UNIVERSITE MOHAMMED V- Agdal FACULTE DES SCIENCES
RABAT
THESE DE DOCTORAT D’ETAT
Présentée par
Amina REGRAGUI
Discipline : Biologie
Spécialité : Phytopathologie
Contribution à l’étude de l’influence de la salinité sur
le couple tomate –Verticillium : Conséquences
physiologiques et impact sur la bioprotection des
tomates contre la verticilliose
Soutenue le…22 décembre 2005…..devant le jury :LAHLOU H.
Professeur à la Faculté des Sciences de Rabat Présidente BENKHEMMAR O.
Professeur à la Faculté des Sciences de Rabat Examinateur FASSI FIHRI O.
Professeur à l’I.A.V. Hassan II, Rabat Examinateur TIJANE M.
Professeur à la Faculté des Sciences de Rabat Examinateur ZAID H.
JE DEDIE CE TRAVAIL…
A mes petits enfants
Zineb
Youssef
Jad
REMERCIEMENTS
Ce travail a été réalisé au sein de l’équipe de phytopathologie du laboratoire de Botanique de la Faculté des Sciences de Rabat. Qu’il me soit permis ici de remercier vivement le professeur H. LAHLOU pour la confiance qu’elle m’ a toujours témoignée ainsi que la formation de chercheur qu’elle m’a donnée avec bienveillance et humanité. Ses qualités scientifiques, ses conseils et ses encouragements incessants m’ont beaucoup soutenue pendant la l’élaboration de ce travail. Je lui exprime ma profonde reconnaissance et lui porte mon grand respect.
Je remercie très vivement le professeur H. ZAID pour sa disponibilité, ses conseils fructueux et son aide scientifique et matérielle. Il a bien voulu accepter de participer à ce jury de thèse en qualité de rapporteur. Qu’il trouve ici l’’expression de ma profonde gratitude.
Madame O. FASSI FIHRI, professeur de microbiologie à l’’IAV HASSAN II, m’a agréablement accueillie dans son laboratoire. J’ai pu bénéficier de ses compétences scientifiques et de son savoir faire. Pour l’accueil chaleureux qu’elle m’a réservé et pour avoir accepté de faire le rapport de ce mémoire, je lui exprime mes plus vifs remerciements.
Mes sincères remerciements sont adressés aux professeurs M. TIJANE et O. BENKHEMMAR pour l’’honneur qu’ils me font de juger cette thèse.
Je désire remercier particulièrement mes collègues les professeurs A. EL AISSAMI, E. BERRAHO et M. RAHOUTI pour leur collaboration efficace.
Je désire remercier également Madame AGOUMY, ingénieur au ministère de l’énergie et des mines pour son aide précieuse lors de la réalisation de l’analyse minérale des échantillons.
Que mes collègues du laboratoire de Botanique qui m’ont apportés une aide morale et matérielle trouvent ici l’’expression de ma meilleure sympathie.
Je remercie vivement M. BENCHAKRI pour son aide technique efficace et son dévouement.
Enfin, mes remerciements vont à tous les membres de ma famille pour leur soutien moral et leur encouragement affectueux.
Liste des publications
1.
REGRAGUI A., LAHLOU H. et ZAID H., 1989. La prémunition de la tomate contre la verticilliose causée par Verticillium albo-atrum, forme à microsclérotes. Conséquences physiologiques du phénomène.Cryptogamie, Mycol. 10 (3) : 243-256
2.
REGRAGUI A. ZAID H. and LAHLOU H., 1990. Verticilliumalbo-atrum effect on nitrate reductase activity in young tomato plants.
Proceeding of 8th congress of the Mediterranean Phytopathological Union, Agadir (Morocco) October 28th ,233-235.
3.
BALESDENT M.H., REGRAGUI A., LAHLOU H. and BOMPEIX, 1990. Early diagnostic of Verticillium albo-atrum microsclerotia form, and inoculum evaluation by enzyme linked immunosorbent assay in infected tomato plants.Proceeding of 8th congress of the Mediterranean Phytopathological Union, Agadir (Morocco) October 28th , 75-77.
4.
REGRAGUI A., RAHOUTI M. et LAHLOU H., 2003. Effet du stress salin sur Verticillium albo-atrum: pathogénécité et production d’enzymes cellulolytiques in vitroCryptogamie, Mycol. 24 (2) : 167-174.
5.
REGRAGUI A. and LAHLOU H., 2005. Effect of Salinity on in vitroTrichoderma harzianum Antagonism against Verticillium dahliae. Pakistan Journal of Biological Sciences 8 (6):872-876
SOMMAIRE
INTRODUCTION GENERALE ... 1
CHAPITRE I : ETUDE DE L’INFLUENCE DE LA SALINITE SUR LE COMPORTEMENT DE LA TOMATE (LYCOPERSICON ESCULENTUM) ET SUR LE DEVELOPPEMENT IN VITRO DE VERTICILLIUM ALBO-ATRUM ... 7
I. EFFET DE LA SALINITE SUR LE COMPORTEMENT DE DEUX GENOTYPES DE TOMATE... 7
Introduction... 7
A. Matériel et méthodes ... 9
1. Le matériel végétal... 9
2. Effet du sel sur la germination des graines ... 9
2.1. Protocole de germination... 9
2.2. Conditions de culture... 9
2.3. Observations... 10
3. Effet du sel sur la croissance végétative des plantes... 10
3.1. Conditions de culture... 10
3.2. Lecture des résultats ... 11
4. Analyse statistique des données... 11
B. Résultats ... 11
1. Action du sel sur le pouvoir germinatif des graines... 11
1.1. Effet sur la vitesse de germination ... 11
1.2. Effet sur le taux de germination ... 13
2. Action du sel sur la croissance végétative des plantes... 15
2.1. Effet sur la taille de l’axe aérien... 15
2.2. Effet sur la biomasse des parties aériennes ... 16
C. Discussion et conclusion ... 18
II. INFLUENCE DE LA SALINITE DU MILIEU SUR LE DEVELOPPEMENT IN VITRO D’UN ISOLAT DE VERTICILLIUM ALBO-ATRUM D’ORIGINE TOMATE... 19
Introduction... 19
A. Matériel et méthodes ... 21
1. La souche fongique... 21
2. Ensemencement ... 22
3. Traitement par le sel ... 22
4. Estimation de l’effet du sel sur la croissance de Verticillium ... 23
4.1. Croissance diamétrale... 23
4.2. Croissance pondérale... 23
5. Effet sur l’intensité de la conidiogénèse ... 23
6. Effet sur le pouvoir germinatif des conidies ... 24
7. Effet sur les organes de résistance ... 24
B. Résultats ... 24
1. Effet de l’apport de NaCl sur la croissance de Verticillium... 24
1.1. Croissance diamétrale... 24
1.2. Croissance pondérale... 25
2. Effet de la salinité sur la conidiogénèse et le pouvoir germinatif des conidies ... 25
2.2. Pouvoir germinatif des conidies ... 28
3. Effet de la salinité sur la sclérogénèse ... 28
4. Influence de l’interaction salinité-température sur la croissance de Verticillium ... 31
C. Discussion et conclusion ... 32
CHAPITRE 2 : ETUDE DES EFFETS DE LA SALINITE SUR LA RELATION HOTE-PARASITE DANS LE CAS DE LA VERTICILLIOSE ... 38
INTRODUCTION... 38
I. EFFET DE L’INTERACTION SALINITE-VERTICILLIUM SUR LA MANIFESTATION DE LA MALADIE CHEZ LA TOMATE... 39
A. Matériel et méthodes ... 39
1. Plantes hôtes et conditions de culture ... 39
2. Inoculation des plantules... 39
2.1. L’agent pathogène : Verticillium albo-atrum, forme à microsclérotes... 39
2.2. Préparation de l’inoculum ... 40
2.3. Protocole d’inoculation ... 40
3. Traitement par le sel ... 40
4. Notations des résultats ... 40
4.1. Croissance des plantes... 40
4.2. Emission des feuilles ... 41
4.3. Colonisation des plantes par le pathogène... 41
4.4. Analyse statistique des résultats ... 42
B. Résultats ... 42
1. Manifestations externes de la combinaison Salinité-Verticillium sur les plantes de tomate hôtes ... 42
1.1. Effet sur la croissance des plantes ... 42
1.2. Effet sur l’émission de feuilles ... 45
2. Manifestations internes ... 48
C. Discussion et conclusion ... 49
II. EFFET DE LA SALINITE SUR LA VARIABILITE DU POUVOIR PATHOGENE DE VERTICILLIUM ALBO-ATRUM... 51
Introduction... 51
A. Impact de la salinité sur le niveau infectieux de l’inoculum du Verticillium sur les plantes de tomate... 53
1. Matériel et méthodes... 53
1.1. Inoculation... 53
1.2. Traitement par le sel et conditions de culture... 54
1.3. Estimation des résultats ... 54
2. Résultats : effet de la salinité sur le pouvoir infectieux de l’inoculum de Verticillium vis-à-vis de la tomate ... 54
2.1. Effet sur la tomate Marmande Claudia... 54
2.2. Effet sur la tomate Marmande VR... 56
3. Discussion et conclusion... 56
B. Influence de la salinité sur l’expression du pouvoir pathogène de deux isolats de Verticillium sur la tomate ... 59
1.1. Test de pathogénécité ... 59
1.2. Lecture des résultats ... 60
2. Résultats... 60
3. Discussion et conclusion... 60
C. Influence de la salinité sur la variabilité de la spécificité parasitaire d’un isolat de Verticillium d’origine tomate ... 63
1. Matériel et méthodes... 63
1.1. Choix des plantes hôtes inhabituelles... 63
1.2. Inoculation et traitement par le sel ... 63
1.3. Lecture des résultats ... 64
2. Effet de la salinité sur la variation du pouvoir pathogène de l’isolat P80 au contact de nouvelles plantes hôtes. ... 64
2.1. Effet sur l’expression du pouvoir pathogène au contact de la luzerne ... 64
