patrons et des processus décrivant les variations d’abondance de ces populations.
Interactions plantes-herbivores
La pression de broutement des herbivores peut radicalement modifier la structure et la composition des communautés végétales et induire des effets négatifs sur la qualité de l’habitat des herbivores (Huntly 1991, Hobbs et al. 1996, Côté et al. 2004, Jefferies et al. 2006). Suite à la diminution de la disponibilité de leurs ressources alimentaires, une rétroaction négative entre la taille des populations d’herbivores et leur taux de croissance peut s’opérer via une réponse dépendante de la densité des composantes biodémographiques (Turchin 2003, Owen-Smith 2010), i.e. via une diminution de la survie, de la reproduction et de la croissance (Danchin et al. 2005). Un tel mécanisme défini la boucle de rétroaction plantes-herbivores. En théorie, cette boucle de rétroaction peut permettre de réguler les populations d’herbivores autour d’un point d’équilibre correspondant à un taux de croissance nul des populations d’herbivores et de leurs ressources alimentaires (MacNab 1985, Turchin 2003). Un tel mécanisme de régulation ne permet toutefois pas d’expliquer à lui seul la dynamique de plusieurs populations d’herbivores (Sæther 1997, Illius et O'Connor 1999, Owen-Smith 2010). Puisque ces populations peuvent avoir des impacts économiques et écologiques majeurs (Côté et al. 2004, Gordon et al. 2004, Côté 2005, Jefferies et al. 2006), il est nécessaire d’approfondir notre compréhension des mécanismes sous-jacents à l’interaction entre les herbivores et leurs ressources alimentaires. Pour ce faire, nous porterons une attention particulière aux facteurs pouvant interférer dans la boucle de rétroaction entre les populations de grands herbivores et leurs ressources alimentaires en absence de prédation naturelle.
Facteurs pouvant interférer dans la boucle de rétroaction
plantes-herbivores
Délai dans la réponse dépendante de la densité chez les herbivores
Un mécanisme suggéré pour expliquer pourquoi certaines populations d’herbivores ne sont pas régulées autour d’un point d’équilibre est le délai dans la réponse du taux de croissance de la population à la diminution de la disponibilité des ressources alimentaires en fonction de la densité (Clutton-Brock et al. 1997, Sæther 1997). Ce mécanisme, par exemple, expliquerait la différence entre la dynamique de la population de moutons de Soay d’Hirta (Ovis aries), qui fluctue de 60-70% à tous les 3-4 ans, et celle de la population relativement stable de cerfs élaphes (Cervus elaphus) de l’île de Rum (Clutton-Brock et al. 1997). D’après ces auteurs, un délai est introduit dans la réponse du taux de croissance de la population de moutons de Soay aux variations de densité car leur fécondité ne diminue pas en fonction de la densité. Par contre, le taux de croissance de la population de cerfs élaphes est plus sensible à la densité en raison de la réponse dépendante de la densité de la reproduction des femelles dans cette population. À forte densité, les femelles cerfs élaphes ne se reproduisent habituellement qu’une année sur deux et l’âge à la primiparité augmente.
Stochasticité environnementale
Les effets stochastiques, comme ceux induits par le climat, peuvent fortement influencer les variations démographiques des populations d’herbivores, et ainsi interférer dans la boucle de rétroaction plantes-herbivores (revu par Sæther 1997). Chez les grands herbivores, le taux de croissance ou les composantes démographiques peuvent par exemple être affectés par les variations stochastiques de la sévérité de l’hiver (Mysterud et al. 2001, Sæther et al. 2002, Tveraa et al. 2007), la quantité de précipitations annuelles (Owen-Smith 1990, Chamaillé-Jammes et al. 2008), les températures printanières (Loison et Langvatn 1998, Pettorelli et al. 2005) et la vitesse de débourrement de la végétation (Herfindal et al. 2006b, Pettorelli et al. 2007b). Chez plusieurs populations de grands herbivores, les facteurs stochastiques comme les variations climatiques interagissent avec la densité et peuvent même être amplifiés à forte densité (Portier et al. 1998, Aanes et al. 2000). Par exemple, dans une population d’éléphants d’Afrique (Loxodonta africana), une faible quantité de précipitations annuelles accentue la réponse négative du taux de croissance aux fortes densités d’éléphants (Chamaillé-Jammes et al. 2008). L’influence de la stochasticité environnementale sur la démographie des populations n’est pas un processus restreint aux grands
herbivores, mais s’exprime également dans plusieurs autres populations animales. Par exemple, dans l’archipel de Svalbard (Norvège), le taux de croissance des populations de campagnols de l’Europe de l’Est (Microtus levis), de lagopèdes de Svalbard (Lagopus muta hyperborea) et de rennes de Svalbard (Rangifer tarandus platyrhynchus) est synchronisé par les événements de redoux hivernaux (Hansen et al. 2013). De plus, ces mêmes auteurs ont observé que le taux de croissance de ces populations d’herbivores est corrélé à celui de la population de renard arctique (Vulpes
lagopus), mais avec un délai d’un an.
