CARACTÉRISATION STRUCTURALE ET FONCTIONNELLE DU RIBORÉGULATEUR SAM-I
par
Anne-Marie Dussault
mémoire présenté au Département de biologie en vue de l'obtention du grade de maître ès sciences (M.Sc.)
FACULTÉ DES SCIENCES UNIVERSITÉ DE SHERBROOKE
Library and Archives Canada Published Héritage Branch Bibliothèque et Archives Canada Direction du Patrimoine de l'édition 395 Wellington Street Ottawa ON K1A 0N4 Canada 395, rue Wellington Ottawa ON K1A 0N4 Canada
YourfHe Votre référence ISBN: 978-0-494-83648-4 Ourfile Notre référence ISBN: 978-0-494-83648-4
NOTICE:
The author has granted a
non-exclusive license allowing Library and Archives Canada to reproduce, publish, archive, preserve, conserve, communicate to the public by
télécommunication or on the Internet, loan, distrbute and sell theses
worldwide, for commercial or non-commercial purposes, in microform, paper, electronic and/or any other formats.
AVIS:
L'auteur a accordé une licence non exclusive permettant à la Bibliothèque et Archives Canada de reproduire, publier, archiver, sauvegarder, conserver, transmettre au public par télécommunication ou par l'Internet, prêter, distribuer et vendre des thèses partout dans le monde, à des fins commerciales ou autres, sur support microforme, papier, électronique et/ou autres formats.
The author retains copyright ownership and moral rights in this thesis. Neither the thesis nor substantial extracts from it may be printed or otherwise reproduced without the author*s permission.
L'auteur conserve la propriété du droit d'auteur et des droits moraux qui protégé cette thèse. Ni la thèse ni des extraits substantiels de celle-ci ne doivent être imprimés ou autrement reproduits sans son autorisation.
In compliance with the Canadian Privacy Act some supporting forms may have been removed from this thesis.
While these forms may be included in the document page count, their removal does not represent any loss of content from the thesis.
Conformément à la loi canadienne sur la protection de la vie privée, quelques formulaires secondaires ont été enlevés de cette thèse.
Bien que ces formulaires aient inclus dans la pagination, il n'y aura aucun contenu manquant.
Le 11 janvier 2012
le jury a accepté le mémoire de Madame Anne-Marie Dussault dans sa version finale.
Membres du jury
Professeur Daniel Lafontaine Directeur de recherche Département de biologie
Professeur Ryszard Brzezinski Membre
Département de biologie
Professeur Vincent Burrus Président rapporteur Département de biologie
SOMMAIRE
Les riborégulateurs sont un sujet de grand intérêt pour une partie de la communauté scientifique depuis leur découverte en 2002. Toutefois, comme ces structures retrouvées dans des ARNm ayant la capacité de réguler l'expression génétique en réponse à la liaison d'un petit métabolite sont présentées comme une cible antibiotique très prometteuse, les riborégulateurs intéressent maintenant les médias ainsi que la population en général. Malgré ce fort potentiel de cible antibiotique, il reste encore beaucoup d'interrogations à élucider avant de commercialiser des molécules ciblant les riborégulateurs afin de contrer une infection bactérienne chez un animal ou un humain. De cette manière, la recherche visant à mieux caractériser les riborégulateurs est d'une grande importance. L'étude présentée dans ce mémoire a donc comme objectif général de mieux caractériser le riborégulateur SAM-I d'un point de vue structural et fonctionnel.
Grâce à des techniques de transcription in vitro à cycle unique (single-round in vitro
transcription), d'acylation sélective du 2'-hydroxyle analysée par extension d'amorce
(SHAPE) et de transfert d'énergie par résonnance de fluorescence (FRET), il a été possible de mieux caractériser deux éléments structuraux importants pour le repliement et la fonction du riborégulateur SAM-I. En effet, il a été démontré que l'appartement du cœur ainsi que l'identité des nucléotides de la jonction 1/4, sont essentiels au bon fonctionnement du riborégulateur. De plus, des essais d'électrophorèse comparative sur gel (CGE) ont permis de caractériser la phase ondulatoire de la rotation de la tige PI du riborégulateur SAM-I ainsi que confirmer sa présence dans le cas d'un aptamère à 3 voies, c'est-à-dire qui ne possède pas de tige P4.
REMERCIEMENTS
D'emblée, je tiens à remercier le professeur Daniel Lafontaine pour plusieurs raisons. Tout d'abord pour m'avoir acceptée avec autant de chaleur dans son laboratoire. Ensuite, pour avoir cru énormément en moi et en mon potentiel, et ce, même lorsque j'en doutais moi-même. Finalement, pour avoir compris le besoin que j'ai éprouvé à rédiger plus tôt que prévu. Sincèrement merci Daniel, tu ne peux pas t'imaginer à quel point cela a compté pour moi. Je voudrais aussi remercier mes conseillers, les professeurs Vincent Burrus et Ryszard Brzezinski qui ont été de très bons conseils, tant au point de vue académique que de la recherche. Merci d'avoir toujours démontré un grand intérêt pour mon projet.
Je tiens aussi à remercier les membres du laboratoire qui m'ont accueillie à mon arrivée, soit Jérôme, Jean-François, Simon, Renaud et Benoit. Merci aussi à mes collègues passés ou présents, soit Audrey, Marie-Pier, Marc-André, Laurène, Patrick, Frédéric, Maxime, Sébastien, Simon, Cindy, Anne-Marie et tous les autres qui ont été là pour moi et qui m'ont aidée et supportée. Merci Audrey pour tes conseils scientifiques, mais aussi pour ton amitié, ta grande générosité et ton soutien de tous les jours, surtout lorsque j'avais le moral à plat. Merci aussi d'avoir investi du temps pour lire et commenter mon mémoire. Bien que je les ai déjà remerciés précédemment^ je tiens à dire merci encore une fois à Audrey et Frédéric participer à ce projet. Sans les résultats que vous m'avez fournis, l'histoire serait beaucoup moins intéressante. Merci aussi de m'avoir permis d'utiliser ces résultats dans mon mémoire.
Finalement, merci à ma famille et à mon amoureux pour absolument tout. Jean-Michel, sans qui je n'aurais sans doute pas complété mes études graduées. Je t'aime.
TABLE DES MATIÈRES
SOMMAIRE ii
REMERCIEMENTS iii
TABLE DES MATIÈRES iv
LISTE DES ABRÉVIATIONS viii
LISTE DES TABLEAUX xii
LISTE DES FIGURES xiv
INTRODUCTION 1
A. Les avancées de la biologie moléculaire 1
B. L'expression génétique et métabolisme 2
B.l. Régulation de l'expression génétique et du métabolisme chez les bactéries 3 B.l. 1. De l'ADN à l'ARNm : régulation de la transcription 5 B.l.2. De l'ARNm à la protéine : régulation de la traduction 7 B.1.3. De la protéine au métabolite cellulaire : régulation des activités enzymatiques
7...