2.2. Effet sur l’expression du pouvoir pathogène au contact de l’aubergine... 65
3. Discussion et conclusion... 68
CHAPITRE 3 : CONSEQUENCES PHYSIOLOGIQUES ET BIOCHIMIQUES DE L’INTERACTION SALINITE-VERTICILLIUM CHEZ LA TOMATE ... 73
INTRODUCTION... 73
A. EFFETS SUR LES TENEURS EN CATIONS MAJEURS... 74
1. Introduction... 74
2. Matériel et méthodes... 76
3. Résultats... 76
3.1. Teneurs en ions sodium ... 76
3.2. Teneurs en ions potassium... 78
4. Discussion et conclusions ... 78
B. EFFETS SUR LES TAUX DE SUCRES SOLUBLES TOTAUX... 81
1. Introduction... 81
2. Matériel et méthodes... 82
2.1. Extraction des sucres solubles totaux ... 83
2.2. Réaction à l’anthrone... 83
3. Résultats... 83
3.1. Chez Marmande Claudia ... 83
3.2. Chez Marmande VR ... 84
4. Discussion et conclusion... 84
C. EFFETS SUR L’ACCUMULATION DE LA PROLINE... 87
1. Introduction... 87
2. Matériel et méthodes... 88
2.1. Extraction ... 88
2.2. Dosage ... 88
3. Résultats... 89
3.1. Chez la variété Marmande Claudia ... 89
3.2.Chez la variété Marmande VR ... 89
4. Discussion et conclusion... 89
D. EFFETS DE L’INTERACTION SALINITE-VERTICILLIUM SUR CERTAINES ACTIVITES ENZYMATIQUES DES PLANTES DE TOMATE... 93
1. Effets sur l’activité nitrate réductase... 93
1.2. Matériel et méthodes ... 94
1.3. Résultats ... 95
1.4. Discussion et conclusion ... 95
2. Effets sur l’activité peroxydasique... 98
2.1. Introduction ... 98
2.2. Matériel et méthodes ... 100
2.3. Résultats ... 100
2.4. Discussion et conclusion ... 103
CHAPITRE 4 : IMPACT DE LA SALINITE DU MILIEU SUR LES MECANISMES D’ACTION DE VERTICILLIUM ALBO-ATRUM... 107
I. EFFET DE LA SALINITE SUR LA TOXINOGENESE DE L’AGENT PATHOGENE... 107
Introduction... 107
A. Matériel et méthodes ... 108
1. Préparation des filtrats de culture de l’isolat P80 de Verticillium... 108
2. Mise en évidence de la toxicité des filtrats de culture de Verticillium par des tests biologiques... 109
2.1. Test graine ... 109
2.2. Test plante entière ... 110
B. Résultats ... 110
1. Effet du NaCl sur la phytotoxicité des filtrats de culture de l’isolat P80 de Verticillium sur la croissance racinaire de deux espèces végétales... 110
1.1. Manifestation sur la tomate ... 110
1.2. Manifestation sur le cresson ... 111
2. Effet du NaCl sur la phytotoxicité des filtrats de culture de l’isolat P80 de Verticillium sur la croissance végétative de deux plantes hôtes ... 111
2.1. Manifestation sur la tomate Claudia... 111
2.2. Manifestation sur l’aubergine Reina negra... 113
C. Discussion et conclusion ... 113
II. INFLUENCE DE LA SALINITE SUR L’ACTIVITE DES ENZYMES PECTOCELLULOSIQUES PRODUITES IN VITRO PAR VERTICILLIUM ALBO-ATRUM... 116
Introduction... 116
A. Matériel et méthodes ... 117
1. Les isolats de Verticillium utilisés ... 117
2. Analyse de l’activité carboxyméthylcellulase... 117
2.1. Production de l’enzyme... 117
2.2. Révélation... 118
3. Analyse de l’activité pectine méthyle estérase ... 119
3.1. Production de l’enzyme... 119
3.2. Révélation... 119
B. Résultats ... 120
1. Croissance de deux isolats de Verticillium développés sur milieu CMC enrichi en NaCl... 120
2. Effet de la salinité sur l’activité carboxyméthylcellulase ... 120
3. Effet de la salinité sur l’activité pectine méthyle estérase ... 120
III. INFLUENCE DE LA SALINITE SUR L’ACTIVITE PHENOLOXYDASIQUE DE VERTICILLIUM
ALBO-ATRUM... 124
Introduction... 124
A. Matériel et méthodes ... 125
1. Les souches fongiques ... 125
2. Détection des phénoloxydases. ... 126
1.2. Les réactifs ... 126
2.2. Révélations ... 126
B. Résultats ... 126
1. Influence de la salinité sur la cinétique de la production des Pox ... 126
1.1. La souche sauvage P 80... 127
1.2. La souche hyaline P1... 127
2. Influence de la salinité sur l’activité phénoloxydasique globale de Verticillium... 127
C. Discussion et conclusion ... 130
IV. EFFET DE LA SALINITE DU MILIEU SUR LE PROFIL PROTEIQUE DE L’ISOLAT P80 DE VERTICILLIUM ALBO-ATRUM... 131
Introduction... 131
A. Matériel et méthodes ... 132
1. Concentration des protéines... 132
2. Dosage des protéines ... 132
3. Analyse des protéines par électrophorèse ... 133
3.1. Préparation des gels... 133
3.2. Préparation des échantillons ... 133
3.3. Migration ... 133
3.4. Révélation... 133
3.5. Lecture des résultats ... 134
B. Résultats ... 134
1. Effet de la salinité sur le taux des protéines secrétées dans les filtrats de culture par Verticillium. ... 134
2. Effet de la salinité sur l’évolution du pH des filtrats de culture... 135
3. Effet de la salinité sur les profils protéiques de l’isolat P80... 136
3.1. Analyse des protéines sur gel de polyacrylamide... 136
3.2. Comparaison de l’expression des protéines... 137
C. Discussion et conclusion ... 139
CHAPITRE 5 : INFLUENCE DE LA SALINITE SUR L’ANTAGONISME MICROBIEN VIS-A-VIS DE VERTICILLIUM ALBO-ATRUM ET SUR LA BIOPROTECTION DES TOMATES CONTRE LA VERTICILLIOSE ... 141
INTRODUCTION... 141
I. EFFET DE LA SALINITE SUR L’ANTAGONISME IN VITRO DE QUELQUES MICROORGANISMES BENEFIQUES DU SOL VIS-A-VIS DE VERTICILLIUM ALBO-ATRUM... 144
Introduction... 144
A. Matériel et méthodes ... 145
1. Choix des microorganismes antagonistes ... 145
1.1. Penicillium sp ... 145
1.3. Talaromyces flavus... 146
1.4. Trichoderma harzianum... 146
1.5. Bacillus sp ... 146
2. Protocole des cultures en confrontations ... 146
3. Estimation de l’effet antagoniste ... 146
B. Résultats ... 147
1. Effets de la salinité sur la compétitivité in vitro de quelques microorganismes fongiques avec Verticillium ... 147
1.1. Confrontations Penicillium sp-Verticillium ... 147
1.2. Confrontation Fusarium oxysporum-Verticillium... 149
1.3. Confrontation Talaromyces flavus-Verticillium ... 149
1.4. Confrontation Trichoderma-Verticillium ... 149
2. Effet de la salinité sur l’antagonisme de Bacillus sp avec Verticillium ... 153
C. Discussion et conclusion ... 153
II. INFLUENCE DE LA SALINITE SUR L’ANTAGONISME IN VITRO DE TRICHODERMA HARZIANUM VIS-A-VIS DE L’ISOLAT P80 DE VERTICILLIUM... 157
Introduction... 157
A. Matériel et méthodes ... 158
1. Le matériel fongique... 158
1.1. Le pathogène ... 158
1.2. L’antagoniste ... 158
2. Mesure du phénomène d’antagonisme... 159
2.1. Capacité de colonisation... 159
2.2. Antagonisme par antibiose ... 159
B. Résultats ... 161
1. Influence de la salinité du milieu sur le développement de T. harzianum... 161
1.1. Morphologie des cultures ... 161
1.2. Poids sec du mycélium ... 161
1.3. Taux de sporulation ... 161
2. Influence de la salinité sur l’expression de l’activité antagoniste de T. harzianum vis-à-vis de Verticillium... 161
2.1. Antagonisme par compétition... 161
2.2. Antagonisme par antibiose ... 164
C. Discussion et conclusion ... 170
III. INFLUENCE DE LA SALINITE SUR L’EXPRESSION DE L’ANTAGONISME IN VIVO DE TRICHODERMA HARZIANUM VIS-A-VIS DE VERTICILLIUM ALBO-ATRUM DE LA TOMATE... 172
Introduction... 172
A. Matériel et méthodes ... 173
1. Protocole de bioprotection des plantes ... 173
1.1. Préinoculation avec Trichoderma... 173
1.2. Inoculation avec la souche pathogène ... 174
2. Evaluation des manifestations de la maladie ... 174
2.1. Croissance des plantes... 174
2.2. Altérations foliaires ... 174
2.3. Ré-isolement du champignon ... 175
B. Résultats ... 175
1. Recherches des conditions optimales pour l’expression de l’antagonisme in vivo de Trichoderma harzianum vis-à-vis de Verticillium albo-atrum. ... 175
1.1. Effet de l’inoculation simultanée par le mélange de spores des deux champignons 1.2. Effet du délai de pré-inoculation par Trichoderma sur l’expression de la
bioprotection... 178
1.2. Discussion et conclusion ... 181
2. Impact de la salinité sur le succès de la bioprotection des tomates contre la verticilliose par Trichoderma harzianum... 184
2.1. Réponses des plantes prétraitées avec Trichoderma à la surinfection par Verticillium en fonction de la salinité du milieu ... 184
2.2. Discussion et conclusion ... 186
CONCLUSION GENERALE ... 189
REFERENCES BIBLIOGRAPHIQUES... 200
Introduction générale
La culture de la tomate Lycopersicon esculentum Mill, a connu une grande extension au Maroc. La superficie globale occupée par les cultures de primeurs et de saison s’élève à 20655 ha en 2001/02 et assure une progression de 18.7% par rapport à 2000/01. Cette progression concerne aussi bien la tomate sous serre que celle de plein champ. La production globale est passée de 307000 tonnes en 1989 à 560000 tonnes en 1999 (S.A.M., 2002).