Utilisation de subsides écologiques
Un autre mécanisme pouvant interférer dans la boucle de rétroaction plantes-herbivores est l’utilisation de subsides écologiques. Le mécanisme de régulation rétroactif des populations d’herbivores via la compétition pour les ressources alimentaires implique que le taux de croissance ou la disponibilité de ces ressources diminue en fonction de la pression de broutement par les herbivores (Turchin 2003). Toutefois, la dynamique de certaines ressources alimentaires peut être indépendante ou découplée spatio-temporellement du la pression de consommation (Ellis et Swift 1988, Polis et Strong 1996, Tremblay et al. 2005). Ces ressources sont définies comme des subsides écologiques lorsqu’elles permettent de maintenir des densités de consommateurs plus élevées que ce qui serait supporté uniquement par les ressources dépendantes de la pression de consommation (Polis et al. 1997, Stapp et Polis 2003). Par exemple, suite à l’addition expérimentale de macroalgues dans un habitat littoral, la densité des populations d’amphipodes (Tethorchestia spp.) et de lézards (Anole brun; Anolis sagrei) ont augmenté (Spiller et al. 2010). Puisque l’importance des macroalgues dans le régime alimentaire de ces populations a augmenté en parallèle, cet exemple illustre comment l’utilisation de ressources alimentaires indépendantes de la pression de consommation peut supporter des populations de consommateurs à des densités plus élevées.
L’utilisation de subsides écologiques par les herbivores peut mener à la surabondance de ces populations et induire des effets catastrophiques sur les communautés végétales (Jefferies et al. 2006). Les impacts induits par les subsides sont causés par une dissociation spatiale ou temporelle entre la diminution de la disponibilité des ressources et la diminution du taux de croissance des herbivores (Polis et al. 1996). Une dissociation spatiale s’opère lorsqu’un consommateur utilise deux habitats qui contiennent respectivement des ressources indépendantes et dépendantes du broutement (Polis et al. 1997). Par exemple, l’augmentation soutenue de la disponibilité de subsides
agricoles sur les aires d’hivernage et les haltes migratoires a contribué à l’augmentation exponentielle de l’abondance des oies des neiges (Chen caerulescens) durant les dernières décennies (Abraham et al. 2005, Gauthier et al. 2005). L’explosion démographique d’une des deux sous-espèces d’oie des neiges, i.e. la petite oie des neiges (Chen caerulescens caerulescens), a entraîné des modifications majeures dans les communautés végétales présentes sur les aires de nidification de la Baie d’Hudson (Jefferies et al. 2006).
Outre la dissociation spatiale des subsides, une dissociation temporelle peut s’opérer lorsqu’il y a un délai entre la diminution de la disponibilité des ressources et la diminution du taux de croissance des herbivores. Un exemple de dissociation temporelle potentielle est le maintien de fortes densités d’ongulés dans des habitats où la pression d’herbivorie a des impacts négatifs sur la survie des semis d’espèces broutées (Tremblay et al. 2007), mais n’affecte pas à court terme la disponibilité de ces espèces sous forme de pluie de litière ou d’arbres renversés (Ditchkoff et Servello 1998, Miyaki et Kaji 2004, Tremblay et al. 2005). À court terme, le taux de croissance des ongulés pourrait être soutenu par la disponibilité de la pluie de litière ou d’arbres renversés et ce malgré les impacts négatifs du broutement sur la régénération de ces espèces. L’effet négatif du broutement sur la disponibilité de pluie de litière ou les arbres renversés pourrait s’exprimer à plus long terme, i.e. lorsque les peuplements d’espèces broutées auront disparu suite à l’absence de recrutement.