C. Les riborégulateurs 9
C.l. Découverte des riborégulateurs 9
C.2. Définition du concept de riborégulateur 10
C.3. Mécanismes de régulation utilisés par les riborégulateurs 12
C.3.1. Mécanisme de régulation transcriptionnelle 13
C.3.3. Épissage alternatif 14 C.3.4. Régulation antisens 15 C.3.5. Ribozyme 15 C.4. Classes de riborégulateurs 16 D. Riborégulateur SAM 21 D.l. 5-adénosylméthionine et 5-adénosylhomocystéine 21 D.2. Riborégulateurs SAM 22 D.2.1. Riborégulateur SAM-1 23 D.2.2. Riborégulateur SAM-II 26
D.2.3. Riborégulateur SAM-III ou SAMMk 27
D.2.4. Riborégulateur SAM-IV 28
D.2.5. Riborégulateur SAM-V 29
D.2.6. Riborégulateur SAM/SAH 30
D.2.6. Les riborégulateurs SAM en résumé 30
E. Riborégulateurs SAM-I chez Bacillus subtilis 31
E.l. Détails concernant la structure 32
H. 1.1. Les tiges : PI, P2, P3 et P4 32
E.l.2. Le cœur de l'aptamère 35
E.2. Repliement en deux étapes 36
CHAPITRE 1 - ÉTUDE DE LA JONCTION VA ET DE L'APPAREMMENT DU COEUR ..38
1.1. INTRODUCTION 38
1.2. MATÉRIEL ET MÉTHODES 42
1.2.1. Transcription in vitro à cycle unique (single-round in vitro transcription) 42
1.2.2. Transcription des ARN 43
1.2.3. Marquage radioactif d'ADN en 5' 44
1.2.4. Acylation sélective du 2'-hydroxyle analysée par extension d'amorce (SHAPE) 44 1.2.5. Préparation des complexes d'ARN pour les essais de fluorescence 45
1.2.6. Fluorescence (FRET) 47
1.3. RÉSULTATS 48
1.3.1. L'appariement du cœur est important pour la régulation induite par le
riborégulateur SAM-1 48
1.3.2. L'absence de l'appariement du cœur n'affecte pas la formation des éléments
structuraux formant l'aptamère du riborégulateur 52
1.3.3. L'appariement du cœur et la jonction 1/4 ne sont pas essentiels à la
préorganisation du site de liaison du ligand 55
1.3.4. Importance de l'appariement du cœur pour l'adoption de l'état natif. 57 1.3.5. Importance de l'identité des nucléotides de la jonction 1/4 pour l'adoption de
l'état natif. 60
CHAPITRE 2 - ÉTUDE DE LA ROTATION DE LA TIGE PI 71
2.1 INTRODUCTION 71
2.2 MATÉRIEL ET MÉTHODES 72
2.2.1 Transcription des ARN 72
2.2.2 Marquage radioactif d'ARN en 5' 72
2.2.3 Électrophorèse comparative sur gel (Comparative gel elecîrophoresis) 73
2.2.4 Cartographie chimique {in line probing) 75
2.3 RÉSULTATS 76
2.3.1. Caractérisation de la rotation de la tige PI 76
2.3.2. La rotation de la tige PI est observable dans le cas d'un aptamère à 3 voies 83
2.4 DISCUSSION 88
CONCLUSION 95
ANNEXE 1 98
ANNEXE 2 101
LISTE DES ABRÉVIATIONS
A Adénine
ADN Acide désoxyribonucléique
ADNdb Acide désoxyribonucléique double brin AdoCbl Adénosylcobalamine ou vitamine B12 AMPc Adénosine monophosphate cyclique
AntiSD AntiShine-Dalgamo
ARN Acide ribonucléique
ARNm ARN messager
ARNnc ARN non codant
ARNpol ARN polymérase
ARNr ARN ribosomal
ARNt ARN de transfert
AT Antiterminateur
ATP Adénosine triphosphate
°C Degré celsius
C Cytosine
C3 Cyanine 3
CAP Protéine activatrice de gènes cataboliques (Catabolite gene Aetivater
Protein)
CGE Électrophorèse comparative sur gel (comparative gel electrophoresis) ci-di-GMP Guanosine monophosphate cyclique
Ci Curie
C0B12 Coenzyme B12
dG 2'-déoxyguanosine
DMS Sulfate de diméthyl
Dithiothréitol ou réactif de Cleland Efficience de FRET
Fluorescéine (essais de FRET) Femto (ÎO15)
Flavine mononucléotide
Transfert d'énergie par résonnance de fluorescence (Fluorescence
resonance energy transfer)
Facteur de transcription Guanine
Gramme
Glucosamine-6-Phosphate Dinucléotide guanine-cytosine
Acide chlorhydrique (hydrochloride acid) dû
Integrated DNA Technologies
Jonction
Constante d'association Constante de dissociation Chlorure de potassium
acide 4-(2-hydroxyéthyl)-l-pipérazine éthane sulfonique (pH ajusté grâce à du KOH) Hydroxyde de potassium Litre Molaire Milli (10 3) Méthionine adenosyltransférase
Concentration apparente de magnésium où la moitié du repliement induit par le magnésium est effectué dans la population de molécules Chlorure de magnésium (magnésium chloride)
MicroARN
n Coefficient de Hill
N.A. Non applicable
NaCl Chlorure de sodium (sodium chloride)
n Nano (109)
NMIA iV-méthylisatoic anhydre (N-methylisatoic anhydride)
nt Nucléotide
O État ouvert du riborégulateur SAM-I Opéron lac Opéron lactose
Opéron trp Opéron tryptophane
PI Tige 1 (paired 1)
P2 Tige 2 (paired 2)
P3 Tige 3 (paired 3)
P4 Tige 4 (paired 4)
P Pico (1012)
pARNn Petit ARN nucléaire
pARNno Petit ARN nucléolaire
pb Paire de bases
PCR Réaction en chaîne par polymérase (Polymerase Chain Reaction)
pH Potentiel hydrogène
PL Transcription pleine longueur
PTI® Photon Technology International (R)
Pré-Ql Pré-queosinel ou 7-aminomethyl-7-deazaguanine
R Purine (adénine ou guanine)
rATP Ribonucléotide adénosine triphosphate rCTP Ribonucléotide cytosine triphosphate rGTP Ribonucléoside guanosine triphosphate
RIG Région intergénique
RM8 Repliement dépendant du magnésium (préorganisation du site de liaison
du ligand)
Rn Repliement menant à l'état natif du riborégulateur SAM-I
RNase Ribonucléase
rNTP Ribonucléotide triphosphate
RT-qPCR PCR quantitative en temps réel (Quantitative RT-PCR) rUTP Ribonucléotide uracile triphosphate
S 5-adénosylméthionine (voir aussi SAM)
SAH S-adénosylhomocystéine
SAM S-adénosylméthionine (voir aussi S)
SAP Phosphatase alcaline de crevette (Shrimp alkaline phosphatase)
SD Séquence Shine-Dalgarno
SHAPE Acylation sélective du 2'-hydroxyle analysée par extension d'amorces
(Selective 2 '-Hydroxyl Acylation analysed by Primer Extension)
Term Terminateur ou tige terminatrice
T Transcription terminée
T4 PNK Polynucléotide kinase du phage T4 (T4 polynucleotide kinase)
TBE Tris - acide borique - EDTA
TE Tris-EDTA
TPP Thiamine pyrophosphate
Tris Trishydroxyméthylaminométhane
TS Type sauvage
Tuco Cofacteur du tungstène
U Uracile
UTR Région non transcrite (untranslated région)
V/V Volume/volume
32P Isotope radioactif de phosphore
5-NU Groupement amino-allyle en 5' d'un uracile
LISTE DES TABLEAUX
INTRODUCTION
Tableau 1.1. Riborégulateurs liant des acides aminés 16
Tableau 1.2. Riborégulateurs liant des bases azotées 17
Tableau 1.3. Riborégulateurs liant des coenzymes et cofacteurs 18
Tableau 1.4 Riborégulateurs liant d'autres types de ligand 20
Tableau 1.5. Distribution phylogénétique des sous-classes de riborégulateurs SAM 24
Tableau 1.6. Résumé des caractéristiques des sous-classes de riborégulateurs SAM 31
CHAPITRE 1 - ÉTUDE DE LA JONCTION 1/4 ET DE L'APPARIEMENT DU CŒUR Tableau 1.1. Identité des nucléotides formant l'appartement du cœur 41
Tableau 1.2. Séquence des ARN composantie vecteur fluorescent 1-3 46
Tableau 1.3. Séquence des ARN composant le vecteur fluorescent 1/4 47
Tableau 1.4. Efficience de FRET de différents mutants de la jonction 1/4 et de l'appariement
du cœur mesuré avec le vecteur fluorescent 1-3 57
Tableau 1.5. Efficience de FRET de différents mutants de la jonction 1/4 et de l'appariement
CHAPITRE 2 - ÉTUDE DE LA ROTATION DE LA TIGE PI
Tableau 2.1. Séquences des ARN qui composent la construction CGE 1 SAM-I à 4 voies contenant une boucle interne en 3' dans la tige PI
-3 du riborégulateur 73
Tableau 2.2. Séquences des ARN qui composent la construction CGE 1-3 du riborégulateur SAM-I à 4 voies contenant une boucle interne en 5' dans la tige PI 74
Tableau 2.3. Séquences des ARN qui composent la construction CGE 1-3 du riborégulateur SAM-I à 3 voies contenant une boucle interne en 3' dans la tige PI * 74
Tableau 2.4. Séquences des ARN qui composent la construction CGE du riborégulateur SAM-I à 3 voies contenant une boucle interne en 5' dans la tige P3 75
LISTE DES FIGURES
INTRODUCTION
Figure 1.1. Principe de base de l'expression génétique 2
Figure 1.2. Mécanismes de régulation génétique chez les bactéries 4
Figure 1.3. Représentation générale d'un ARNm contenant un riborégulateur 12
Figure 1.4. Mécanismes de régulation transcriptionnelle et traductionnelle 14
Figure 1.5. Structures chimiques de la S-adénosylméthionine (SAM) et de la
S-adénosyl-homocystéine 21
Figure 1.6. Sous-classes de riborégulateurs impliquées dans la régulation de la biosynthèse de la S-adénosylméthionine (SAM) et de la 5-adénosylhomocyatéine (SAH) 23
Figure 1.7. Séquence et structure consensus des riborégulateurs SAM 25
Figure 1.8. Les riborégulateurs SAM-I chez Bacillus subtilis et emplacement dans le génome
de la bactérie 32
Figure 1.9. Structures prédites du riborégulateur SAM-I en absence et en présence de SAM ..33
Figure 1.10. Structure du riborégulateur SAM-I yitJ 34
Figure 1.11. Modèle des mécanismes de repliement et de reconnaissance du ligand par Heppell
CHAPITRE 1 - ÉTUDE DE LA JONCTION 1/4 ET DE L'APPARIEMENT DU CŒUR Figure 1.1. Éléments structuraux importants pour le repliement et le fonctionnement du
riborégulateur SAM-1 40
Figure 1.2. Implication des nucléotides de la jonction 1/4 et de l'appariement du cœur pour
le fonctionnement du riborégulateur 50
Figure 1.3. Importance de l'appariement du cœur pour le fonctionnement du riborégulateur. 51
Figure 1.4. Structure globale de Faptamère de type sauvage et du mutant Al 38U 54
Figure 1.5. L'appariement du cœur n'est pas impliquée dans la préorganisation du site de
liaison du ligand (RMg) 56
Figure 1.6. L'appariement du cœur est important pour le repliement menant à l'état natif
(RN) du riborégulateur SAM-1 59
Figure 1.7. La présence de pyrimidines dans la jonction 1/4 nuit à l'activité et au repliement