Cependant cette culture est confrontée à de nombreuses contraintes qui affectent aussi bien le rendement que la qualité des fruits. Ces contraintes sont liées à des changements dans l’environnement de la plante, notamment la température et la salinité, et au développement des maladies.
La culture de la tomate a dominé depuis les quatre dernières décennies les régions du littoral atlantique, régions où règnent une humidité relative élevée et des températures hivernales douces. Or, la proximité de l'océan atlantique fait que cette culture subit la salinité des eaux d'arrosage puisées de la nappe phréatique salée et les embruns qui entraînent une importante accumulation de sel dans le sol (Besri, 1981). Par ailleurs, ces conditions semblent propices au développement des maladies vasculaires dues principalement aux champignons du genre Verticillium. La verticilliose de la tomate due à Verticillium albo-atrum, forme à microsclérotes représente actuellement l’une des principales maladies de cette culture au Maroc. Elle se manifeste par un rabougrissement des parties aériennes, des altérations foliaires et des flétrissements conduisant parfois à la mort de la plante. L’agent pathogène est un adélomycète vasculaire qui se conserve dans le sol sous forme de microsclérotes. Ces propagules dormantes se développent sur les débris végétaux au cours de la vie saprophytique du champignon et représentent l’inoculum de la maladie. En présence de la plante hôte, les microsclérotes germent en réponse aux exsudats racinaires et pénètrent dans les racines même en absence de toute blessure. Le parasite se développe et colonise la totalité de la plante puis reste localisé à l’intérieur des vaisseaux conducteurs provoquant leur obstruction et par voie de conséquence le flétrissement de la plante infectée.
La colonisation des tissus par le parasite est rendue aisée par la production d’enzymes de dégradation des parois cellulaires de l’hôte (Cooper et al., 1978 ; Durand & Cooper, 1988). Une corrélation positive existe entre l’activité des cellulases et des pectinases produites par des isolats de Verticillium et leur agressivité vis-à-vis de leurs plantes hôtes (Gupta & Heale, 1971, Durand & Cooper, 1988). En outre, ce champignon est apte à produire des métabolites toxiques à l’intérieur des tissus infectés et qui seraient impliqués dans l’expression des symptômes de la maladie (Nachmias et al, 1982 ; 1984 et Meyer et al., 1994).
Il est connu que le comportement des plantes envers les agents pathogènes est conditionné par certains facteurs de l’environnement. Ayres, (1984) a rapporté que les stress abiotiques comme la sécheresse, la pollution, la chaleur ou la salinité peuvent augmenter les symptômes des maladies par un effet direct sur la plante ou sur le pathogène.
Quelques travaux ont montré l’influence de la salinité du milieu sur l’augmentation de la sensibilité de la tomate à la fusariose (Standaert, 1975) et à la verticilliose (Besri & Afaïlal, 1993). Selon ces auteurs, cette plus grande sensibilité est en relation avec une colonisation plus intense des tiges par le parasite.
Le sel semble avoir une action néfaste aussi bien sur la plante hôte que sur l’agent pathogène. Quelques rares hypothèses ont été avancées pour expliquer les mécanismes par lesquels agit le chlorure de sodium pour accentuer la sévérité de ces maladies. Messiaen & Lafon (1971) ont émis l’hypothèse que l’ion sodium augmente la sensibilité de la tomate à la fusariose vasculaire par suite d’une insuffisance en calcium provenant d’une interférence ionique entre ces deux cations. Sur d’autres pathosystèmes, Mac Donald, (1984) et Sulistyowati & Keane, (1992), estiment que la salinité prédispose les plantes aux attaques de
Phytophthora en inhibant la synthèse de phytoalexines, substances intervenant
dans la protection des plantes contre l’invasion par les agents pathogènes. Benyahyia, (1998), travaillant sur le couple Citrus-Phytophthora a montré l’effet spécifique des ions chlore sur l’interaction hôte-parasite. Cet ion agit sur la
physiologie de la plante et prédispose les porte-greffes à des attaques sévères par l’agent pathogène.
La sévérité de la maladie en présence de sel serait liée également à une action directe du sel sur l’agent pathogène. En effet, l’augmentation du chlorure de sodium dans le sol stimule le développement et la conservation des champignons vasculaires (Besri, 1981 ; Afailal, 1987 ). De la même manière, la croissance et la production de sporanges chez Phytophthora citrophthora sont stimulées par la présence de sel dans le sol et dans les eaux d’irrigation (Benyahyia, 1998).
D’autres facteurs biotiques sont incriminés dans l’augmentation de l’incidence des trachéomycoses chez la tomate. On peut citer entre autres l’intervention des nématodes sur la plante hôte et sur l’agent pathogène et l’augmentation de l’inoculum dans le sol (Besri, 1991). Ces facteurs peuvent interagir avec le facteur salinité et accentuer, par conséquent, la sévérité de la maladie.
Si de multiples travaux ont porté sur le rôle du sel dans la prédisposition des plantes aux maladies, peu d’études ont été entreprises pour élucider les mécanismes physiologiques et biochimiques qui régissent le changement de la sensibilité des plantes stressées aux agressions fongiques.
Dans la nature, les relations hôte-parasite-environnement sont très complexes. La réponse des plantes à la fois aux stress abiotique et biotique est la résultante de plusieurs processus physiologiques et métaboliques mis en route pour adapter la plante d’une part aux contraintes hydriques et nutritionnelles et d’autre part, pour élaborer les mécanismes de défense contre les outils pathogéniques de l’agent causal de la maladie.
L’interprétation de l’influence du milieu nutritif sur la sensibilité des plantes à la maladie requiert une connaissance approfondie des caractéristiques physiologiques des plantes stressées par la salinité et des modifications métaboliques liées à l’interaction salinité-pathogène.
Dans cette directive, nous avons successivement :
- Evalué séparément l’effet de la salinité sur le comportement de la plante et sur les activités biologiques in vitro de l’agent pathogène. Ces recherches ont été conduites respectivement sur deux variétés de tomate choisies pour
leur sensibilité à la verticilliose et sur un isolat de Verticillium albo-atrum se montrant agressif sur les deux variétés de tomate testées,
- Montré l’influence de la salinité sur la manifestation de la maladie et sur la variabilité du pouvoir pathogène d’un isolat de Verticillium d’origine tomate.
- Etudié les conséquences physiologiques et biochimiques liées à la salinité en relation avec l’infection par Verticillium chez les tomates doublement stressées et enfin,
- Mis en évidence l’impact de la salinité sur les mécanismes d’action de l’agent pathogène notamment sa capacité à produire les toxines et les enzymes cellulolytiques.
Cette étude s’est appuyée sur la modélisation des interactions hôte-pathogène-environnement représentées par Van Der Plank sous forme d’un triangle ; le triangle de la maladie. Ce modèle peut être développé et inclure l’Homme, ce qui implique une nouvelle schématisation des interactions de la maladie sous forme d’un tétraèdre (Agrios, 1988), l’Homme occupant le sommet de la pyramide. En effet, l’Homme peut influencer les trois autres facteurs en intervenant dans le choix des espèces végétales, le choix des sols et sur les méthodes de lutte.
A ce propos, en raison de la gravité des problèmes posés par la salinité sur le développement de la verticilliose et de la nécessité de mettre au point une stratégie de lutte efficace contre Verticillium dans de telles conditions de l’environnement, nous avons visé, dans la dernière partie de ce travail, d’examiner l’influence de la salinité sur le succès de la bioprotection des tomates contre la verticilliose. Mais pourquoi une lutte biologique ?