Ces exemples mettent en évidence que, pour mieux comprendre les mécanismes sous-jacents à l’interaction plantes-herbivores, il est primordial de prendre en considération la dynamique des ressources alimentaires relative au broutement, mais aussi l’influence des ressources alimentaires ayant différentes dynamiques relatives au broutement sur les herbivores. L’influence des ressources alimentaires sur les herbivores peut être quantifiée via les variations du taux de croissance d’une population, mais comme discuté ci-bas, également via les variations dans la survie, la reproduction et la croissance des individus.
Composantes biodémographiques des grands herbivores
Influence relative des composantes biodémographiques sur le taux de croissance Certaines composantes biodémographiques ont plus d’influence que d’autres sur la dynamique d’une population. L’influence relative des composantes biodémographiques sur le taux de croissance est fonction de leur variation temporelle et de leur élasticité, i.e. de l’effet d’un changement proportionnel
d’une composante sur le taux de croissance (Caswell 2000). La dynamique des populations de grands herbivores serait influencée principalement par la survie juvénile et la fécondité des jeunes femelles qui, malgré leur faible élasticité, sont très variables dans le temps (Gaillard et al. 1998, 2000, Raithel et al. 2007). À l’opposé, la survie des adultes aurait un faible impact sur le taux de croissance en raison de sa stabilité temporelle, malgré sa forte élasticité.
Survie juvénile et fertilité
La masse à la naissance serait critique pour la survie juvénile, surtout à forte densité (Clutton-Brock et al. 1982, Côté et Festa-Bianchet 2001a). La survie juvénile serait également influencée par les caractéristiques maternelles qui déterminent la qualité des soins maternels, soit la masse corporelle, l’âge et l’expérience de reproduction (Guinness et al. 1978, Cameron et al. 1993, Festa-Bianchet et al. 1995, Côté et Festa-Bianchet 2001b). Les conditions climatiques durant la gestation et après la naissance influenceraient aussi la survie juvénile (Skogland 1990, Gaillard et al. 1997, Portier et al. 1998, Forchhammer et al. 2001, Lomas et Bender 2007), notamment via leurs effets sur la disponibilité de nourriture de qualité (Pettorelli et al. 2007b). La survie des juvéniles durant leur premier hiver serait influencée par la condition corporelle à la fin de l’automne (Cook et al. 2004, Taillon et al. 2006), la sévérité de l’hiver (Leader-Williams 1980, Milner et al. 1999, Taillon et al. 2006) et la densité (Clutton-Brock et al. 1987, Skogland 1990, Portier et al. 1998, Milner et al. 1999). La masse à l’automne serait également critique dans la détermination de l’âge de primiparité des jeunes femelles ainsi que la probabilité de conception et/ou la taille de portée à l’ovulation des femelles adultes (Verme 1969, Skogland 1985, Albon et al. 1986, Langvatn et al. 1996, Cook et al. 2004, Pachkowski et al. 2013). La masse des femelles à l’automne serait influencée par la densité, le climat et la disponibilité de nourriture (Mysterud et al. 2001, Herfindal et al. 2006a).
Ces études mettent en évidence que les effets densité-dépendants et stochastiques agissent sur les composantes biodémographiques via des variations de la masse corporelle à différentes périodes du cycle annuel. Pour mieux comprendre l’interaction entre les populations d’herbivores et leurs ressources alimentaires, il est donc primordial d’identifier ce qui influence la masse corporelle durant les différentes périodes du cycle annuel.