du riborégulateur SAM-1 62
Figure 1.8. Interactions moléculaires de l'interaction triple entre les nucléotides A52, U114
CHAPITRE 2 - ÉTUDE DE LA ROTATION DE LA TIGE PI
Figure 2.1. Rotation de la tige PI suite à la liaison de la SAM à Faptamère du riborégulateur
SAM-1 77
Figure 2.2. Exemples de constructions CGE 1-3 utilisées pour caractériser la phase ondulatoire de la rotation de la tige PI du riborégulateur SAM-1 78
Figure 2.3. Caractérisation de la phase ondulatoire de la rotation de la tige PI 82
Figure 2.4. La rotation de la tige PI est indépendante de la présence de la tige P4 87
Figure 2.5. Changement de conformation du riborégulateur SAM-1 90
Figure 2.6. Variabilité dans la tige P3 du riborégulateur SAM-1 92
ANNEXES
Figure A.l. Vecteurs fluorescents 1-3 (gauche) et 1-4 (droite) utilisés pour les essais de
FRET 98
Figure A.2. Effet de différents mutants ponctuels de la jonction 1/4 (CAA et GUA) et mutants de l'appariement du cœur (GAA* et GAU) sur la préorganisation du site de liaison du
ligand du riborégulateur SAM-I 99
Figure A.3. Effet de différents mutants ponctuels de la jonction 1/4 (CAA, UAA, GCA, GUA, CCA, ACA, AAC, GUU, CCC et UUU) et mutants de l'appariement du cœur (GAA* et GAU) sur le réarrangement menant à la conformation native du riborégulateur SAM-1 100
INTRODUCTION
A. Les avancées de la biologie moléculaire
De nombreuses découvertes scientifiques survenues au cours du XXe siècle ont, sans contredit,
mené à une révolution au niveau de la compréhension des mécanismes cellulaires et moléculaires des organismes vivants. C'est en 1938 que Warren Weaver utilisa pour la première fois le concept de biologie moléculaire afin de décrire les techniques modernes utilisées pour étudier les détails de certains procédés de la vie (Rees, 1987). Quinze années plus tard, soit en 1953, le biologiste James Watson et le physicien Francis Crick ont découvert la molécule à la base de la vie soit l'acide désoxyribonucléique (ADN) (Watson et Crick, 1953). Au fil des années, l'avancement des technologies a permis de déchiffrer le code génétique qui contient l'ensemble des informations nécessaires au développement et au fonctionnement de tous les organismes vivants. En effet, il y a maintenant dix ans, le premier génome humain complet a été séquencé, soit celui du biologiste américain John Craig Venter (Venter et al., 2001). De nos jours, la biologie moléculaire est considérée comme étant une discipline à la croisée entre la génétique, la biochimie et la physique qui vise à élucider les mécanismes reliés aux fonctions métaboliques d'un organisme. En effet, il est maintenant possible, comme il a été mentionné précédemment de séquencer des génomes, mais aussi de séquencer des transcriptomes complets (Carninci, 2009; Tariq et al., 2011), d'analyser et comparer des protéomes (Langen et al., 1997), de connaître la structure tridimensionnelle de plusieurs molécules (Wenk et al., 1992) ou encore, de faire des études à l'échelle de la molécule unique (single molecule) (Anazawa et al., 2002).
B. L'expression génétique et le métabolisme
L'expression génétique, principe sur lequel repose la biologie moléculaire, est définie comme étant le procédé par lequel l'information contenue dans un gène est utilisée afin de produire un élément fonctionnel, c'est-à-dire une protéine ou un acide ribonucléique non codant (ARNnc). Dans le cas où l'élément fonctionnel à obtenir est une protéine, l'expression génétique est décrite selon deux étapes principales soit la transcription et la traduction. En effet, l'ADN est transcrit par TARN polymérase (ARN pol) en ARN messager (ARNm). L'ARNm est alors reconnu par les ribosomes qui le traduisent en protéine (Figure 1.1). De manière différente, les ARNnc, tels que les ARN ribosomaux (ARNr), les ARN de transfert (ARNt), les petits ARN nucléaires (pARNn), les petits ARN nucléolaires (pARNno), les microARN (miARN) ou autres sont des ARN fonctionnels qui ne sont pas reconnus par les ribosomes, ainsi ils ne sont pas traduits dans le but de produire une ou plusieurs protéines.
Transcription Traduction
gsg
> jyip
Figure 1.1. Principe de base de l'expression génétique. Les flèches pleines représentent les étapes d'expression génétiques menant de l'ADN à la formation d'une protéine.
Bien que les protéines assurent divers rôles dans la cellule comme des fonctions structurales, des fonctions motrices ou des fonctions de transport intra et extracellulaire, bon nombre d'entre elles sont essentielles aux fonctions associées à l'expression génétique ou au métabolisme. En effet, certaines protéines agissent au niveau de la signalisation cellulaire ou au niveau du contrôle de l'expression de certains gènes. De plus, certaines protéines assurent des fonctions enzymatiques, c'est-à-dire qu'elles catalysent directement des réactions associées au métabolisme cellulaire. Le métabolisme cellulaire peut être défini comme étant
l'ensemble des réactions chimiques permettant le catabolisme (dégradation) ou l'anabolisme (synthèse) de molécules organiques. Ainsi, le métabolisme permet la formation de métabolites et d'énergie essentiels à la survie de la cellule.
B.l. Régulation de l'expression génétique et du métabolisme chez les bactéries
La survie d'une bactérie dépend de son habileté à croître, et ce, même lorsque les conditions ne sont pas idéales. Son énergie est utilisée afin de synthétiser différents métabolites nécessaires à sa croissance et à son fonctionnement. Afin d'économiser au maximum cette énergie, une bactérie doit être en mesure de réguler finement l'expression de ses gènes et de son métabolisme. De cette manière, plusieurs mécanismes de régulation permettent une production adéquate de différentes molécules fonctionnelles (Figure 1.2). Ainsi, divers acteurs comme les facteurs sigma (facteurs a), les facteurs de transcription (FT), les systèmes d'antiterminaison (AT), les facteurs d'élongation de la transcription, les ARN régulateurs dépendants d'un signal, les petits ARN régulateurs, les protéines qui lient des ARN et des ribonucléases sont responsables de contrôler les différentes étapes de l'expression génétique ainsi que du métabolisme cellulaire (Roth et Breaker, 2009). Grâce à tout cela, la bactérie exerce un contrôle précis sur la transcription et la traduction de ses gènes et par conséquent, sur les réactions enzymatiques propres à son métabolisme.
ARNpo).rNTP ?rx4.**,<r 'ie 'A-'/rptior rma £WSK | Mécanismes de régulation | Facteurs sigma
Autres facteurs de transcription
Systèmes d'anttterminaiscn Fadeurs tféiongation de la transcription
Signal-réponse, ARN régulateurs en en
ARNm " Rfoosomes
RNase
Petits ARN régulateurs Protéines liant les ARNm
Signai-réponse, ARN régulateurs en cis
x (Réactions enzymatiques)
Petits ARN régulateurs Protéines liant les ARNm
Signai-réponse, ARN régulateurs en cis S X
a
Figure 1.2. Mécanismes de régulation génétique chez les bactéries. (A) Les facteurs sigma et autres facteurs de transcription (FT) contrôlent l'initiation de la transcription. Les étapes qui suivent l'initiation de la transcription sont régulées par différents mécanismes tels que (B) des systèmes d'antiterminaison et d'autres protéines influençant l'élongation, (C) des ARN régulateurs répondant à un signal et agissant en cis pour contrôler la terminaison de la transcription et (D-E) des petits ARN régulateurs, des protéines liant des ARNm et des ARN régulateurs en cis contrôlent l'expression génétique au niveau de la traduction et de la dégradation des ARNm. Adaptée de Roth et Breaker (2009).
B.l.l. De l'ADN à I'ARNm : régulation de la transcription
Tout d'abord, puisque la transcription est la première étape de l'expression génétique, un contrôle à ce niveau diminue grandement le gaspillage d'énergie. En effet, l'inhibition de la transcription permet d'éviter de synthétiser des molécules qui ne sont pas requises par la cellule. De cette manière, l'initiation de la transcription est une cible majeure pour différents mécanismes de régulation de l'expression génétique chez les bactéries (Winkler et Breaker, 2005). L'association entre différents facteurs sigma et l'ARN polymérase permet de reconnaître spécifiquement le promoteur approprié et ainsi d'initier la transcription de gènes nécessaires à un stade particulier de croissance de la bactérie ou encore en conditions de stress (Roth et Breaker, 2009). En effet, la présence de certains facteurs de stress ou de nouvelles molécules associées à un changement de conditions physiologiques peut mener à une compétition entre les différents facteurs sigma pour la liaison à l'ARN polymérase. Cette compétition aboutit ainsi à l'activation ou la répression de l'expression de divers gènes (Mooney et al., 2005; Navarro Llorens et al., 2010). À l'instar des facteurs sigma, l'initiation de la transcription peut aussi être régulée en réponse à l'association entre différents facteurs de transcription et l'ADN ou l'ARN polymérase (Browning et Busby, 2004). Le séquençage de génomes bactériens a permis d'identifier des centaines de facteurs de transcription putatifs, et ce, pour une seule bactérie. Par exemple, il est prédit que plus de 200 facteurs de transcription seraient codés par le génome de Bacillus subtilis (Winkler et Breaker, 2005).