En fait, plusieurs méthodes ont été préconisées pour réduire l’incidence de la maladie.
La rotation culturale a montré des résultats intéressants, mais son efficacité est très contestée vu le mode de conservation du champignon dans le sol sous forme de microsclérotes. Ces propagules ont une longue survie et peuvent contaminer des plantes adventices non hôtes, ce qui contribue à l’augmentation du taux d’inoculum dans le sol.
La lutte chimique est largement utilisée. Plusieurs fongicides comme le benlate, le propiconazole et le paclobutrazol ont réussi à diminuer l’incidence de la maladie. Cependant, ces produits très coûteux présentent des inconvénients pour l’Homme et l’environnement auxquels s’ajoute le risque d’apparition de nouveaux pathotypes résistants.
La lutte génétique est l’une des méthodes les plus efficaces pour lutter contre la verticilliose. L’utilisation des variétés résistantes à la race 1 de Verticillium a fait ses preuves pendant plusieurs années. Néanmoins, son efficacité a diminué avec l’apparition d’une nouvelle race, la race 2 (Besri et al. ; 1984 ; Tjamos, 1984), à laquelle aucune variété ne présente une résistance satisfaisante.
La prise en conscience des limites des méthodes chimique et génétique de lutte, considérées un moment comme susceptibles à elles seules de résoudre les problèmes des maladies des plantes, a incité les chercheurs à s’orienter vers la lutte biologique. Ce moyen de lutte met en œuvre des organismes vivants ou des substances biologiques pour réduire les dégâts causés par les agents phytopathogènes.
Le contrôle biologique avec les microorganismes bénéfiques permet d’augmenter le rendement en supprimant directement l’inoculum pathogène et/ou en induisant la résistance des plantes. La résistance induite représente actuellement une nouvelle stratégie de défense des plantes contre les agressions pathogènes.
Plusieurs tentatives de lutte biologique contre la verticilliose ont abouti soit par l’induction de la résistance soit par l’utilisation des microorganismes antagonistes. C’est le cas de la bioprotection de l’aubergine par l’utilisation de champignons antagonistes comme Talaromyces flavus (Fravel, 1996 ; Kim et al., 1988 ) et
Trichoderma harzianum (D’Ercole et al., 2000), celle de l’érable par Bacillus subtilis
(Hall & Schreiber, 1984) et de la luzerne par Sinorhizobium meliloti (El Aissami, 1999). La bioprotection des tomates contre Verticillium a été obtenue par prémunition à la suite de la pré-inoculation des jeunes plants par une souche avirulente du même champignon avant l’infection agressive (Regragui et al., 1989). Cependant, il a été établi que l’efficacité des microorganismes bénéfiques à supprimer les maladies est influencée par les facteurs de l’environnement (Lewis & Papavizas, 1987 ; Harman et al., 1981). Il est donc important de connaître dans la mesure du
possible l’écologie des agents du contrôle biologique et leurs interactions avec le pathogène, la plante hôte et la communauté microbienne de la rhizosphère (Larkin & Fravel, 2002).
A la lumière de ces données, et dans l’objectif de vérifier dans quelle mesure la salinité peut affecter la réussite du phénomène de bioprotection, on s’est proposé de protéger les tomates soumises au stress salin contre la verticilliose par l’utilisation de
Trichoderma harzianum, un puissant agent antagoniste. Cette tentative a été
précédée par des essais conduits in vitro afin d’examiner l’effet de la salinité sur le développement et la reproduction de l’agent de lutte biologique et sur l’efficacité de ses différents modes d’action antagoniste à l’encontre du champignon pathogène.
Chapitre I : Etude de l’influence de la salinité sur le
comportement de la tomate (Lycopersicon esculentum) et
sur le développement in vitro de Verticillium albo-atrum
I. Effet de la salinité sur le comportement de deux génotypes
de tomate
Introduction
La culture de la tomate au Maroc connaît une grande extension en zone irriguée et en zone côtière. Les eaux utilisées pour l’irrigation des parcelles de tomate le long du littoral atlantique appartiennent aux classes C3 (0.48 g/l – 1.44 g/l) et C4 ( 1.4 g/l – 3.2 g/l) définies par Richards (1969). Ce sont donc des eaux salées à très salées (Amor, 1991). Il en résulte une accumulation de sel dans le sol en plus des embruns qui apportent de grandes quantités de NaCl (Besri, 1977). Lorsque la salinité est de 2.5 g/l, le rendement baisse de 10%. Cependant, la baisse du rendement peut atteindre 25% à une salinité de 4g/l.
La salinité des sols et des eaux d’irrigation ne constituent pas un facteur limitant la culture de la tomate mais peut constituer un facteur qui limite la qualité de la production. En effet, l’impact de la salinité est plus grave sur le rendement export suite à la réduction du calibre des fruits ( PNTTA, 1999)
La salinité a été rapportée comme un facteur de l’environnement qui augmente la sensibilité des tomates aux maladies d’origine fongique principalement la fusariose (Standaert, 1978), la verticilliose (Afailal 1987, Besri, 1990) et la pourriture racinaire due à Phytophthora parasitica (Swiecki & Mac Donald, 1991).
Cette différence de sensibilité induite par le sel serait la résultante de l’interaction du milieu nutritif avec la physiologie de la plante d’une part, et celle du parasite d’autre part. Toute interprétation de l’augmentation de l’incidence de la maladie dans ce cas requiert une bonne connaissance du comportement de la plante vis-à-vis de la salinité du milieu.
Dans cette optique, nous avons évalué le niveau de tolérance vis-à-vis du stress salin de deux variétés de tomate choisies pour leur grande sensibilité à la verticilliose, principal handicap de cette culture au Maroc.
La tolérance d’une plante à la salinité est définie par son aptitude à se développer normalement en conditions salines. Le degré de tolérance dépend du stade physiologique de la plante. En général, les plantes sont plus sensibles au stade germination et émergence. Le degré de sensibilité diminue ensuite avec l’âge.
La résistance des plantes à la salinité dépendrait de leur capacité à maintenir des conditions favorables à leur fonctionnement en évitant ou en tolérant les fortes concentrations ioniques à l’intérieur de leurs cellules. Les différences entre les espèces résident dans l’efficacité des moyens mis en œuvre pour la protection de leur équilibre (Hamza, 1980).
La tolérance de la tomate à la salinité a fait l’objet de plusieurs études ( Shalhevet & Yaron, 1973 ; Perez-alfocea et al., 1996 ; Katerji et al., 1998). D’après ces auteurs, la tomate est moyennement tolérante à la salinité. Cependant, cette tolérance varie d’une espèce à l’autre et même entre les variétés d’une même espèce (Cruz & Cuartero, 1990). Des différences de sensibilité au sel ont été constatées entre différentes variétés de tomate appartenant à l’espèce cultivée Lycopersicon
esculentum et entre les espèces sauvages de tomate aussi bien in vitro qu’in vivo
(Amor, 1991).
Plusieurs critères sont utilisés pour tester la tolérance à la salinité des tomates en phase de croissance végétative. Cruz & Cuartero (1990) ont testé la tolérance de 39 génotypes appartenant à 5 espèces de Lycopersicon en évaluant 16 caractères différents. Ces auteurs recommandent d’utiliser quatre caractères fiables ; la hauteur des tiges, le poids frais des tiges et les concentrations foliaires en Na+ et en Cl-. Ces caractères changent avec la concentration saline indépendamment des génotypes.
C’est ainsi que nous avons testé la tolérance au sel des deux variétés de tomate lors de la germination et du développement végétatif. Celui ci a été évalué par la hauteur des plantes et le poids frais des parties aériennes. Les teneurs en éléments minéraux qui caractérisent hautement la tolérance à la salinité seront étudiées dans le prochain chapitre parmi les effets physiologiques et biochimiques du sel sur le couple tomate-Verticillium.
A. Matériel et méthodes
1. Le matériel végétal
La tomate (Lycopersicon esculentum, Mill.) est une plante maraîchère largement cultivée au Maroc. Deux variétés ont été retenues pour notre étude ; la Marmande Claudia sensible à la verticilliose et la Marmande VR possédant le gène Ve de la résistance à la race 1 de Verticillium, mais sensible à la race 2 du même champignon. Les semences nous ont été gracieusement fournies par " Semences Clauses " de Brétiny (France).
2. Effet du sel sur la germination des graines
2.1. Protocole de germination
Les graines des deux variétés de tomate sont stérilisées par trempages successifs dans les bains suivants :
- Hypochlorite de sodium à 15% pendant 2 minutes - Deux rinçages à l'eau distillée stérile
- Alcool 90° pendant une minute
Les graines sont séchées sur papier filtre stérile avant d'être déposées dans des boîtes de Pétri contenant deux couches de papier filtre stérile imbibé d'une solution saline à différentes concentrations :0 ; 20 ; 40 ; 60 ; 80 et 100 mM de NaCl soit 0 ; 1.16 ; 2.36 ; 3.50 ; 4.67 et 5.84 g/l. Les lots témoins sont placés sur papier filtre imbibé d'eau distillée stérile.
2.2. Conditions de culture
Les boîte de Pétri, contenant chacune 25 graines, sont placées à l'obscurité dans une étuve à 24±1°C pendant une semaine. De l'eau distillée stérile est ajoutée en cas de dessèchement du papier filtre. Dix répétitions sont prévues pour chaque traitement et par variété.