Masse corporelle et contraintes saisonnières
Les grands herbivores des régions tempérées accumulent des réserves lipidiques et de la masse protéique durant l’été et l’automne, saisons durant lesquelles la nourriture de qualité est abondante et les coûts énergétiques liés à la thermorégulation et aux déplacements sont relativement faibles (Mitchell et al. 1976, Mautz 1978, Parker et al. 1993, Parker et al. 1999, Therrien et al. 2007). Ces caractéristiques saisonnières influencent l’efficacité d’approvisionnement (taux d’ingestion par rapport aux dépenses énergétiques) qui constitue un lien critique entre les ressources alimentaires et la masse corporelle (Parker et al. 1996). L’élevage des jeunes et la lactation se déroulent durant cette même période, ce qui génère des coûts énergétiques importants pour les femelles reproductrices (Oftedal 1985). Par exemple, l’allaitement accroît les besoins énergétiques des femelles de 27% chez le chevreuil (Capreolus capreolus) durant le premier mois (Mauget et al. 1997). Pour les mâles, une grande partie des réserves accumulées est utilisée à l’automne, soit durant la période énergétiquement coûteuse du rut (Yoccoz et al. 2002, Mysterud et al. 2005). L’hiver est caractérisé par une faible disponibilité de nourriture de qualité et des coûts énergétiques relativement élevés (Mautz 1978, Parker 2003). La faible disponibilité de nourriture de qualité est le résultat de l’absence de croissance végétale durant l’hiver, la qualité des plantes étant inversement reliée à leur stade de maturation (Van Soest 1982). L’accessibilité des ressources alimentaires est également réduite par la neige qui recouvre partiellement ou complètement la plupart des espèces d’herbacées et d’arbustes (Hovey et Harestad 1992, Nordengren et al. 2003, Visscher et al. 2006). Les coûts énergétiques élevés en hiver sont générés par les faibles températures qui poussent parfois les herbivores en dehors de leur zone de thermoneutralité (Parker et Robbins 1985) et par la présence de neige qui accroît les coûts de locomotion et d’acquisition de nourriture (Parker et al. 1984, Fancy et White 1985). Ces études ont démontré que l’augmentation des coûts associés à la présence de neige est corrélée à la profondeur et à la densité de neige. Puisque les besoins énergétiques hivernaux ne peuvent être comblés uniquement par les ressources alimentaires acquises durant l’hiver (Schwartz et al. 1988, Parker et al. 1996), l’accumulation de réserves et de masse protéique durant l’été et l’automne permet de tamponner la balance énergétique hivernale négative (Parker 2003). La masse corporelle est à son plus bas niveau à la fin de l’hiver (Cederlund 1991). La remontée des températures et le débourrement printanier de la végétation sont des événements clés qui contribuent à mettre fin à la période énergétiquement coûteuse qu’est l’hiver. Cette période demeure toutefois relativement coûteuse pour les femelles gestantes. En effet, la
gestation accroît les besoins énergétiques d’environ 45% chez les femelles de cerfs de Virginie (Odocoileus virginianus) et 90% de ces coûts sont assumés durant le dernier trimestre, soit durant la transition entre l’hiver et le printemps (Pekins et al. 1998).
En résumé, la masse corporelle des grands herbivores est considérée comme une variable charnière qui fait le lien entre la disponibilité ou l’utilisation des ressources alimentaires et les processus démographiques (Parker et al. 2009). La masse corporelle automnale est une variable clé expliquant les variations de la survie juvénile hivernale et de la fertilité des femelles, tandis que la masse à la fin de l’hiver serait déterminante via son influence sur la survie ainsi que sur la qualité et la quantité des soins maternels. La compréhension de l’influence des ressources alimentaires ayant différentes dynamiques relatives au broutement sur la masse corporelle durant ces deux périodes clés du cycle annuel est donc nécessaire pour mieux comprendre les mécanismes sous-jacents à l’interaction plantes-herbivores.
Problématique à l’étude
La problématique au cœur de cette thèse est celle de l’interaction entre une population surabondante de cerfs de Virginie et ses ressources alimentaires à l’île d’Anticosti (7,943 km2) dans le Golfe du
St-Laurent (49.06–49.95°N, 61.67–64.52°O). Après l’introduction d’environ 220 cerfs de Virginie en 1896-1897 sur cette île sans prédation naturelle, la population a rapidement augmenté pour atteindre des hautes densités environ 30 ans après son introduction (Potvin et al. 2003). Dans les dernières décennies, les densités de cerfs sont demeurées élevées (>20 cerfs/km2 en moyenne) malgré la
détérioration persistante des communautés végétales (Potvin et al. 2003, Tremblay et al. 2005) et de la condition corporelle des individus (Simard et al. 2008). Les hautes densités de cerfs ont entraîné l’éradication quasi-complète de la strate arbustive, à l’exception des arbustes éricacés dans les tourbières (Pellerin et al. 2006) et de la régénération d’épinettes blanches et noires (Picea glauca et
mariana Potvin et al. 2003). Plusieurs espèces d’arbustes décidus telles que l’érable à épis (Acer spicatum), le cornouiller stolonifère (Cornus stolonifera) et le noisetier à long bec (Corylus cornuta)
ont été pratiquement éliminées dû à la pression de broutement par le cerf (Pimlott 1963, Potvin et al. 2003). De plus, le brout décidu a presque disparu de la strate d’alimentation hivernale (Tremblay et al. 2005) et de nos jours, les cerfs sont contraints à s’alimenter principalement de conifères en hiver (Lefort et al. 2007). Bien que le sapin baumier soit normalement considéré comme une nourriture de famine utilisée en dernier recours pour éviter la mort par inanition (Ullrey et al. 1968, Crête et
Courtois 1997), cette ressource est fortement sélectionnée par les cerfs en hiver à l’île d’Anticosti (Sauvé et Côté 2007). En effet, le sapin baumier représente au moins 70% du régime alimentaire hivernal (Lefort et al. 2007), malgré sa faible disponibilité dans la strate d’alimentation hivernale (Tremblay et al. 2005, Massé et Côté 2012a). En fait, les trois principales sources de sapin baumier accessibles en hiver par le cerf sont les branches basses des arbres matures, les sapins renversés et les ramilles dans la pluie de litière (Tremblay et al. 2005, Massé et Côté 2012a). Une telle restriction de la disponibilité du sapin baumier est la conséquence de la pression chronique de broutement sur la régénération de sapin baumier, qui a également mené à la conversion des sapinières en pessières blanches et en landes forestières (Potvin et al. 2003).
