Technological Features of Selected Kivuguto Strains during Milk Fermentation
Texte intégral
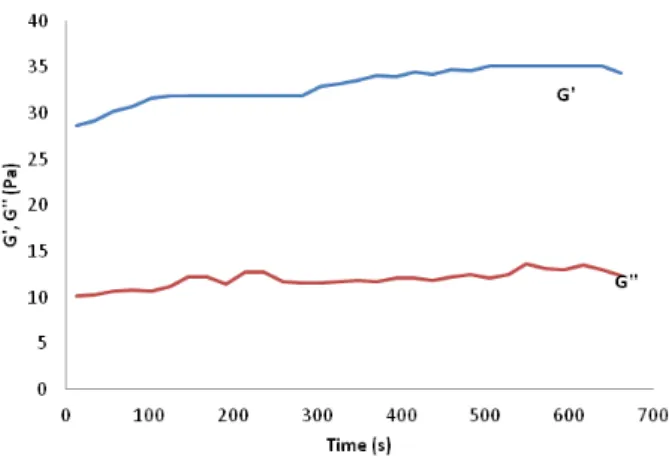
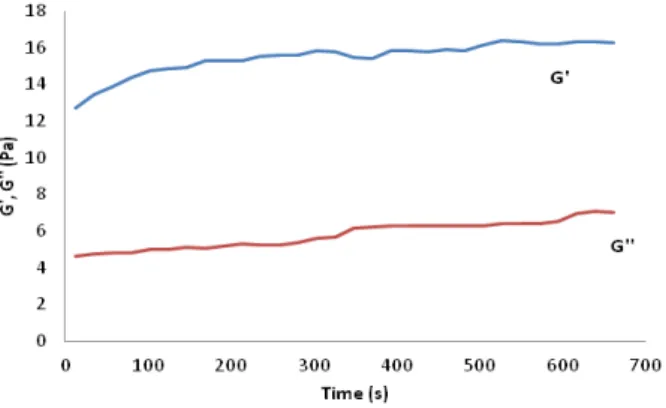
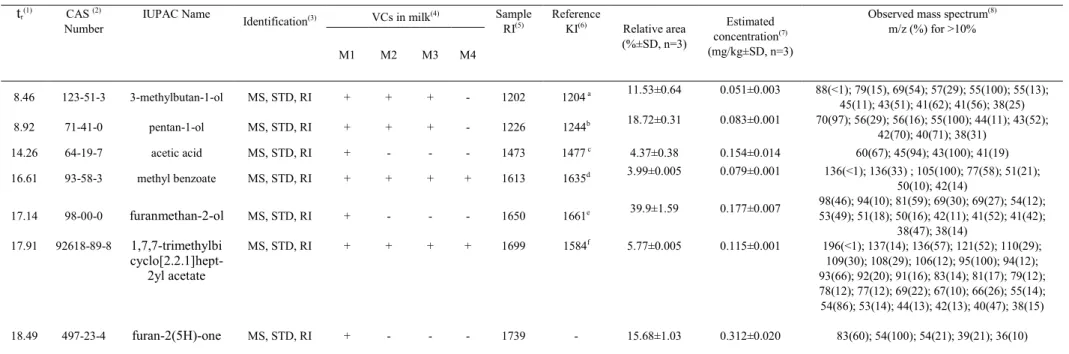
Figure

Documents relatifs
When fermented milk (inoculated with or without LAB at 0 or 24 h of fermentation) was cooked at 50°C for Ayib-making, DT104 survived up to 60 min of cooking with survival varied
Inoculations of raw camel milk with selected LAB strains (E. duransR03, E.faecium NWL and L. plantarum BJ6 and their combination as well as the control - fermentation without
The quantity of faeces lead in groups treated by fermented milk product containing or not lead nitrate was high relatively to Control and Water Pb groups.. Such observation
They were divided into four treatment groups: (1) cavies not receiving lead and used as control group, (2) treated group with 2 mL of solution containing Lead Nitrate (0.5 ppm)
Alternatively, one could also envisage that more frequent udder evacuation and milk removal would not only reduce accu- mulation of FIL, but would also avoid any deleterious effects
The desulfurization yields calculated from the total sulfur content of various types of diesel oil after 96 h incubation in the presence of resting cells from the selected strains
Fabienne Feutry, Paloma Torre, Ines Arana, Susana Garcia, Nathalie Desmasures, et al.. Lactococcus lactis strains from raw ewe’s milk samples from the PDO Ossau-Iraty cheese
Presence of milk phospholipids and milk lipids modified the immunomodulatory properties of infant formula in neonatal pigs.. 4th World Congress of Pediatric Gastroenterology,