2.3. Observations
Les boîtes de Pétri sont examinées tous les deux jours pour suivre la germination des graines. Le nombre de graines ayant germé est noté et le pourcentage de germination est ainsi calculé. Une graine germée est celle qui parvient à émettre une radicule et les deux cotylédons. L'effet du NaCl est estimé par le pourcentage d'inhibition de la germination apprécié comme suit :
%IG = PGte - PGtr / PGte x 100 PGte: pourcentage de germination du lot témoin
PGtr : pourcentage de germination du lot traité par le sel
3. Effet du sel sur la croissance végétative des plantes
3.1. Conditions de culture
3.1.1. Le substrat
Les cultures de tomate sont conduites sur du sable calibré de 1 à 2 mm de grosseur. Il est lavé à l'eau , puis mis à tremper dans l'acide chlorhydrique technique pendant 24 heures. Il est ensuite rincé une dizaine de fois avec de l'eau distillée jusqu'à stabilisation du pH autour de 3.5, puis lavé trois fois avec la solution nutritive jusqu'à ce que le pH qui percole s'établisse aux alentours de 6. Ce sable est ensuite stérilisé deux fois à l'autoclave pendant une heure à 24 heures d'intervalle. Avant son utilisation, le substrat est ré-imbibé avec la solution nutritive d’arrosage (annexes).
3.1.2. Préparation des pépinières
Les graines sont stérilisées par trempage dans l'alcool à 90° pendant deux minutes. Elles sont ensuite séchées sur papier filtre puis semées en pépinière. Elles sont arrosées tous les deux jours avec la solution nutritive complète sans sel.
3.1.3. Traitement par le sel
Arrivées au stade « premières vraies feuilles », les plantules sont déterrées délicatement et replantées dans des pots de sable de 450 cm3. Les plantules sont arrosées tous les deux jours avec la solution nutritive enrichie avec différentes concentrations de NaCl :0 ; 20 ; 40 ; 60 ; 80 et 100 mM soit 0 ; 1.16 ; 2.33 ; 3.50 ;
4.73 et 5.84 g/l. Les pots sont percés à la base pour permettre à la solution d'arrosage de percoler en cas d'excès.
3.1.3. La chambre de culture
Les cultures en pépinière et en pots sont effectuées dans une chambre de culture climatisée à 24 ± 1°C sous une photopériode de 16 heures et une humidité relative de 65%.
3.2. Lecture des résultats
Après six semaines d’observation, la croissance des plantes est estimée par deux variables : la hauteur de l’épicotyle et le poids frais des parties aériennes. Quatre heures après le dernier arrosage, les plantes sont délicatement déterrées et excisées au niveau de la cicatrice des cotylédons. Elles sont ensuite pesées. Les tests sont réalisés sur des lots de 15 plantes pour chaque variété de tomate et par concentration saline.
4. Analyse statistique des données
Chaque test est répété au moins deux fois. Les analyses statistiques sont effectuées à l’aide du logiciel SPSS. Les moyennes sont comparées par analyse des variances au seuil de 5% suivie par le test de Newman-Keuls.
B. Résultats
1. Action du sel sur le pouvoir germinatif des graines
1.1. Effet sur la vitesse de germination
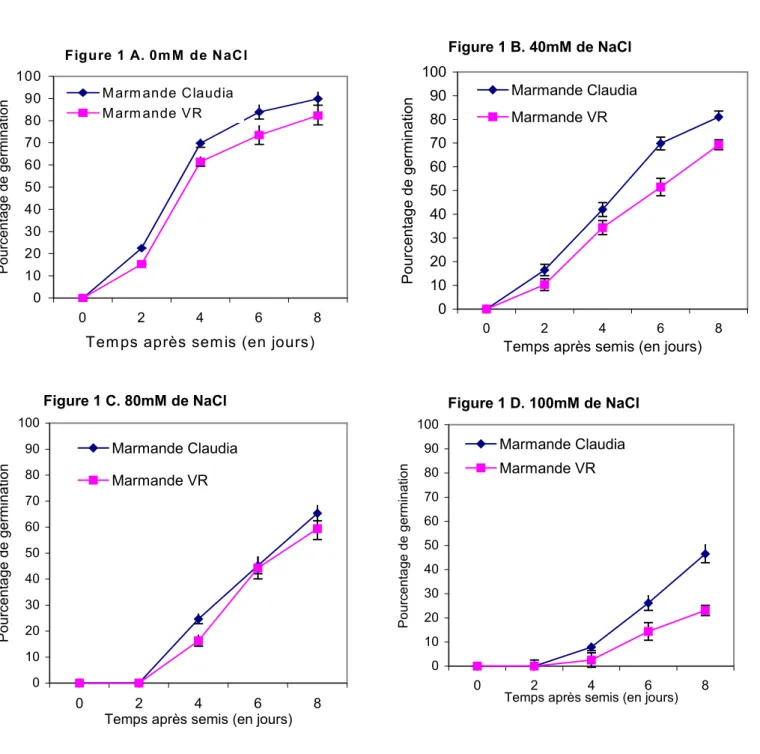
Les pourcentages de germination des variétés Marmande Claudia et Marmande VR ont été relevés tous les deux jours. Leur évolution en fonction du temps est illustrée sur la figure 1.
En absence de sel (figure 1A), les deux courbes obtenues, en forme de S, sont parallèles et présentent une zone linéaire située entre 2 et 4 jours au cours de
Figure 1. Effet de la salinité sur la vitesse de germination des graines de deux variétés de tomate.
Figure 1 C. 80mM de NaCl 0 10 20 30 40 50 60 70 80 90 100 0 2 4 6 8
Temps après semis (en jours)
Pourcentage de germination Marmande Claudia Marmande VR Figure 1 D. 100mM de NaCl 0 10 20 30 40 50 60 70 80 90 100 0 2 4 6 8
Temps après semis (en jours)
Pourcentage de germination Marmande Claudia Marmande VR Figure 1 A. 0m M de NaCl 0 10 20 30 40 50 60 70 80 90 100 0 2 4 6 8
Tem ps après semis (en jours)
Pourcentage de germination
Marm ande Claudia Marm ande VR Figure 1 B. 40mM de NaCl 0 10 20 30 40 50 60 70 80 90 100 0 2 4 6 8
Temps après semis (en jours)
Pourcentage de germination
Marmande Claudia Marmande VR
laquelle la vitesse de germination est maximale. Entre 4 et 8 jours, la pente des courbes diminue et les valeurs des % de germination tendent à se stabiliser.
En présence de sel, la vitesse de germination diminue avec l’augmentation de la salinité comme le montrent les pentes des courbes des figures 1B ; 1C et 1D. De même, un retard de la germination est noté. L’apparition de la radicule a lieu 48 heures après le semis chez les témoins ou chez les graines soumises aux concentrations modérées de sel. Cependant, elle ne se manifeste qu’entre 2 et 4 jours aux concentrations supérieures ou égales à 80mM de NaCl.
1.2. Effet sur le taux de germination
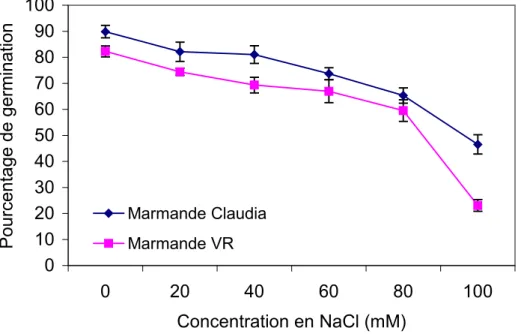
L’analyse des résultats consignés sur la figure 2 montre que la salinité affecte le taux de germination des deux variétés Marmande Claudia et Marmande VR. Les pourcentages de germination diminuent en fonction des concentrations croissantes de NaCl. Pour chacune des deux variétés, on note des différences significatives entre les différents traitements salins.
Néanmoins, pour un même traitement, les % de germination des deux variétés diffèrent statistiquement. En effet, après 8 jours de semis, les % de germination en absence de sel sont de 89.80% et 82.27% respectivement pour Claudia et VR. Cette différence est maintenue en présence de sel mais s’amplifie aux fortes concentrations. A 100mM de NaCl, le pourcentage de germination est de 46.53 % pour Marmande Claudia alors qu’il ne dépasse guère 23.5% pour Marmande VR. Cette dernière semble plus affectée par les fortes concentrations de sel par rapport à Marmande Claudia pour l’expression du pouvoir germinatif.
L’effet inhibiteur du sel se manifeste sur le pouvoir germinatif des variétés Claudia et VR dès la concentration de 20mM de NaCl (figure 3). Aux concentrations faibles et modérées, l’effet du sel s’exprime avec la même importance ; les pourcentages d’inhibition de la germination par rapport aux témoins sans sel ne montrent pas de différences significatives entre les deux variétés après 8 jours de semis. Aux fortes concentrations (100 mM de NaCl), les pourcentages d’inhibition de la germination s’écartent de manière hautement significative et sont de 48.21 % pour Marmande Claudia et 71.96 % pour Marmande VR.