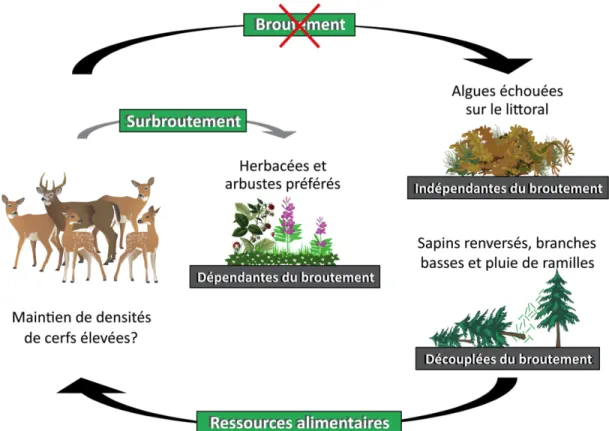
Pour expliquer pourquoi une rétroaction négative n’est apparemment pas observée entre la détérioration persistante des communautés végétales et la densité de la population, un des mécanismes suggéré est l’utilisation par le cerf de sources de nourriture qui seraient indépendantes ou découplées temporellement du broutement par le cerf (Tremblay et al. 2005). Premièrement, les algues échouées sur le littoral sont un exemple de ressources consommées par les cerfs à l’île d’Anticosti (observations des résidents et guides de chasse de l'île d'Anticosti; Huot 1982, Sénéchal 1989) que l’on considère indépendantes du broutement par le cerf. En effet, leur disponibilité sur le littoral dépend principalement de la productivité de l’écosystème marin, du cycle des marées, de la fréquence des tempêtes, de l’orientation de la côte et de l’exposition aux vents et aux vagues (Spiller et al. 2010, Lastra et al. 2014). Deuxièmement, les sources de sapin baumier disponibles en hiver sont un exemple de ressources alimentaires découplées temporellement du broutement. En effet, à court terme, les hautes densités de cerfs affectent négativement la régénération de sapin baumier (Tremblay et al. 2007), mais n’affectent pas l’abondance de branches basses d’arbres matures, de sapins renversés et de ramilles dans la pluie de litière disponibles dans les sapinières matures. À long terme par contre, les hautes densités de cerfs affectent fort probablement la disponibilité de ces trois sources de sapin baumier via la conversion des sapinières en pessières blanches et en landes forestières. L’utilisation de ressources indépendantes ou découplées temporellement du broutement pourrait permettre à cette population de cerfs, à court terme, de se maintenir à des densités plus élevées que ce qui serait supporté par les ressources dépendantes de la densité de cerfs comme les herbacées et les arbustes préférés (Tremblay et al. 2006). Les éléments clés de cette problématique sont illustrés à la Figure 1.1.
Figure 1.1. Illustration des éléments clés de la problématique sous-jacente à la thèse. Cette problématique est centrée sur les deux éléments suivants: 1- le rôle joué par les ressources alimentaires indépendantes (algues échouées) et découplées temporellement du broutement (sources de sapin baumier disponibles en hiver) dans le maintien de densités de cerfs élevées à l’île d’Anticosti (flèches noires) et 2- l’impact que l’utilisation de ces ressources alimentaires pourrait avoir sur les ressources alimentaires dépendantes du broutement (herbacées et arbustes préférés) via leur rôle potentiel dans le maintien de densités de cerfs élevées (flèches noires et grises).