De cette manière, une bactérie peut réguler son métabolisme cellulaire à la suite de réactions enzymatiques. Ainsi, le niveau de production d'enzymes inductibles, c'est-à-dire non constitutives, peut être contrôlé à la hausse ou à la baisse en réponse à l'absence ou la présence d'un certain substrat. L'exemple le plus souvent cité dans la littérature concernant ce type de régulation est, sans contredit, l'opéron lactose (lac) qui résulte en l'activation ou la répression de la synthèse d'enzyme (Jacob et Monod, 1961). En effet, l'opéron lactose comprend deux sites régulateurs nommés le promoteur et l'opérateur ainsi que les gènes lacZ, lacY et lacA
codant pour des protéines impliquées dans le transport et le métabolisme du lactose. En absence de lactose, une protéine répressive se lie sur l'opérateur, ce qui empêche l'ARN polymérase de transcrire les gènes de l'opéron lac. Par contre, lorsque du lactose est présent dans la cellule, il se lie à la protéine répressive et l'inactive, permettant ainsi de produire les enzymes codées par les gènes lac. De cette manière, le lactose présent dans la cellule peut être métabolisé. De plus, cet opéron comprend un deuxième mécanisme de régulation, celui-ci en réponse au glucose. En effet, en présence de lactose et de glucose dans la cellule, le répresseur est inactivé, mais il n'y a pas d'expression des gènes lacZ, lacY et lacA. Le glucose est utilisé par la cellule, ce qui résulte en l'accumulation d'adénosine monophosphate cyclique (AMPc) qui forme un complexe avec la protéine activatrice des gènes cataboliques (CAP). Le complexe CAP-AMPc se lie au promoteur de gènes de l'opéron lactose se qui permet d'augmenter l'affinité de l'ARN polymérase au promoteur et ainsi augmenter la transcription des gènes.
Par la suite, lors de l'élongation de la transcription, l'incorporation de chaque ribonucléotide à l'ARNm naissant peut avoir différents effets sur la transcription. En effet, l'ARN polymérase peut simplement continuer l'élongation ou encore, peut faire une pause, arrêter, revenir en arrière ou terminer la transcription (Greive et von Hippel, 2005; Vassylyev, 2009). De cette manière, plusieurs facteurs d'élongation, séquences spécifiques sur l'ARN et systèmes d'antiterminaison permettent de favoriser l'une ou l'autre des << décisions » que peut prendre l'ARN polymérase lors de l'élongation de la transcription (Roth et Breaker, 2009). Finalement, la dernière étape de la transcription, soit la terminaison, peut être régulée via des ARN régulateurs qui agissent en cis, c'est-à-dire qui régulent leur propre expression. D s'agit habituellement d'une portion d'ARNm sensible à un certain signal qui engendre sa propre régulation, soit la continuation de la transcription ou encore un arrêt prématuré de celle-ci. Par exemple, les riborégulateurs agissent de cette manière en empêchant ou favorisant la formation d'une tige terminatrice indépendante de la protéine Rho (Roth et Breaker, 2009). La régulation transcriptionnelle induite par les riborégulateurs sera discutée plus en détail à la section C de l'introduction.
B.1.2. De l'ARNm à la protéine : régulation de la traduction
Plusieurs mécanismes permettent un contrôle fin de la synthèse de la protéine ou des protéines codées par un ARNm. Ce type de régulation peut avoir lieu en même temps que la transcription, c'est-à-dire de manière co-transcriptionnelle, ou encore une fois la transcription de l'ARNm achevée. Par exemple, certains ARN permettent la régulation en cis de la traduction en réponse à un signal. Effectivement, il existe des riborégulateurs qui contrôlent l'initiation de la traduction en séquestrant la séquence Shine-Dalgarno (SD) et/ou le codon d'initiation de la traduction (Roth et Breaker, 2009). De cette manière, le riborégulateur empêche la liaison du ribosome à l'ARNm, ainsi la traduction ne peut pas avoir lieu et aucune protéine ne sera produite à partir de cet ARNm. La régulation traductionnelle via les riborégulateurs sera discutée plus en détail à la section C de cette introduction.
En plus de ces ARN régulateurs agissants en cis, certains petits ARN peuvent agir en trans, c'est-à-dire au niveau d'un autre ARNm. Par exemple, un petit ARN peut se lier à la région non codante en 5' d'un ARN messager, empêchant ou favorisant son accès par le complexe d'initiation de la traduction (Gottesman et Storz, 2010; Waters et Storz, 2009). De plus, il a été récemment démontré que des petits ARN pouvaient s'apparier à la région codante d'un ARNm. Cependant, il reste à déterminer si ces interactions sont impliquées dans la régulation de la traduction ou sur la stabilité de l'ARNm (Pfeiffer et al., 1009). De plus, certains petits ARN agissent en trans via la liaison à une protéine liant l'ARN. L'exemple le plus connu est retrouvé chez Escherichia coli où les petits ARN CsrB et CsrC se complexent avec la protéine CsrA. La protéine CsrA est une protéine liant l'ARN de façon post-transcriptionnelle, ce qui influence l'accessibilité des ribosomes et la stabilité de cet ARNm (Baker et al., 2002; Liu et al., 1997; Liu et Romeo, 1997). De plus, certaines protéines lient directement les ARNm, sans l'intermédiaire de petits ARN régulateurs, et influencent la stabilité de ces derniers empêchant, par exemple, la dégradation des ARNm via des ribonucléases (Roth et Breaker, 2009).
Un autre exemple de régulation au niveau de la traduction est l'atténuation traductionnelle comme celle effectuée par l'opéron tryptophane (Morse et al., 1969). À l'inverse de l'opéron lactose, l'expression des gènes de l'opéron tryptophane est atténuée en présence de tryptophane. En effet, lorsque le tryptophane est présent dans la cellule, cet acide aminé se lie sur une protéine répressive et l'active. Ainsi, la protéine répressive liée au tryptophane diminue la transcription des gènes de l'opéron qui codent pour des enzymes nécessaires à la synthèse de cet acide aminé. À l'inverse, lorsque le tryptophane n'est pas disponible dans la cellule, la protéine répressive non liée à l'acide aminé reste inactive. Puisque la protéine ne peut pas assurer sa fonction de répression, les enzymes de synthèse du tryptophane sont donc synthétisées.
B.1.3. De la protéine au métabolite cellulaire : régulation métabolique
Le dernier mécanisme de régulation du métabolisme est la rétro-inhibition d'enzyme. D s'agit d'un processus par lequel le produit final de la voie métabolique inhibe, de manière allostérique, l'activité de la première enzyme unique à cette voie de biosynthèse. De cette manière, la liaison du métabolite final au site allostérique de l'enzyme altère le site de liaison du substrat sur l'enzyme qui ne peut plus exercer sa fonction. Par contre, les autres enzymes de la voie métabolique restent actives même si leur substrat n'est pas produit. La répression du métabolisme esLmaintenue jusqu'à ce que la concentration Hn métahnlite final soit insuffisante pour agir sur l'enzyme.
C. Les riborégulateurs
C.l. Découverte des riborégulateurs
Pendant plusieurs années de nombreux groupes de recherche se sont intéressés à des gènes connus pour contrôler certaines voies métaboliques bactériennes, sans toutefois connaître les protéines impliquées dans leur régulation (Christiansen et al., 1997; Gelfand et al., 1999; Grundy et Henkin, 1998; Miranda-Rios et al., 2001; Nou et Kadner, 2000; Patte et al., 1998). Par exemple, le groupe de recherche de Mikhail S. Gelfand a démontré que certaines mutations introduites à l'intérieur d'une structure complexe adoptée dans la région 5' non traduite (5' UTR) de certains ARNm codant pour des gènes associés à l'opéron riboflavine chez les bactéries Bacillus subtilis et Bacillus amyloliquefaciens affectaient la synthèse de la riboflavine (Gelfand et al., 1999). C'est alors que ce groupe de recherche a postulé qu'il était possible que cette structure d'ARNm interagisse avec un élément de régulation. De cette façon, l'interaction entre l'élément régulateur inconnu et l'ARNm pourrait affecter la conformation de ce dernier, et ainsi, agir sur le niveau d'expression du gène (Gelfand et al., 1999). À cette époque, il était pratiquement inconcevable de croire à une régulation génétique où il n'y avait aucune implication protéique. D'ailleurs, ce n'est que trois années plus tard que le groupe de recherche de Ronald R. Breaker a démontré que la liaison d'un petit métabolite à la région 5' non traduite d'un ARNm naissant permettait de contrôler l'expression génétique. (Nahvi èt al., 2002; Winkler et al., 2002a). Le terme riborégulateur (riboswitch) a été utilisé pour la première fois par l'équipe de Ronald R. Breaker en 2002 pour décrire ce type d'ARNm régulateur agissant en cis (Winkler et al., 2002a). Aujourd'hui, plus d'une vingtaine de classes de riborégulateurs ont été identifiées chez des organismes faisant partie des trois domaines du vivant, soit les procaryotes, les eucaryotes et les archées (Poiata et al., 2009; Roth et Breaker, 2009). D est estimé que près de 4 % des gènes bactériens seraient régulés par un mécanisme impliquant la liaison d'un métabolite à un ARNm (Kelley et Hamelberg, 2010).