Figure 2. Effet de la salinité sur le taux de germination de deux variétés de tomate après 8 jours de semis
0 10 20 30 40 50 60 70 80 90 100 0 20 40 60 80 100 Concentration en NaCl (mM) Pourcentage de germination Marmande Claudia Marmande VR
Figure 3. Pourcentage d'ihnibition de la germination des graines de deux variétés de tomate en fonction de la salinité
0 10 20 30 40 50 60 70 80 20 40 60 80 100 Concentration en NaCl (mM) Pourcentage d'inhibition de la germination Marmande Claudia Marmande VR
2. Action du sel sur la croissance végétative des plantes
Cette étude a été menée pour évaluer l’influence de la salinité sur la croissance végétative de la plante entière. Les résultats des mesures de la taille et du poids frais ont été analysés.
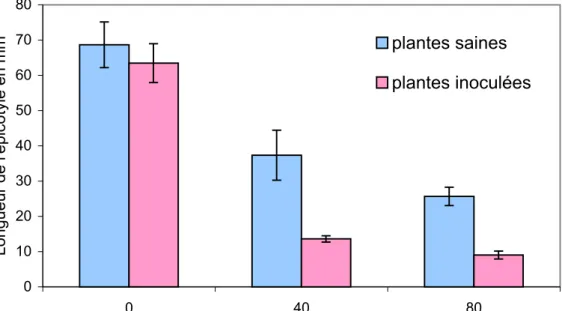
2.1. Effet sur la taille de l’axe aérien
Après six semaines d’observation, les deux variétés de tomate soumises au traitement par le sel montrent une réduction régulière et significative de la taille de l’épicotyle sous l’effet des concentrations croissantes de NaCl de la solution d’arrosage (figure 4). Pour chacune des deux variétés, on note des différences significatives entre les différents traitements par le sel à l’exception des traitements de 60mM et 80mM dont les effets sont similaires.
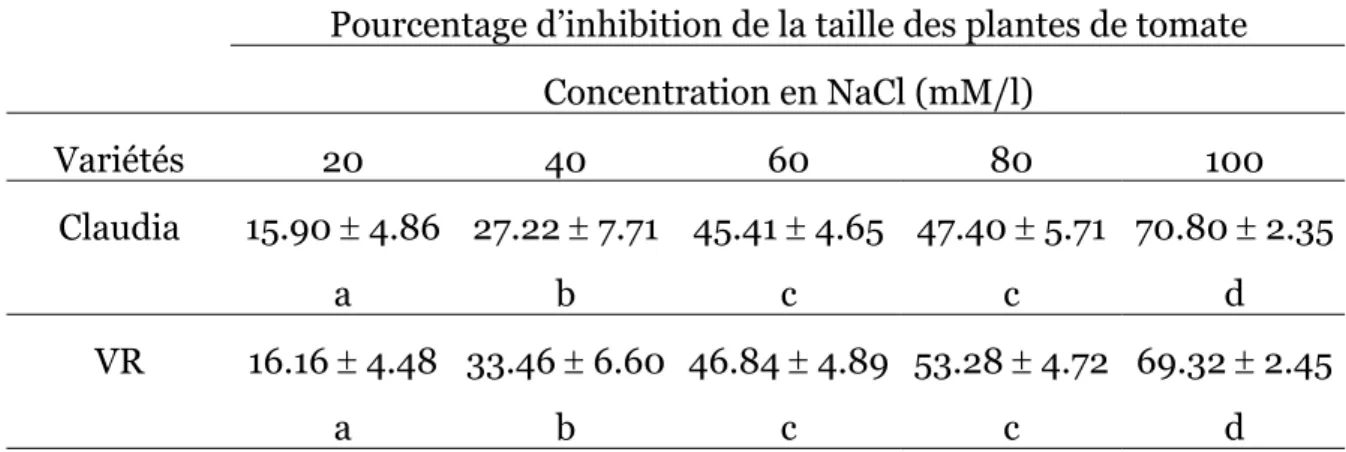
Les réductions de la croissance par rapport aux témoins sans sel sont reportées sur le tableau 1.
Tableau 1. Inhibition de la taille des tomates après traitement par NaCl.
Pourcentage d’inhibition de la taille des plantes de tomate Concentration en NaCl (mM/l) Variétés 20 40 60 80 100 Claudia 15.90 ± 4.86 a 27.22 ± 7.71 b 45.41 ± 4.65 c 47.40 ± 5.71 c 70.80 ± 2.35 d VR 16.16 ± 4.48 a 33.46 ± 6.60 b 46.84 ± 4.89 c 53.28 ± 4.72 c 69.32 ± 2.45 d Deux résultats lus sur une même ligne diffèrent statistiquement au seuil de 5% s’ils ne sont affectés d’aucune lettre en commun.
Les résultats montrent une certaine analogie dans les réponses des deux variétés. Pour une même salinité, les % de réduction de la croissance sont statistiquement similaires. L’inhibition de la croissance, faible aux premières concentrations de l’expérience, atteint à 60mM 45.41% pour Claudia et 46.84% pour VR. Elle
s’accentue avec l’augmentation de la salinité pour atteindre respectivement à 100mM de NaCl 70.8% et 69.32% pour Claudia et VR. L’analyse de la variance à deux critères ne montre pas de différence significative entre les deux variétés.
2.2. Effet sur la biomasse des parties aériennes
Les plantes ayant servi à l’étude de l’élongation des tiges sont excisées au niveau des cicatrices des feuilles cotylédonnaires puis pesées. Les résultats sont consignés sur la figure 5.
L’effet de la salinité s’est manifesté sur les deux variétés par une réduction significative du poids frais des tiges et feuilles corrélée avec l’augmentation de la salinité des eaux d’arrosage. Des différences significatives existent entre les traitements salins. L’effet de la salinité se manifeste selon le même schéma que pour l’effet sur la taille (tableau 2.). Les pourcentages d’inhibition de la biomasse des parties aériennes à 60 mM sont de 50.21 % et 51.59 % respectivement pour Claudia et VR, et de 67.27 % et 72.68 % à 100 mM. La différence notée entre les deux variétés n’est pas significative pour ce paramètre de la croissance.
Tableau 2. Inhibition du poids frais des parties aériennes des tomates après traitement par NaCl
Pourcentage d’inhibition du poids frais des parties aériennes Concentration en NaCl (mM/l) Variétés 20 40 60 80 100 Claudia 18.17 ± 9.67 a 35.06 ± 9.56 b 50.21 ± 6.61 c 53.12 ± 6.88 c 67.27 ± 5.25 d VR 24.51±10.60 a 24.51±10.60 b 51.59 ± 7.74 c 53.66 ± 5.70 c 72.68 ± 4.64 d Deux résultats lus sur une même ligne diffèrent statistiquement au seuil de 5% s’ils ne sont affectés d’aucune lettre en commun.
Figure 4. Effet du sel sur la hauteur de l'épicotyle des plantes de tomate après six semaines
0 5 10 15 20 25 30 35 40 0 20 40 60 80 100
Concentration des solutions d'arrosage en NaCl (mM)
Longueur de l'épicotyle (en cm)
Marmande VR Marmande Claudia
Figure 5. Effet du sel sur le poids frais des parties aériennes des plantes de tomate après six semaines
0 1 2 3 4 5 6 7 0 20 40 60 80 100
concentration des solutions d'arrosage en NaCl (mM)
Poids frais de l'axe aérien en g
Marmande Claudia Marmande VR
C. Discussion et conclusion
L’influence de la salinité sur le pouvoir germinatif des deux génotypes de tomate s’est manifestée par une réduction du taux et de la vitesse de germination par rapport aux témoins, réduction d’autant plus importante que la concentration en sel est élevée. L’analyse des variances a montré des différences significatives entre les deux variétés dans leur pourcentage de germination aux fortes concentrations de sel. La variété Marmande Claudia semble mieux tolérer les concentrations élevées de NaCl lors de la germination que Marmande VR. Nos résultats confirment les recherches de Berstein, (1975) et Bozouk, (1981) qui ont montré que chez la tomate cultivée (Lycopersicon esculentum), la germination est inhibée par les conditions salines.
La tolérance au sel au cours de la germination est une réponse directe de l’embryon à ses conditions nutritionnelles. Elle est directement liée à une sélectivité efficace du plasmalemme à l’égard de l’ion sodium. Cette sélection au stade embryonnaire est associée à une accumulation de calcium par la graine lors de la phase de maturation (Guerrier, 1983).
L’étude de l’effet du NaCl sur la germination de semences est insuffisante pour estimer la tolérance de la tomate au stress salin. En effet, la résistance au stress salin peut apparaître au stade germination et changer ou non pendant les phases suivantes de la croissance.
Le sel a eu un effet négatif sur la taille et le poids frais des tiges et feuilles des deux variétés cultivées dans nos conditions expérimentales. Les concentrations modérées de NaCl (60 mM) réduisent les performances de la croissance d’environ 50 % par rapport au témoin sans sel. Ce résultat correspond aux observations de Babas (1985), sur les variétés Vémone et Carmello, et de Amor (1991) qui a rapporté une baisse de la croissance de trois variétés de tomate, collectées dans les régions pré-sahariennes marocaines, en fonction de la salinité du milieu. Cet auteur a montré en outre, des différences nettes dans la réponse des différentes variétés suivant leur niveau de tolérance. D’après nos données, il n’y a pas de différence significative entre les deux génotypes pour les deux paramètres de la croissance évalués. La réponse au stress salin de la variété Marmande VR s’est
alignée avec celle de Marmande Claudia durant la phase végétative de la croissance contrairement au comportement différentiel observé lors de la germination.