Objectifs et organisation de la thèse
L’objectif général de cette thèse est de déterminer comment la masse corporelle d’un grand herbivore surabondant est influencée par les ressources alimentaires dépendantes, indépendantes, ou découplées temporellement du broutement. Le premier objectif spécifique est de déterminer si la masse corporelle des cerfs de 1,5 an répond aux variations de l’abondance des herbacées et des arbustes préférés, résultant potentiellement d’une réduction expérimentale de la densité de cerfs et des variations climatiques interannuelles. Le chapitre 2 vise à remplir ce premier objectif spécifique (Figure 1.2).
Figure 1.2. Illustration de l’organisation de la thèse, structurée par les trois objectifs spécifiques et
les cinq chapitres rédigés sous forme d’articles scientifiques. Pour chacun des objectifs spécifiques, l’interaction entre les cerfs et les différentes ressources alimentaires est illustrée dans l’encadré. Dans cet encadré, les petites flèches grises indiquent le lien entre la masse ou la perte de masse corporelle et la densité de cerfs, élément qui n’est pas spécifiquement étudié dans le cadre de cette thèse.
Le deuxième objectif spécifique de cette thèse est de déterminer si la consommation multi-annuelle d’algues par les femelles adultes explique mieux les variations de la masse corporelle automnale que la consommation multi-annuelle de ressources alimentaires disponibles dans les habitats ouverts et fermés. Avant de pouvoir répondre à la question écologique à la base de cet objectif spécifique, il était essentiel de déterminer si la contribution des ressources provenant d’habitats ouverts et fermés au régime alimentaire des herbivores en régions tempérées pouvait être quantifiée via l’analyse d’isotopes stables. Cette question technique est abordée dans le chapitre 3. Ensuite, le chapitre 4 utilise l’approche décrite dans le chapitre 3 pour répondre à la question écologique à la base du deuxième objectif spécifique (Figure 1.2).
Le troisième objectif spécifique de cette thèse est d’estimer l’influence de l’utilisation hivernale de sapin baumier sur la perte de masse corporelle hivernale des femelles adultes durant deux hivers
aux conditions climatiques très contrastées. Pour remplir cet objectif spécifique, il était impératif d’utiliser une technique d’échantillonnage du régime alimentaire individuel adaptée aux contraintes de notre système d’étude. Le chapitre 5 vise donc à décrire le développement d’une telle technique, basée sur la coloration des fèces et la télémétrie satellitaire. Ce chapitre représente une base essentielle pour la réalisation du chapitre 6, qui lui, vise à répondre à la question écologique du troisième objectif spécifique (Figure 1.2). Ainsi, ma thèse vise à mieux cerner le rôle joué par les ressources alimentaires caractérisées par différentes dynamiques relatives au broutement et le climat dans la dynamique de la masse corporelle d’un herbivore surabondant.
CHAPITRE 2
F
ORAGE
-
MEDIATED DENSITY AND CLIMATE EFFECTS ON
BODY MASS IN A TEMPERATE HERBIVORE
:
A MECHANISTIC
APPROACH
M
ARIE-A
NDRÉEG
IROUX,
J
EAN-P
IERRET
REMBLAY,
M
ARIEA
NOUKS
IMARD,
N
IGELG.
Y
OCCOZ,
ANDS
TEEVED.
C
ÔTÉ© Réimpression de l’article «Giroux, M.-A., Tremblay, J.-P., Simard, M. A., Yoccoz, N. G. & Côté, S. D. 2014. Forage-mediated density and climate effects on body condition in a temperate herbivore: a mechanistic approach. Ecology 95: 1332-1340» autorisée par Ecology, Ecological Society of America
Résumé
Puisqu’il est maintenant bien connu que le climat et la densité peuvent affecter simultanément la dynamique des populations d’herbivores, la recherche contemporaine vise plutôt à identifier les mécanismes sous-jacents aux effets du climat et de la densité sur les individus et les populations. Nous avons déterminé si la disponibilité des ressources alimentaires était le médiateur des effets de la densité et du climat sur la masse corporelle des cerfs de Virginie (Odocoileus virginianus) de 1,5 an. Pour ce faire, nous avons réduit expérimentalement la densité de cerfs à 7,5 et 15 cerfs/km2
durant huit saisons de croissance et utilisé des modèles de causalité (graphiques) hiérarchiques et des modèles hiérarchiques bayésiens pour évaluer les relations entre les différentes variables. L’abondance des ressources alimentaires préférées a diminué avec la densité de cerfs et a varié de façon quadratique (parabole positive) avec l’oscillation nord-atlantique hivernale, alors que la masse automnale des cerfs de 1,5 an a augmenté avec l’abondance des ressources alimentaires préférées et la masse printanière. La masse automnale n’était pas différente entre les densités de cerfs réduites expérimentalement, mais les cerfs expérimentaux étaient 30% plus lourds que ceux récoltés à densités ambiantes. Ainsi, l’abondance des ressources alimentaires préférées a modulé les effets de la densité et du climat sur la masse automnale, qui était aussi influencée par l’effet reporté de la masse printanière. Nos résultats améliorent notre capacité à prédire comment les grands herbivores des régions tempérées réagiront aux changements en cours des facteurs intrinsèques (e.g. densité de grands herbivores) et extrinsèques (e.g. climat).
