Modélisation de maladies cérébrovasculaires associées
aux variations génétiques de RNF213 par le génie
tissulaire et la culture cellulaire 3D
Thèse
Vincent Roy
Doctorat en médecine expérimentale
Philosophiæ doctor (Ph. D.)
Résumé
Le gène RNF213 a été identifié comme un facteur de risque associé au développement de maladies cérébrovasculaires (MCV) notamment, la maladie de Moyamoya (MMM) et les anévrismes intracrâniens (AIC). Malgré une incompréhension des fonctions biologiques exactes, la ring finger protein 213 (RNF213) serait impliquée dans la régulation de la prolifération cellulaire, de l’angiogenèse et de l’inflammation. Les travaux présentés dans cette thèse se concentrent sur le développement de modèles vasculaires in vitro afin de mieux caractériser le rôle de RNF213 dans MCV. L’hypothèse est que l’invalidation de la protéine RNF213 dans des cellules endothéliales (CE) cerébrales pourrait recréer certains phénotypes associés au développement de la MMM et à la formation d’AIC.
Des cellules endothéliales microvasculaires humaines de cerveau (hCMEC/D3) invalide en RNF213 (RNF213-/-) ont été initialement générées par la méthode Clustered Regularly Interspaced Short Palindromic
Repeats et la protéine associée Cas9 (CRISPR-Cas9) double nickase. La première partie des travaux porte sur le rôle que jouerait RNF213 dans l’homéostasie de la barrière hémato-encéphalique (BHE) et dans les étapes précoces de la pathogenèse associée à la MMM. Plus précisément, la perte des jonctions adhérentes provoquée par l’invalidation de RNF213 dans les hCMEC/D3 a été évaluée in vitro sur plusieurs paramètres, tels que la morphologie endothéliale, l’expression génique des protéines de jonctions, la localisation cellulaire, la perméabilité, l’infiltration immunitaire et le sécrétome des cytokines inflammatoires. Les résultats ont démontré que la déficience en RNF213 provoque une diminution de l’expression de la platelet endothelial cell adhesion molecule-1 (PECAM-1) qui affecte conséquemment la formation adéquate du complexe jonctionnel. De plus, une diminution de l’expression des gènes de la claudine-5, de la b-caténine et de la plakoglobine a été mesurée. La perte de RNF213 est également accompagnée d’un relargage de plusieurs cytokines pro-inflammatoires.
En deuxième lieu, les travaux de cette thèse ont également démontré que RNF213 joue un rôle prépondérant dans le processus angiogénique des hCMEC/D3. Ceci a été étudié sous plusieurs angles d’approches, tels que la prolifération et la migration cellulaire, la formation de micro-capillaires sur un support Matrigel® et dans un modèle tridimensionnel (3D) reconstruit par génie tissulaire, l’expression génique et le sécrétome angiogénique. Les résultats ont démontré une diminution du taux de division cellulaire et une augmentation de la migration. Les études in vitro ont démontré également, pour la première fois, une augmentation significative de la formation de micro-capillaires et de la sécrétion abondante de facteurs pro-angiogénique, tels que le vascular endothelial growth factor (VEGF). Plus précisément, les hCMEC/D3 déficientes en RNF213 forment un réseau plus vaste, dense et étendu de micro-capillaires sur un support de Matrigel®.
Lorsqu’ensemencées dans un modèle 3D plus complexe structurellement, les hCMEC/D3 forment un réseau pouvant s’apparenter au réseau de capillaires compensatoire retrouvé chez les patients MMM. Dans l’ensemble, l’invalidation du gène RNF213 dans un modèle in vitro 3D de cellules endothéliales cérébrales permet de reproduire certains phénotypes pathologiques de la MMM et devient donc ainsi le 1er modèle in vitro pour l’étude de cette maladie et des autres maladies associées à RNF213.