C.2. Définition du concept de riborégulateur
Tout d'abord, il est important de mentionner que tous les biologistes ne s'entendent pas sur la définition de ce qu'est un riborégulateur. Cependant, certaines caractéristiques sont majoritairement acceptées lorsqu'il s'agit de définir le concept. De cette manière, un riborégulateur est défini comme étant une structure d'ARNm, généralement retrouvée dans la région 5' non traduite, qui contrôle le niveau d'expression génétique (Bastet et al., 2011). La liaison spécifique d'un métabolite (ou ligand) à cette structure d'ARN entraîne un changement de conformation, ce qui entraîne une régulation positive ou négative de l'expression du gène ou des gènes en aval du riborégulateur (Barrick et Breaker, 2007). De manière générale, la fonction de la protéine ou des protéines codées par le gène sous le contrôle du riborégulateur est directement reliée au métabolisme du ligand ayant entraîné cette régulation. La confusion dans la définition du concept de riborégulateurs vient souvent lorsqu'il s'agit de déterminer si certains ARN régulateurs répondant au changement de pH (Nechooshtan et al., 2009) ou de température (Chowdhury et al., 2003) sont des riborégulateurs ou non. En effet, comme le pH et la température ne sont pas des métabolites, certains biologistes considèrent qu'ils ne constituent pas des classes de riborégulateurs, mais plutôt ils feraient partie d'une catégorie complètement différente de régulateurs. Cependant, il est à noter que ces ARN répondant au pH ou à la température sont considérés, dans ce document, comme des classes de riborégulateurs.
Plusieurs groupes de recherche voient en ces petits ARN régulateurs un fort potentiel en tant que cible d'une nouvelle classe d'antibiotiques puisqu'ils sont retrouvés chez une multitude de bactéries pathogènes tout en n'étant pas retrouvés chez les humains (Blount et Breaker, 2006; Blount et al., 2007; Lee et al., 2009; Mulhbacher et al., 2010; Nudier et Mironov, 2004; Ott et al., 2009; Sudarsan et al., 2005). Le principe qui sous-tend cette hypothèse de fort potentiel de cible pour un antibiotique est le suivant. Puisque la majorité des riborégulateurs inhibent l'expression génétique lorsqu'ils sont en association avec leur ligand, il suffit de trouver un
métabolite analogue à celui-ci qui pourrait aussi se lier au riborégulateur. Ainsi, la liaison de cet analogue exogène au riborégulateur permettrait d'inhiber une ou plusieurs voies métaboliques essentielles à la survie de la bactérie pathogène ce qui permettrait ainsi de combattre l'infection efficacement. De plus, ils possèdent une spécificité très élevée pour certains métabolites, ce qui permet de discriminer entre plusieurs molécules semblables et ainsi rendre les riborégulateurs très efficaces (Blount et Breaker, 2006; Deigan et Ferre-D'Amare, 2011). À cet effet, notre groupe de recherche a publié récemment un article concernant l'utilisation d'un analogue de la guanine afin de réduire une infection causée par la bactérie Staphylococcus aureus dans le cas de la mammite murine (Mulhbacher et al., 2010).
La structure d'un riborégulateur est composée de deux domaines. En premier lieu, la partie très structurée en 5' du riborégulateur possédant la capacité de lier le ligand est nommée
aptamère (Figure 1.3). La séquence et la structure de l'aptamère sont généralement hautement
conservées dans la même classe de riborégulateur, c'est-à-dire chez les différents riborégulateurs liant le même ligand. Par contre, deux ligands particuliers, soit la pré-queuosine, (Pré-Qi) et la 5-adénosylméthionine (SAM), sont connus pour lier plus d'une structure d'ARN. Dans ces cas particuliers, les différents motifs de riborégulateur pouvant lier le même métabolite sont considérés comme étant des sous-classes de ce riborégulateur (Poiata et al., 2009; Roth et al., 2007). En second lieu, la régulation génétique est possible grâce à une structure beaucoup moins conservée se retrouvant en 3' de l'aptamère du riborégulateur. Cette structure est retrouvée majoritairement sous la forme de tige-boucle et est nommée plateforme
d'expression (Figure 1.3). D est à noter qu'une partie en 3' de l'aptamère ainsi qu'une partie en
i
Aptamère ---<
. Plateforme .
' d'expression '
Figure 1.3. Représentation générale d'un ARNm contenant un riborégulateur. Le riborégulateur, se retrouvant dans la région 5' non traduite (5' UTR) de l'ARNm, comprend deux domaines. L'aptamère est la structure ayant la capacité de lier de manière spécifique un ligand et la plateforme d'expression est le domaine qui permet la régulation génétique. Adaptée et modifiée de Roth et Breaker (2009).
C.3. Mécanismes de régulation utilisés par les riborégulateurs
Les riborégulateurs contrôlent l'expression génétique en utilisant cinq mécanismes. Les deux mécanismes les plus répandus et les plus étudiés sont ceux qui entraînent une régulation au niveau transcriptionnel ou traductionnel en cis. Cependant, il existe aussi des riborégulateurs qui contrôlent l'épissage alternatif d'introns du groupe I (Cheah et al., 2007; Hartig, 2010; Poiata et al., 2009; Wachter et al., 2007), qui régulent en antisens (André et al., 2008; Loh et al., 2009) ou qui agissent sur la stabilité de l'ARNm (Winkler et al., 2004). De façon générale, les gènes régulés par les riborégulateurs codent pour des protéines de synthèse appartenant à la voie métabolique menant à la formation du métabolite liant ce riborégulateur. En absence de ligand, la conformation du riborégulateur, nommé apo-riborégulateur, est dite active (ON) et permet l'expression du gène ou des gènes en aval du riborégulateur. À l'inverse, en présence de ligand et à la suite de sa liaison au riborégulateur, celui-ci adopte une conformation dite
inactive (OFF) qui empêche l'expression du gène ou des gènes en aval du riborégulateur. Il
existe aussi des riborégulateurs, comme celui liant l'adénine où la présence de ligand favorise l'adoption de la conformation active (Lemay et al., 2006).
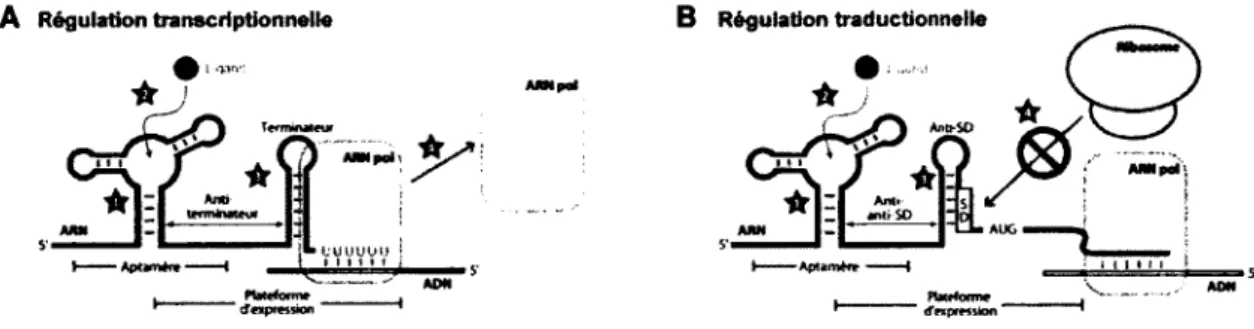
C.3.1. Mécanisme de régulation transcriptionnelle
Le réarrangement structural du riborégulateur suivant la liaison du ligand entraîne la formation d'une tige terminatrice intrinsèque (Figure 1.4). Cette structure en tige-boucle suivie d'une série d'uraciles est connue pour déstabiliser le complexe formé entre l'ADN, l'ARN polymérase (ARN pol) et l'ARNm naissant (Landick et al., 1996; Merino et Yanofsky, 2002; Vitreschak et al., 2004). De cette façon, la polymérase et l'ARNm naissant décrochent de la matrice d'ADN, ce qui empêche la transcription d'être poursuivie. Ainsi, l'expression génétique est réprimée puisque l'ARNm n'est pas complété (Mandai et Breaker, 2004; Poiata et al., 2009; Tomsic et al., 2008). De plus, il est à noter que cet arrêt prématuré de la transcription s'effectue avant même que le codon d'initiation de la traduction soit transcrit.
C.3.2. Mécanisme de régulation traductionnelle
À l'instar de la régulation transcriptionnelle, la liaison du ligand à l'aptamère entraîne la formation d'une tige-boucle (Figure 1.4). Cependant, il ne s'agit pas d'une tige terminatrice. Cette structure entraîne plutôt une séquestration de la séquence Shine-Dalgarno (SD) et/ou du codon d'initiation de la traduction empêchant ainsi le recrutement du ribosome sur l'ARNm naissant. De cette façon, l'expression génétique est réprimée au niveau de l'initiation de la traduction (Fuchs et al., 2006; Pôîata et al., 2009). Par contre, contrairement à la régulation transcriptionnelle, l'ARNm est transcrit au complet.
A Régulation transe ri ptionnelle B Régulation traductionnalte
S' S'i
Figure 1.4. Mécanismes de régulation transcriptionnelle et traductionnelle.
(A) Riborégulateur réprimant l'expression d'un gène via un mécanisme de régulation transcriptionnelle. (1) L'aptamère adopte une structure permettant (2) la liaison du ligand, (3) ce qui mène à la formation d'une tige terminatrice (4) permettant le décrochage de l'ARN polymérase. (B) Riborégulateur réprimant l'expression d'un gène via un mécanisme de régulation traductionnelle. Les numéros (1) et (2) ont été expliqués précédemment. (3) La tige formée séquestre la séquence Shine-Dalgarno (antiSD) ce qui (4) empêche la liaison du ribosome à l'ARNm. Adaptée et modifiée de Roth et Breaker (2009).