Abstract
The interplay between density and climate in shaping the dynamics of herbivore populations is widely acknowledged, and current research is fueled by the identification of mechanisms underlying their effects on individuals and populations. We assessed whether forage availability mediated the effects of density and winter climate on body mass of white-tailed deer (Odocoileus virginianus) yearlings by experimentally reducing deer density to 7.5 and 15 deer/km² during eight growing seasons, and by
using causal (graphical) hierarchical models and Bayesian hierarchical modeling to assess relationships. The abundance of preferred forage decreased with deer density and varied quadratically (positive parabola) with winter NAO, whereas the fall mass of yearlings increased with forage abundance and spring mass. Fall mass did not differ between experimentally reduced deer densities, yet experimental yearlings were 30% heavier than yearlings harvested at ambient densities. Hence, forage abundance simultaneously mediated the effects of density and climate on fall body mass, which was also influenced by carry-over effects of spring body mass. Our findings increase our ability to anticipate how temperate large herbivores will respond to ongoing changes in intrinsic (e.g. large herbivore density) and extrinsic (e.g. climate) factors.
Introduction
The role of density-dependent processes in shaping the dynamics of populations has intrigued ecologists for decades (Nicholson 1933, Brook and Bradshaw 2006). While the existence of density-dependence is now widely recognized, current research is fueled by the identification of mechanisms underlying its effect on populations (Krebs 1995, Bonenfant et al. 2009). Density does not act directly on population abundance; it rather affects demographic processes (e.g. survival and reproduction) resulting from density-induced responses of life-history traits (Caswell 2000). Hence, one approach adopted to unravel the mechanisms of density-dependence has been to search for the effect of density on life-history traits of individuals and on demographic parameters of populations (Lebreton and Clobert 1991). Another complementary approach, the so-called mechanistic paradigm (Krebs 1995), aims to identify the trophic interactions, such as exploitative competition, that convey the effect of density on either individuals or populations.
In addition to intrinsic factors such as density, extrinsic factors such as climate can also shape life-history traits, and hence demographic processes (Birch 1957). In herbivores, climate contributes to drive variations in life-history traits in addition or in interaction with density (Sæther 1997, Coulson et al. 2000), and the effects of both factors can be mediated by the same trophic interactions. For instance, herbivore density or climate can drive the abundance of vegetation, which can in turn modulate the degree of food competition between herbivores and affect their life-history traits (e.g. density: Skogland 1985, climate: Forchhammer et al. 2001). In recent years, much attention has been devoted to the interplay between density and climate in shaping the life-history traits of herbivores (e.g. Sæther 1997, Coulson et al. 2000, Solberg et al. 2001, Patterson and Power 2002, Creel and Creel 2009, Garel et al. 2011). Fewer studies, however, investigated simultaneously the influence of forage, density and climate on life history traits of large herbivores (e.g. Skogland 1985, Herfindal et al. 2006a, Brown 2011, Dou et al. 2013). Among those studies, only a handful assessed the indirect role of foraging conditions in mediating the effects of climate on life history traits, using analytical methods than can quantify direct and indirect relationships (Mysterud et al. 2008, Nielsen et al. 2012, Texeira et al. 2012). Yet, to our knowledge, the hypothesis that both herbivore density and climate could simultaneously drive foraging conditions, which would in turn affect the life-history traits of herbivores, remains to be tested.