Slama (1991) a évalué la tolérance au sel à partir du ralentissement de la vitesse de croissance des plantes à la concentration de 50mM de NaCl. Sous des concentrations voisines, la croissance des deux variétés de tomate a été réduite d’environ 50%. Ces résultats reflètent donc le niveau moyen de la tolérance au sel de ces deux génotypes. Ces derniers offrent ainsi un matériel de choix pour l’étude des effets combinés de la salinité et de l’infection avec Verticillium, principal agent des trachéomycoses au Maroc.
II. Influence de la salinité du milieu sur le développement in
vitro d’un isolat de Verticillium albo-atrum d’origine tomate
Introduction
Le développement des champignons phytopathogènes du sol dépend du comportement de la population des plantes hôtes et des caractéristiques de l’environnement. Les fluctuations de l’une ou l’autre de ces caractéristiques peuvent agir sur le développement et les activités biologiques de ces agents pathogènes et sur la relation hôte-parasite.
Verticillium est un champignon tellurique qui se conserve dans le sol sous forme
de microsclérotes. Ces formes de résistance peuvent rester à l’état dormant pendant plusieurs années en l’ absence de toute culture sensible. En présence de la plante hôte, les microsclérotes peuvent germer et attaquer les plantes par les racines. Le champignon prolifère à l’intérieur des vaisseaux du xylème et provoque par la suite des altérations foliaires et le flétrissement de la plante. Ce champignon vasculaire est ainsi soumis aux variations physico-chimiques du sol avant l’infection et à la variation de la composition chimique de la sève qui en découle une fois installé dans les vaisseaux du xylème.
Plusieurs travaux ont rapporté que l’augmentation de la salinité du milieu favorise la croissance et la conservation de Verticillium dahliae et de Fusarium oxysporum
fsp lycopersici , principaux agents des trachéomycoses au Maroc (Besri, 1977 ;
potentiels osmotiques extrêmement bas, alors que la salinité excessive peut être toxique pour d’autres champignons du sol. De même, la réduction du potentiel osmotique à des valeurs comprises entre –2 et –20 bars stimule la croissance mycélienne et la sporulation de Verticillium dahliae, mais la production des organes de résistance est moins sensible à la baisse du potentiel osmotique (Ioannou et al., 1977 ). Une stimulation de la formation des organes de conservation chez Fusarium oxysporum fsp lycopersici par la baisse du potentiel osmotique correspond aux observations de Besri (1977 et 1981 ) à condition que la disponibilité en eau du sol soit satisfaisante.
Ducan & Himelik (1986) ont montré que lorsqu’on cultive artificiellement
Verticillium dahliae sous un potentiel de pression négative de –0.039 MPa, la
production de conidies augmente de 800% par rapport au témoin non traité. L’influence de la salinité sur la germination des conidies de quelques isolats de
Verticillium lecanii a été étudiée par Chandler et al. (1994). Selon ces auteurs, le
pouvoir germinatif diminue avec la baisse du potentiel osmotique. Un seul isolat de V. lecanii a été sélectionné pour sa rapide germination après 12 heures à –2.8 MPa.
L’effet de la salinité a été étudié sur d’autres champignons pathogènes. C’est le cas des Phytophthora. Spp. Benyahya, (1998) a montré que l’augmentation de la salinité du milieu favorise la croissance mycélienne in vitro de Phytophthora
citrophthora et P. parasitica , agents de la pourriture racinaire des agrumes, avec
un optimum situé entre –1.44 et –3.11 bars. Au delà de cet intervalle, les activités biologiques régressent et la production de sporanges est complètement inhibée. D’autres espèces du même genre tolèrent des valeurs du potentiel osmotique beaucoup plus basses, comme Phytophthora cinnamomi qui montre une croissance optimale à –15 bars, alors que pour P. megasperma, la croissance diminue avec la baisse du potentiel osmotique (Sommers et al., 1970).
Les travaux de Swiecki & Mac Donald (1991) rapportent que la formation des sporanges de Phytophthora parasitica est plus importante en présence de concentrations modérées de NaCl et de CaCl2 par rapport aux témoins sans sel.
Cependant le nombre de zoospores produites et leur mobilité sont nettement plus faibles que ceux des témoins.
Parallèlement à ces travaux concernant l’action directe du sel sur le développement de ces champignons phytopathogènes, des études ont montré l’impact de la salinité sur la sensibilité des plantes aux maladies d’origine fongique. Dans ce sens, plusieurs auteurs rapportent une augmentation de la sensibilité des tomates à
Fusarium oxysporum f. sp. Lycopersici (Davet, 1966, Standaert, 1978) et à Verticillium dahliae (Besri, 1991 ; Besri & Afailal, 1993).
L’interprétation de l’effet de la salinité sur la plus grande sensibilité de la tomate à ces maladies est à rechercher, comme nous l’avons déjà précisé, au niveau de l’interaction de la plante et du pathogène avec l’environnement. La baisse de la croissance des plantes de tomate suite au stress salin décrite au paragraphe précédent représente un des aspects physiologiques de cette interaction. Il paraît donc nécessaire d’examiner dès lors l’impact de la salinité sur le développement de l’agent pathogène. A cette fin, nous nous proposons d’étudier in vitro l’effet de la salinité sur le développement de l’isolat P80 de Verticillium connu pour son fort pouvoir pathogène vis-à-vis des variétés Marmande Claudia et VR. Cette étude in
vitro contribuera à mieux comprendre le comportement du champignon dans les
sols riches en sel et à l’intérieur des vaisseaux conducteurs des plantes hôtes soumises au stress salin partant du fait que la composition chimique de la sève est le reflet de l’interaction physiologique de la plante avec son milieu nutritif .
A. Matériel et méthodes
1. La souche fongique
Lahlou (1983) a isolé Verticillium d’une tige de tomate, sévèrement atteinte de verticilliose, de la région de Dar Bouazza. L’isolat initial a été cloné et identifié comme appartenant à l’espèce Verticillium albo-atrum R & B, forme à microsclérotes. Le clone P3 , de phénotype sauvage, s’est montré très agressif sur la variété Marmande sensible.
Au cours de nos expériences préliminaires, des plantes de tomate, variété Marmande VR, ont été artificiellement infectées avec l’isolat P3 et régulièrement
arrosées avec une solution enrichie en NaCl. En fin d’expérience, le champignon a été ré-isolé d’une plante montrant des symptômes caractéristiques de rabougrissement. Cet isolat fut nommé P80 et a été utilisé dans tous nos essais. Le champignon est cultivé en boîte de Pétri sur milieu PDA ou milieu Malt gélosé, à l’obscurité et à 24± 1° C.
Le clone P80 est caractérisé par un mycélium blanchâtre d’aspect cotonneux et une sclérogénèse abondante apparaissant après 6 à 7 jours de culture. Il est très agressif sur la variété Marmande Claudia sensible mais aussi sur la variété Marmande VR possédant le gène Ve de la résistance. Il a été identifié donc comme appartenant à la race 2 de Verticillium.
2. Ensemencement
Les cultures liquides sont réalisées dans des fioles de 250 ml à raison de 100 ml par fiole. Elles sont ensemencées avec 1 ml d’une suspension de spores à 10 6 spores /ml fraîchement préparée à partir d’une culture de Verticillium âgée de 12 jours.
Les cultures solides, conduites sur boites de Pétri, sont ensemencées avec des rondelles de 4 mm de diamètre préalablement découpées à l’emporte pièce dans la zone de croissance active d’une culture âgée de deux semaines et présentant l’aspect caractéristique du type sauvage. Les thalles montrant des secteurs hyalins sont systématiquement écartés. Les boutures sont déposées au centre de la boîte et à l’envers pour que le champignon soit en contact direct avec le milieu de culture. Les boîtes sont fermées avec un ruban de parafilm pour éviter tout changement dans la pression osmotique du milieu par une éventuelle perte d’eau. Toutes les cultures sont mises à incuber à l’obscurité et à 24 ± 1°C.
3. Traitement par le sel
Les cultures fongiques sont conduites sur milieu à base d’extrait de malt liquide ou gélosé (2%) enrichi de chlorure de sodium à différentes concentrations 0 ; 5 ; 10 ; 15 ; 20 ; 30 ; 40 ; 50 ; 60 ; 80 et 100g/l selon le type d’expérience. Dix répétitions sont prévues pour chaque traitement salin.
4. Estimation de l’effet du sel sur la croissance de Verticillium 4.1. Croissance diamétrale
Les cultures sont examinées régulièrement, tous les jours. Les mesures des moyennes des deux diamètres perpendiculaires de chaque colonie sont faites tous les 5 jours. Le test a duré 20 jours.
4.2. Croissance pondérale
Bien que la croissance diamétrale soit la méthode la plus utilisée pour estimer l’effet d’un facteur de l’environnement sur un champignon, elle reste toutefois insuffisante car elle ne tient pas compte de la densité des filaments mycéliens ni du taux de la croissance spécifique. Pour palier à cette lacune, le poids sec du mycélium a été examiné comme un autre paramètre de la croissance pour estimer l’effet de la salinité sur le développement de Verticillium.