C.3.3. Épissage alternatif
Le riborégulateur liant la thiamine pyrophosphate (TPP) est le seul riborégulateur retrouvé chez les eucaryotes où il agit sur l'épissage alternatif (Cheah et al., 2007; Poiata et al., 2009; Wachter et al., 2007). L'aptamère du riborégulateur est situé dans un intron qui contient deux sites potentiels d'épissage en 5'. En absence de ligand, la partie conservée de l'aptamère permettant la liaison au TPP fait une interaction de type paire de bases avec une séquence près du deuxième site d'épissage, ce qui rend le premier site plus accessible pour le complexe d'épissage (spliceosome). À l'inverse, lorsque le TPP est lié au riborégulateur, le deuxième site d'épissage est reconnu préférentiellement par le complexe d'épissage. En plus du riborégulateur TPP, un riborégulateur agissant sur l'épissage alternatif chez Clostridium
liant la guanosine monophosphate cyclique (c-di-GMP) (Hartig, 2010). Par contre, le mécanisme de ce riborégulateur est particulier puisque sa plateforme d'expression constitue un site d'autoépissage, c'est-à-dire qu'il s'agit d'un ribozyme agissant sur l'épissage alternatif d'introns du groupe I (Hartig, 2010). En effet, en absence de ligand, l'ARNm pleine longueur ne peut pas être traduit puisque le site d'initiation de la traduction est séquestré par une partie de la séquence d'introns. Cependant, lorsque la c-di-GMP se lie à l'aptamère du riborégulateur, l'autoclivage de l'ARNm libère le site d'initiation de la traduction, ce qui permet à l'ARNm d'être traduit en protéine (Hartig, 2010).
C.3.4. Régulation antisens
Jusqu'à présent, deux exemples de riborégulateurs ont été démontrés pour agir en trans, c'est-à-dire, au niveau de l'expression d'un autre gène. D s'agit des riborégulateurs SAM et ARNt (André et al., 2008; Loh et al., 2009). Ce mécanisme de régulation génétique est possible grâce à l'hybridation entre deux ARNm. Par exemple, l'appariement entre une partie de l'aptamère du riborégulateur SAM et sa séquence complémentaire retrouvée au niveau de l'ARNm du gène d'intérêt inhibe l'expression de ce dernier. Évidemment, l'interaction entre le riborégulateur et le gène d'intérêt dépend de la liaison du ligand (Loh et al., 2009).
C.3.5. Ribozyme
Deux riborégulateurs contrôlent l'expression génétique grâce à un autoclivage, c'est-à-dire qu'il s'agit de ribozymes. En effet, il s'agit du riborégulateur liant la c-di-GMP dont il a été question à la section C.3.3 et le riborégulateur liant la glucosamine-6-phosphate (GlcN6P). La liaison du ligand au riborégulateur mène à un autoclivage rapide de l'ARNm qui est ensuite dégradé par différents mécanismes de dégradation de la cellule (Winkler et al., 2004).
C.4. Classes de riborégulateurs
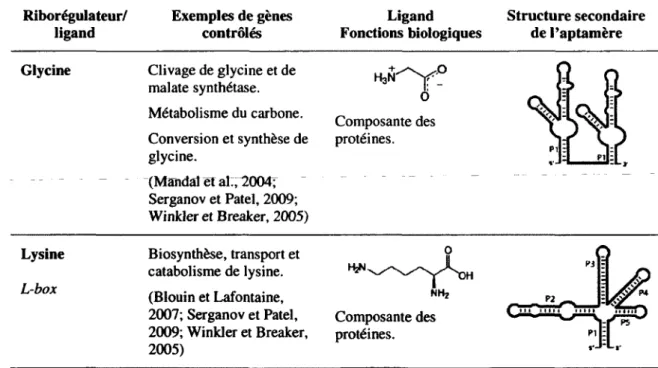
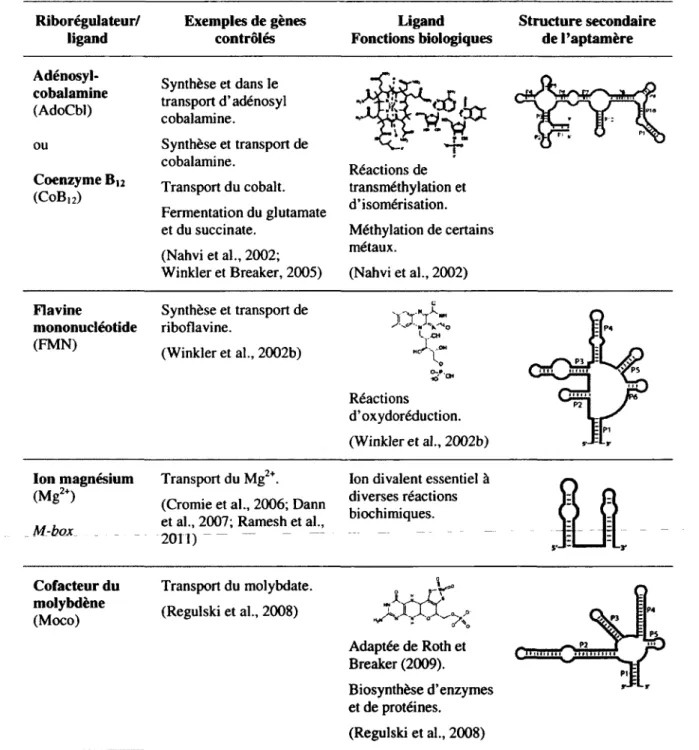
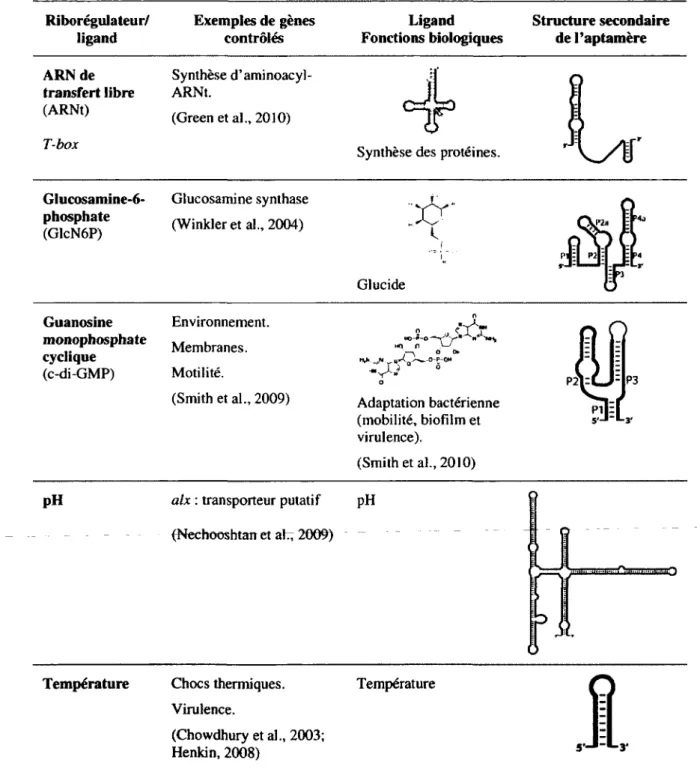
Comme il a été mentionné précédemment, plus d'une vingtaine de classes de riborégulateurs ont été découvertes jusqu'à ce jour. Ces éléments régulateurs sont connus pour lier de multiples métabolites tels que des acides aminés (glycine et lysine; tableau 1.1), des bases azotées (adénine, 2'-déoxyguanosine, guanine et pré-queuosinej ; tableau 1.2), des coenzymes et cofacteurs (adénosylcobalamine, flavine mononucléotide, ion magnésium, cofacteur du molybdène, S-adénosylhomocystéine, 5-adénosylméthionine, thiamine pyrophosphate, cofacteur du tungstène; tableau 1.3) ou d'autres types de ligand (ARNt, glucosamine-6-phosphate, guanosine monophosphate cyclique, pH et température; tableau 1.4).
Tableau 1.1. Riborégulateurs liant des acides aminés
Riborégulateur/ ligand Exemples de gènes contrôlés Ligand Fonctions biologiques Structure secondaire de l'aptamère
Glycine Clivage de glycine et de
malate synthétase. Métabolisme du carbone. Conversion et synthèse de glycine. (Mandai et al:, 12004^ Serganov et Patel, 2009; Winkler et Breaker, 2005) H3N O Composante des protéines. Lysine L-box Biosynthèse, transport et catabolisme de lysine. (Blouin et Lafontaine, 2007; Serganov et Patel, 2009; Winkler et Breaker, 2005) H2N NH2 Composante des protéines.
Tableau 1.2. Riborégulateurs liant des bases azotées Riborégulateur / ligand Exemples de gènes contrôlés Ligand Fonctions biologiques Structure secondaire de l'aptamère Adénine A-box Métabolisme de l'adénine. Transport de protéines et de purines. (Johansen et al., 2003; Lemay et al., 2006; Nygaard et Saxild, 2005) H Purine : composante de l'ADN et de TARN. 2'-déoxy-guanosine (dG) Ribonucléotide réductase. Transporteur de phosphate. (Edwards et Batey, 2009; Kim et Breaker, 2008) V-NH Ho-~t!rVY0 HCJ' H N~f NHj Analogue de la guanine. Guanine G-box Métabolisme et le transport des purines et de la guanine plus particulièrement. (Mulhbacher et al., 2010) Purine : composante de l'ADN et de l'ARN. Pré-queuosine] (PréQ.) Biosynthèse de la queosine. (Roth et al., 2007) H«N—„ O
V-
N-^N^nh, Intermédiaire dans la biosynthèse de la queuosine. (Iwata-Reuyl, 2003; Roth et al., 2007) PréQi-I.aîg.