Our objective was to use a mechanistic approach to assess whether life-history traits could be simultaneously shaped by the indirect effects of density and climate mediated by forage availability in a large herbivore in absence of predation. As stipulated by the food hypothesis (Sinclair et al. 1985), density-induced variations in forage abundance is the most likely mechanism through which density can act on life-history traits in large herbivores not subject to predation (Bonenfant et al. 2009). Yet, as climate-induced variations in vegetation abundance are ubiquitous across the temperate biome (reviewed by Kreyling 2010), the potential dual action of density and climate on vegetation cannot be overlooked. We addressed this objective using a key life-history trait of large temperate herbivores, namely fall body mass of yearlings. Body mass is indeed considered a pivotal life-history trait through which density and climate can operate on demographic processes (reviewed by Gaillard et al. 2000, Bonenfant et al. 2009), and variations in life-history traits of juveniles or yearlings account for most of the variation in growth rates of large herbivore populations (Gaillard et al. 2000). As climate during the previous winter can contribute to shape variations in the fall body mass of temperate herbivores (e.g. Loison and Langvatn 1998, Post and Stenseth 1999), determining whether its effect is mediated by summer foraging conditions (Mysterud et al. 2008) or carry-over effects of spring body mass (Harrison et al. 2011) should also contribute to enhance our understanding of climate effects on temperate herbivores.
We examined the causal relationships underlying the effects of density and climate on body mass of yearling white-tailed deer (Odocoileus virginianus; hereafter “deer") by experimentally reducing deer density during the growing season, and by using causal (graphical) hierarchical models (Shipley 2009) and Bayesian hierarchical modeling (Gelman and Hill 2007) to unravel relationships. We hypothesized that: 1) density would negatively impact summer forage abundance, 2) which in turn would negatively affect fall mass; 3) fall mass would also decrease directly with density; and the influence of winter climate on fall mass would be mediated by both 4) summer forage abundance and 5) spring mass.
Methods
Study areaThe experiment was conducted from 2002 to 2009 on Anticosti Island in the Gulf of St. Lawrence, Québec, Canada (49.06–49.95°N, 61.67–64.52°W; 7943 km2). Mean air temperatures are 16 °C in
annually, while rainfall averages 61 cm (Environment Canada 1982). The forests of Anticosti Island belong to the boreal zone and are part of the eastern balsam fir-white birch bioclimatic region. At the end of the 19th century, a founding population of approximately 220 deer was introduced on the
island. In the absence of natural predators, the population peaked in the 1930s, has remained high thereafter (averaging ~20 deer/km2; Potvin et al. 2003) and has been exerting dramatic negative
impacts on plant communities (Tremblay et al. 2005). Experimental design
The randomized experimental design consisted of three replicated blocks. Each block was composed of two adjacent experimental units receiving one level of the density treatment: 7.5 deer/km2 and 15
deer/km2 (three deer in 40 ha and 20 ha enclosures, respectively; for a complete description of the
experiment, see Tremblay et al. 2006). From 2002 to 2009, we captured deer on Anticosti Island, relocated them in the experimental enclosures each spring (28 April to 12 June) and euthanized them in late fall (14 November to 13 December). We distributed mainly yearlings (11-12 months old at the time of capture; hereafter “experimental yearlings”) and adults among the experimental units (yearlings averaged 73% of deer introduced in the units). Enclosures were located in balsam fir-dominated forests partially cut (about 70%) in the early summer of 2001.
Forage abundance
We estimated forage abundance in 40 permanent sampling stations randomly established in each experimental unit in 2002. From 2002 to 2004, in 2007 and in 2009, we estimated percent of ground cover and height of preferred field-layer plant species (Chamerion angustifolium, Cornus canadensis,
Maianthemum canadense, Rubus idaeus, Rubus pubescens) in two one-m2 plots per station
randomly located in a 10 X 10 m quadrat (cluster sampling). We then predicted the aboveground dry biomass (g/m2) per species using regressions between percent plant cover, height and mass of dried
plants (see details in Tremblay et al. 2006). From 2002 to 2004, and from 2007 to 2009, we also
counted the number of saplings (>10 cm-200 cm) of preferred browse species (Betula papyrifera and Abies balsamea) in three 4-m2 circular plots established at 5-m intervals north of each sampling station. We obtained an index of forage abundance per experimental unit using the first axis of a principal component analysis (PCA) computed with the summed biomass of field-layer plants and the summed number of saplings of browse species. This index of forage abundance (hereafter “forage