Après trois semaines d’incubation, les cultures liquides de Verticillium sont filtrées sur mousseline. Le mycélium recueilli est lavé deux fois à l’eau distillée, puis essoré entre plusieurs couches de papier filtre avant d’être séché à 80°C pendant 24 heures. Le poids sec du mycélium (mg) est ensuite déterminé.
5. Effet sur l’intensité de la conidiogénèse
Sur les boîtes ayant déjà servi pour l’étude de la croissance des colonies, dix rondelles de 5 mm de diamètre sont découpées à l’emporte pièce à 2mm environ du front de la croissance de chaque thalle. Elles sont ensuite placées dans un tube à vis contenant 10 ml d’eau stérile. Les tubes sont agités au vortex pendant 20 secondes, ce qui permet l’éclatement des sphérules portées par les conidiophores et la libération des conidies. Le contenu de chaque tube est ensuite filtré sur mousseline afin d’éliminer les fragments mycéliens et les morceaux de gélose. Le comptage du nombre total des conidies libérées par les dix rondelles est effectué avec un hématimètre ( cellule de Malassez) à raison de 5 comptages par suspension et de 10 boîtes par traitement salin. Les résultats sont exprimés en nombres de conidies par unité de surface (cm2).
6. Effet sur le pouvoir germinatif des conidies
Pour chaque traitement salin, une goutte contenant 100 conidies de Verticillium, issue d’une suspension de spores de l’isolat P80 fraîchement préparée, est étalée sur la surface de milieu gélosé à base d’extrait de malt enrichi en NaCl : 0 ; 4 ; 8 ; 12 ; 16 et 20 g/l. Dix boîtes sont prévues pour chaque concentration saline. Les étalements, incubés à l’obscurité et à 24°C, sont observés 24, 48 et 72 heures après ensemencement. Les pourcentages de germination des conidies sont alors déterminés.
7. Effet sur les organes de résistance
Cette étude a été faite en parallèle avec celle de la croissance diamétrale. L’effet de l’apport de NaCl dans le milieu de culture sur les microsclérotes est apprécié d’une part, par le délai d’apparition des organes de conservation et d’autre part, par l’estimation de l’étendue de la zone pigmentée visible sur le revers des boîtes de Pétri, la coloration noire des cultures de Verticillium étant due à la mélanisation des parois des microsclérotes. Tous les résultats sont comparés par analyse de la variance selon le test de Newman-Keuls.
B. Résultats
1. Effet de l’apport de NaCl sur la croissance de Verticillium
1.1. Croissance diamétrale
La croissance diamétrale des colonies de Verticillium soumis à différentes concentrations de sel a été notée tous les cinq jours durant 20 jours. Les résultats sont reportés sur la figure 6. Dès les cinq premiers jours qui suivent la mise en culture, l’effet de la salinité s’est manifesté par une stimulation de la croissance diamétrale du champignon pour les concentrations comprises entre 0 et 10 g/l de NaCl. Au delà de cet intervalle, la croissance diminue en corrélation avec l’augmentation de la salinité du milieu.
Après 20 jours de culture, l’intervalle des concentrations stimulatrices de la croissance s’étend à 20 g/l de NaCl. Au delà de cette concentration et jusqu’à 60
g/l , l’effet du sel s’est traduit par une réduction linéaire de la croissance mais qui reste toutefois supérieure et de manière significative à celle des témoins sans sel. Seules les très fortes concentrations inhibent la croissance mycélienne de
Verticillium. Cette inhibition est de l’ordre de 51.7% et 78.5% respectivement à 80
et 100 g/l de NaCl par rapport aux témoins sans sel.
L’apport de sel dans le milieu de culture agit également sur la vitesse de la croissance diamétrale des colonies (figure 7) par rapport aux témoins sans sel comme le montrent les pentes des différentes courbes de croissance. Durant la période d’incubation, la vitesse moyenne est de 1.27 mm /jour en absence de sel alors qu’elle atteint 2.05 mm /jour en présence de 20g/l de NaCl.
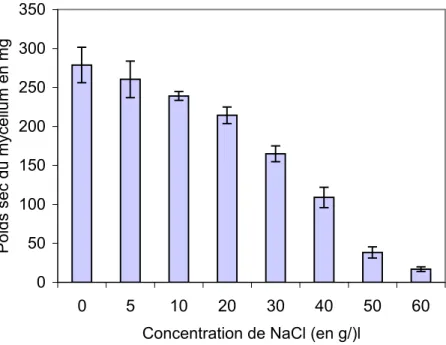
1.2. Croissance pondérale
Le poids sec du mycélium est évalué après un mois de culture en milieu liquide enrichi en NaCl. Les résultats reportés sur la figure 8 montrent que le poids sec du mycélium n’est pas modifié pour les concentrations de NaCl comprises entre 0 et 10 g/l. A partir de 10 g/l, on remarque une réduction régulière de la croissance pondérale du mycélium en fonction des salinités croissantes du milieu de culture. L’inhibition du poids sec est de 23.1% ; 60.9% et 93.9% respectivement à 20; 40 et 60 g/l de NaCl.
2. Effet de la salinité sur la conidiogénèse et le pouvoir germinatif des conidies
2.1. Intensité de la sporulation
Les résultats de l’estimation de la sporulation de Verticillium en fonction de la salinité du milieu sont résumés sur la figure 9. Il apparaît clairement que l’effet du sel se traduit par une stimulation de l’intensité de la conidiogénèse pour les
Figure 6. Effet de la salinité sur la croissance diamétrale de l'isolat P80 de Verticillium
0 5 10 15 20 25 30 35 40 45 50 0 5 10 20 30 40 50 60 80 100
Concentration en sel (en g/l de NaCl)
Moyenne des deux diamètres des
colonies (en mm)
après 5 jours après 10 jours après 15 jours après 20 jours
Figure 7. Vitesse de la croissance des colonies de l'isolat P80 de Verticillium en fonction de la salinité
du milieu 0 5 10 15 20 25 30 35 40 45 50 0 5 10 15 20
Temps d'incubation (en jours)
Diamètre moyen des colonies (en
mm)
Figure 8. Effet de la salinité sur le poids sec du mycélium de l'isolat P80 de Verticillium
après un mois de culture
0 50 100 150 200 250 300 350 0 5 10 20 30 40 50 60
Concentration de NaCl (en g/)l
Poids sec du mycélium en mg
Figure 9. Effet de la salinité sur l'intensité de la conidiogénèse de l'isolat P80 de Verticillium
après un mois de culture
0 20 40 60 80 100 120 140 160 180 200 0 5 10 15 20 40 60
Concentration de NaCl (en g/)l
Nombre de spores x 10
5 /cm
concentrations de NaCl allant de 0 à 15 g/l. En effet, à la concentration optimale, (15g/l), on enregistre une augmentation du nombre de conidies produites de 130 % par rapport au témoin sans sel. Pour les autres concentrations de l’expérience supérieures à 15 g/l, l’intensité de la sporulation diminue mais reste statistiquement supérieure à celle du témoin sans sel jusqu’à 40g/l.
2.2. Pouvoir germinatif des conidies
La germination in vitro des conidies de Verticillium a été suivie pendant 72h. Les résultats montrent que les conidies ont germé à toutes les salinités de l’expérience (figure 10). A 4 g/l de NaCl, le pourcentage de germination des conidies est semblable à celui du témoin non traité et atteint 71.8 %. La germination subit une hausse significative sous les concentrations salines comprises entre 8 et 16g/l de NaCl (Planche 1). En revanche, l’apport dans le milieu de 20g/l de sel réduit le pouvoir germinatif des conidies traitées. Le pourcentage de germination, bien que statistiquement plus faible que celui des conidies témoins sans sel, est encore appréciable ; il est de 67.6%.
3. Effet de la salinité sur la sclérogénèse
L’estimation de l’abondance des microsclérotes par la mesure du diamètre de la zone pigmentée où se localisent les microsclérotes n’est certes pas très précise mais permet de faire des comparaisons entre les effets des différents traitements salins sur la formation des organes de résistance.
Des résultats présentés dans la figure 11, il ressort que l’apport de faibles quantités de sel (0 à 5 g/l) n’altère pas l’importance de la formation des microsclérotes. Leur abondance ne diffère pas de celle des témoins sans sel. Cependant l’augmentation des concentrations de NaCl dans le milieu de culture a entraîné une diminution nette de la sclérogénèse. Cette réduction est d’autant plus marquée que la concentration saline est élevée. L’effet de la présence de concentrations de sel supérieures à 5 g/l touche aussi bien l’étendue de la zone pigmentée que l’intensité de la coloration qui devient de plus en plus claire.
Figure 10. Effet de la salinité sur le pouvoir germinatif des conidies de l'isolat P80 de
Verticillium après 72 heures
40 50 60 70 80 90 100 0 4 8 12 16 20
Concentration en NaCl (en g/)l
Pourcentage de germination des conidies
Figure 11. Effet de la salinité sur l'intensité de la sclérogénèse de l'isolat P80 de Verticillium après
un mois de culture 0 0,5 1 1,5 2 2,5 3 3,5 4 4,5 0 5 10 15 20 40 60
Concentration en NaCl (en g/l)
Diamètre moyen de la zone
Planche 1. Effet de la salinité du milieu sur la germination des conidies de l’isolat P80 de Verticillium albo-atrum après 4 jours d’incubation.