PréQ,-Ujfi
Tableau IJ. Riborégulateurs liant des coenzymes et cofacteurs Riborégulateur/ ligand Exemples de gènes contrôlés Ligand Fonctions biologiques Structure secondaire de l'aptamère Adénosyl-cobalamine (AdoCbl) Synthèse et dans le transport d'adénosyl cobalamine. Cg T tj F|PI« 1 ou Synthèse et transport de •r! ' 2T *T* r ~ Coenzyme Bt2 (COB12) cobalamine. Réactions de Coenzyme Bt2
(COB12) Transport du cobalt. transméthylation et
Coenzyme Bt2 (COB12) Fermentation du glutamate et du succinate. (Nahvi et al., 2002; Winkler et Breaker, 2005) d'isomérisation. Méthylation de certains métaux. (Nahvi et al., 2002) Flavine mononucléotide (FMN) Synthèse et transport de riboflavine. (Winkler et al., 2002b) YY"7X"" S'04 A: Réactions d'oxydoréduction. (Winkler et al., 2002b) Ion magnésium (Mg2+) M-box _ . _ Transport du Mg2+.
(Cromie et al., 2006; Dann et al., 2007; Ramesh et al.,
2011) ~
Ion divalent essentiel à diverses réactions biochimiques. Cofacteur du molybdène (Moco) Transport du molybdate. (Regulski et al., 2008) Adaptée de Roth et Breaker (2009). Biosynthèse d'enzymes et de protéines. (Regulski et al., 2008) — M MH
Tableau 1.3. Riborégulateurs liant des cœnzymes et cofacteurs (suite)
Riborégulateur/ ligand
Exemples de gènes contrôlés
Ligand Structure secondaire
Fonctions biologiques de l'aptamère
S-adénosyl-homocystéine (SAH) Conversion du SAH en homocystéine. (Montange et al., 2009;
Wang et Breaker, 2008) Intermédiaire de synthèse de cystéine. (Wang et Breaker, 2008) 5-adénosyl-méthionine (SAM) S-box Métabolisme du soufre. Biosynthèse de la cystéine. Biosynthèse et recyclage de la méthionine. Biosynthèse de la SAM. Réductase de méthylène tétrahydrafolate. (Grundy et Henkin, 2004; Heppell et Lafontaine, 2008; Nudler et Mironov, 2004; Winkler et Breaker, 2005)
la
Dérivé de la méthionine. Réactions de méthylation (ADN, ARN, protéines). (Roje, 2006) SAM-I SAM-II SAM-IIl SAM-IV SAM-V Thiamine pyrophosphate (TPP) Thi-box Biosynthèse, phosphorylation et transport de thiamine. Épissage d'ARN chez les eucaryotes.(Cheah et al., 2007; Winkler et al., 2002a)
J.»
Participe aux réactions de décarboxylation. (Cheah et al., 2007) Cofacteur du tungstène (Tuco) Transport du tungstène. (Regulski et al., 2008) Réactions d'oxydoréduction (Bevers et al., 2008)
Tableau 1.4 Riborégulateurs liant d'autres types de ligand Riborégulateur/ ligand Exemples de gènes contrôlés Ligand Fonctions biologiques Structure secondaire de l'aptamère ARNde transfert libre (ARNt) T-box Synthèse d'aminoacyl-ARNt. (Green et al., 2010)
Synthèse des protéines.
Glucosamine-6-phosphate (GlcN6P) Glucosamine synthase (Winkler et al., 2004)
'XX'
Glucide Guanosine monophosphate cyclique (c-di-GMP) Environnement. Membranes. Motilité. (Smith et al., 2009) o " Y ]T un n j—\ o o»-Hi** /* u«<0./-«fc.-0-P-0M Adaptation bactérienne (mobilité, biofilm et virulence). (Smith et al., 2010)pH alx : transporteur putatif
(Nechooshtan et ah, 2009) pH ysssû A o maO
Température Chocs thermiques.
Virulence.
(Chowdhury et al., 2003; Henkin, 2008)
D. Riborégulateur SAM
D.l. 5-adénosylméthionine et S-adénosylhomocystéine
C'est en 1953 que le chercheur italien Giulio Catoni a découvert la S-adénosylméthionine (SAM) (Catoni, 1953; Loenen, 2006). À cette époque, cette molécule était principalement destinée au traitement de la dépression. La SAM est une molécule essentielle chez tous les organismes vivants grâce à son rôle de donneur de groupements méthyles. En effet, il s'agit d'une coenzyme clé lors des réactions de méthylation d'ADN, d'ARN, de protéines, d'acides nucléiques et de phospholipides (Grillo et Colombatto, 2008; Heppell et Lafontaine, 2008; Nudler et Mironov, 2004; Poiata et al., 2009; Weinberg et al., 2008). Puisque tous les groupements de la molécule sont utilisés dans un large éventail de réactions biochimiques, la SAM est considérée comme un des substrats d'enzyme les plus utilisés après l'adénosine triphosphate (ATP). Certains se plaisent même à décrire la SAM comme étant Vhomme à tout
faire ou le maître de tout de la cellule (Loenen, 2006). De plus, la SAM est un métabolite
pouvant lier des aptamères d'ARN nommés riborégulateurs SAM qui seront décrits plus en détail dans les prochaines pages. Il est intéressant de noter que la S-adénosylhomocystéine (SAH), analogue de la SAM produit à la suite de la donation du groupe méthyl de la SAM, est aussi un métabolite ayant la capacité de lier des riborégulateurs (Figure 1.5) (Poiata et al., 2009; Wang et Breaker, 2008).
Figure 1.5. Structures chimiques de la 5-adénosylméthionine (SAM) et de la
S-adénosyl-homocystéine. Le groupement méthyle (CH3) sur l'atome de soufre (S) est ce qui
différencie les deux molécules. Adaptée et modifiée de Wang et Breaker (2008).
SAM SAH
D.2. Riborégulateurs SAM
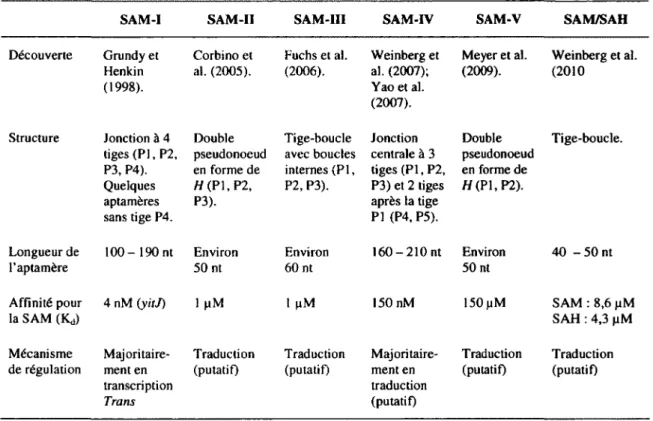
C'est en 1998 que la première séquence et structure bidimentionnelle de riborégulateur SAM a été découverte. Puisqu'à ce moment, le concept de riborégulateur n'était pas connu, ce motif d'ARN a été nommé S-box (Grundy et Henkin, 1998). En effet, ce n'est que cinq années plus tard qu'il a été renommé riborégulateur SAM (Winkler et al., 2003). Jusqu'à ce jour, cinq différents motifs ont été identifiés et confirmés expérimentalement comme étant des riborégulateurs SAM. Ces cinq sous-classes de riborégulateurs SAM ont été nommées SAM-I, SAM-II, SAM-III, SAM-IV et SAM-V selon la chronologie de leur découverte. De plus, très récemment, une nouvelle classe de riborégulateur SAM/SAH a été découverte lors d'une étude génomique comparative effectuée par le groupe de recherche de Ronald R. Breaker (Weinberg et al., 2010).
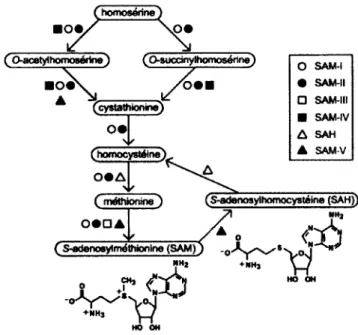
Les riborégulateurs SAM sont principalement retrouvés chez des bactéries à Gram positif (Barrick et Breaker, 2007). Les gènes qu'ils régulent sont reliés au métabolisme du soufre, incluant des gènes impliqués dans la biosynthèse de la cystéine, de la méthionine, de la SAM et de la SAH (Heppell et Lafontaine, 2008; Poiata et al., 2009). Chez plusieurs organismes, une méthionine synthase convertie l'homocystéine en méthionine qui sera convertit à son tour en S-adénosylméthionine par une SAM synthase, aussi nommée méthionine adenosyhransférase (MAT) (Figure 1.6) (Grundy erHenkiir, 1998; Loenen, 2006).
r homoséfine J IOt> ( Q-acetylhomo»érine) Ç O-sucdnylhomosérine) Oi (cystathionine) (homocystéine) Ç méthionine) o SAM-I
•
SAM-II•
SAM-III•
SAM-IV A SAH • SAM-V ( S-adeno«y1méthionirte (SAM) j i -oV^Vj CS-adenoaylhomocystéine (SAH)) NH, A » <yS o'*Y^'sV,oJ + «Hj \ 7 HO OHH
HO OHFigure 1.6. Sous-classes de riborégulateurs impliquées dans la régulation de la
biosynthèse de la S-adénosylméthionine (SAM) et de la S-adénosylhomocystéine (SAH). Il est à noter que la dernière sous-classe de riborégulateurs SAM, soit le SAM/SAH n'est pas représentée sur cette figure. Adaptée et modifiée de Wang et Breaker (2008) avec ajout d'informations tirées de Sowell et al (2008).
D.2.1. Riborégulateur SAM-I
Comme son nom l'indique, le SAM-I est le tout premier motif de riborégulateur SAM découvert (Grundy et Henkin, 1998). Il s'agit aujourd'hui de la sous-classe de riborégulateur SAM la plus étudiée d'entre toutes (Barrick et Breaker, 2007; Poiata et al., 2009). Le motif SAM-I est le plus répandu de tous les riborégulateurs SAM (tableau 1.5). En effet, il est retrouvé dans plus de 250 souches bactériennes (Weinberg et al., 2008).
Tableau 1.5. Distribution phylogénétique des sous-classes de riborégulateurs SAM
SAM-I SAM-II SAM-III SAM-IV SAM-V SAM/SAH
Acidobactéries X Actinomycètes X X Bactéroïdes X X X Chlorobi X Chloroflexi X Cyanobactéries X Deinococcus-Thermus X Fusobactéries X Firmicutes Bacillales X Clostridia X Lactobacilles X X Mollicutes X Proteobacteria a-proteobactéries X X X X X P-proteobactéries X 8/E- proteobactéries X y- proteobactéries X X
Adapté et modifié de Wang et Breaker (2008) (2010); Poiata et al. (2009).
avec ajout d'informations de Weinberg et al.
La structure cristalline du riborégulateur SAM-I de la bactérie Thermoanaerobacîer
tengcongensis en présence de SAM a été résolue pour la première fois en 2006 (Montange et
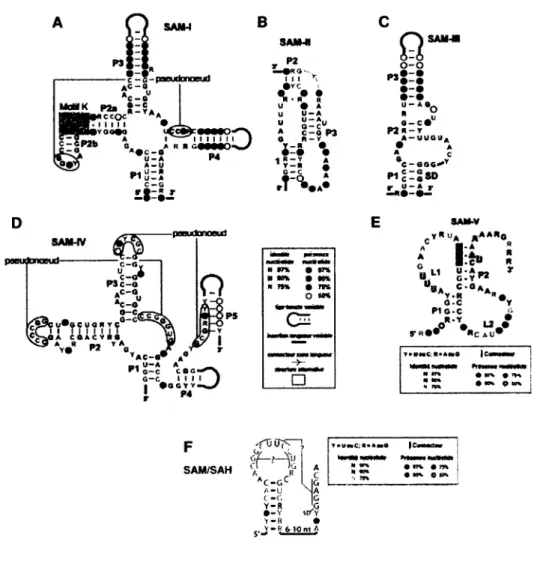
Batey, 2006). Cela a permis de mieux comprendre l'arrangement spatial qu'occupent les quatre tiges (PI, P2, P3, P4) de l'aptamère toutes reliées autour d'une jonction centrale nommée cœur (Figure 1.7A). La longueur de l'aptamère du riborégulateur SAM-I peut varier de 100 à 190 nucléotides à cause de l'énorme variation observée à l'extrémité de la tige P3.
SANM - ?—ff- paeudonoeud A*-® B "C-0U W» S-ÏA ce oc A i i 111 >YOOfQ
»?
*#C-0 U-A A-U WU-A U-* C-Go
P4 pseudonoeud SANMI y ? —#RO-i —#RO-i t \ #YC Î u • A U U-A Il ""* Ay o— c S g:?" i:i *• 1 *-* A 1 Y-« 1 G — C T N V7% c-o O *o% OCUONT U-AQ
O-O SAM-M P2*-* a-uuoi»a «MjlgSD r O - A y SAM-V Y ^ U A « A A R JI
:«• -* s " ,» S:$« * ®Av.S °**b« o-c y o-c Pi o-c .«•V y « _ « C t-2 A U T»*MC;l*A«0 • «r% # • »* O <0* F SAM/SAH -roffrrn, GÏ— >-4ÎI cl ' JG A \ r AC-CC L IG c A-U N A ( - G G y-R \G •-Y st/¥ V-R • y»R 6-10nt A » rr% « m I wv o mFigure 1.7. Séquences et structures consensus des riborégulateurs SAM. Chacune de ces cinq sous-classes (A) SAM-I, (B), SAM-II, (C) SAM-III ou SAMMK? (D) SAM-IV, (E) SAM-V et (F) SAM/SAH possède une séquence et une structure consensus propre. Les figures (A) à (D) partagent la même légende (à droite de la figure (D)), tandis que les figures (E) et (F) possèdent leurs propres légendes. Adaptée de Wang et Breaker (2008), de Poiata et al. (2009) et de Weinberg et al. (2010).
D'entre toutes les sous-classes de riborégulateurs SAM, le SAM-I est celui qui a la meilleure affinité pour son ligand. En effet, sa constante de dissociation (K<j) pour la SAM est d'environ 4 nM (Winkler et al., 2003). De manière moins efficace, le riborégulateur SAM-I peut aussi lier des analogues de la SAM, comme la SAH. La grande majorité des riborégulateurs SAM-I semblent fonctionner grâce à un mécanisme de régulation transcriptionnelle (McDaniel et al., 2006; Poiata et al., 2009; Winkler et al., 2003). En effet, selon une analyse de séquences, la plupart de ces aptamères seraient suivis d'une tige terminatrice indépendante de Rho. Cependant, certains riborégulateurs SAM-I possèdent une plateforme d'expression ambiguë, c'est-à-dire où aucune tige terminatrice ne peut être prédite (Winkler et al., 2003). De plus, en 2008, il a été démontré que le riborégulateur SAM-I de Clostridium acetobutylicum agissait de façon antisens sur le contrôle l'expression d'ARN de l'opéron ubiG impliqué dans le métabolisme de la méthionine (André et al., 2008). Un an plus tard, une autre équipe démontra qu'un autre riborégulateur SAM-I pouvait aussi agir de façon antisens sur la régulation d'un gène de virulence chez Listeria monocytogenes (Loh et al., 2009).
D.2.2. Riborégulateur SAM-II
C'est en 2005 que le deuxième motif de riborégulateur SAM, le SAM-II, a été identifié lors d'une étude comparative de séquences de régions intergéniques chez des a-protéobactéries (Tableau 1.5) (Corbino et al., 2005; Poiata et al., 2009). Bien qu'il soit majoritairement retrouvé chez des protéobactéries, le riborégulateur SAM-II est aussi présent chez des bactéroïdes (Poiata et al., 2009). Sa structure secondaire est illustrée à la figure I.7B, il s'agit d'un double pseudonoeud arrangé en forme de H (Haller et al., 2011). Jusqu'à la découverte du riborégulateur SAM-V, possédant une structure secondaire très semblable, le riborégulateur SAM-II était le plus petit riborégulateur SAM connu, comportant seulement une cinquantaine de nucléotides (Corbino et al., 2005). La structure cristalline de ce riborégulateur a été résolue en 2008 (Gilbert et al., 2008).
Selon Haller et al. (2011), le riborégulateur SAM-II agirait sur l'expression génétique via un mécanisme de régulation traductionnelle. L'aptamère du riborégulateur SAM-II possède une affinité pour la SAM 250 fois plus faible que celle du riborégulateur SAM-I. En effet, il possède un K<j d'environ 1 pM (Corbino et al., 2005; Ham et al., 2011). De plus, il est intéressant de noter que la SAM adopte une conformation différente qui dépend du riborégulateur auquel elle est liée. En effet, sa conformation est plus compacte lorsqu'elle est liée au riborégulateur SAM-I que lorsque cette liaison se fait avec un riborégulateur SAM-II (Gilbert et al., 2008; Montange et Batey, 2006; Poiata et al., 2009). Le riborégulateur SAM-II lie la SAM, mais aussi la SAH et d'autres analogues avec une affinité beaucoup plus faible (Lim et al., 2006). Certains analogues de la SAM ont été testés dans le but de créer de nouveaux antibiotiques (Ham et al., 2011).
D.2.3. Riborégulateur SAM-III ou SAMMK
Un an après la découverte du riborégulateur SAM-II, soit en 2006, le premier représentant des riborégulateurs SAM-III a été identifié grâce à l'étude de régions 5' non traduites situées sur l'ARNm de gènes metK codant pour des SAM synthases chez plusieurs organismes (Fuchs et al., 2006). Déjà, à ce moment, le gène metK était connu pour être sous le contrôle de riborégulateurs SAM chez différentes espèces. Le riborégulateur SAM-III est principalement retrouvé chez des bactéries de l'ordre desjactobacilles (Tableau 1.5) (Fuchs et al., 2006; Poiata et al., 2009). Tout comme le riborégulateur SAM-II, la structure cristalline de ce riborégulateur a été résolue en 2008 (Lu et al., 2008). Bien entendu, le riborégulateur SAM-III possède une structure bien distincte des autres riborégulateurs SAM déjà identifiés (Poiata et al., 2009). En effet, sa structure est une des plus simples dans le monde des riborégulateurs (Figure I.7C). Il s'agit simplement d'une tige-boucle d'environ soixante nucléotides contenant deux boucles internes.