Finalement, nous avons mis au point un nouveau modèle de vaisseaux sanguins de petit calibre reconstruit par génie tissulaire (TEBV) pour son utilisation dans l’étude in vitro de maladies vasculaires et de MCV complexes. L’ensemencement de fibroblastes ou de cellules musculaires lisses (CML) directement sur un mandrin de polyéthylène téréphtalate glycol (PETG) prétraité aux rayons ultraviolets C (UV-C) a permis de former des feuillets circulaires, manipulables et superposables. Avec cette technique, nous avons généré des TEBV complets avec les trois principales couches, soit l’adventitia, la media et l’intima tunica, qui possèdent des propriétés histologiques et mécaniques similaires aux artères humaines natives. Ce modèle optimisé et uniformisé de TEBV permettra de modéliser des pathologies vasculaires complexes, telles que la MMM et les AIC. En effet, la génération de vaisseaux complets à partir de cellules pathologiques ou de cellules éditées génétiquement pourrait faciliter la caractérisation de la pathogenèse et aider au développement de médicaments.
Abstract
RNF213 has been associated as a susceptibility gene for the development cerebrovascular diseases (CVDs), in particular, moyamoya disease (MMD) and intracranial aneurysms (ICA). While the exact biological functions of RNF213 remain to be demonstrated, it is known to be involved in the regulation of cell proliferation, angiogenesis and inflammation. The work presented in this thesis focuses on the development of vascular models in vitro to better characterize the role of RNF213 in CVDs. The hypothesis is that the complete invalidation of the RNF213 protein in brain endothelial cells (EC) could recreate evident phenotypes associated with the development of MMD and the formation of ICA.
We have initially generated human brain microvascular endothelial cells (hCMEC/D3) deficient in RNF213 (RNF213-/-) using the robust CRISPR-Cas9 double nickase method. At first, our work described the role that
RNF213 would play in the homeostasis of the blood-brain barrier (BBB) maintenance and in the early stages of MMD pathogenesis. More specifically, the loss of adherent junctions caused by the invalidation of RNF213 in hCMEC/D3 was evaluated in vitro on several parameters, such as endothelial morphology, gene expression of junctional proteins, cellular localization, permeability, immune infiltration and the secretome of inflammatory cytokines. Our data demonstrated that RNF213 deficiency provokes a significant decrease in the platelet endothelial cell adhesion molecule-1 (PECAM-1) expression, which consequently affects the proper formation of the junctional complex. A decrease in the expression of the claudin-5, b-catenin and plakoglobin genes was also measured. In addition, RNF213 loss is accompanied with a release of several pro-inflammatory cytokines.
Thereafter, the present work also demonstrated that RNF213 plays a preponderant role in the angiogenic process of hCMEC/D3. Angiogenesis has also been characterized on several aspects, such as proliferation, migration, formation of micro-capillaries on a Matrigel®-based support and in a 3D model reconstructed by tissue engineering, gene expression and secretion of angiogenic factors. Our data demonstrates a decrease in cell division rate and an increase in cell migration. In vitro studies have also shown, for the first time, a significant increase in micro-capillary formation and abundant secretion of pro-angiogenic factors, such as the vascular endothelial growth factor (VEGF). More precisely, the hCMEC/D3 deficient in RNF213 forms a wider, denser and more extensive network of micro-capillaries on a Matrigel®-based support. When seeded in a more structurally complex 3D model, hCMEC/D3 form a network that can resemble to the compensatory capillary network found in MMM patients. Overall, the invalidation of the RNF213 gene in a 3D in vitro model of cerebral endothelial cells makes it possible to reproduce certain pathological phenotypes of MMM and
therefore becomes the 1st in vitro model for the study of this disease and other diseases associated with
RNF213.
Finally, we developed a new model of small-caliber blood vessels reconstructed by tissue engineering (TEBV) to be used to study vascular diseases and complex CVD in vitro. The direct seeding of fibroblasts or smooth muscle cells (CML) onto a polyethylene terephthalate glycol (PETG) mandrel that was pretreated with ultraviolet C (UV-C) radiation facilitate the formation of circular cell sheets, which could be manipulated and stacked in top of each other. Using this novel technique, we were able to successfully generate complete TEBVs with the three main arterial layers: the adventitia, the media and the intima tunica. Taken together, our TEBV model has histological and mechanical properties similar to native human arteries. Furthermore, this optimized and standardized 3D vascular construct will accelerate the scientific progress to modelized complex vascular pathologies, such as MMD and AIC. Indeed, the generation of complete vessels derived from pathological cells or genetically edited cells could facilitate the characterization of pathogenesis and help in the development of drugs.
Table des matières
Résumé ... ii
Abstract ... iv
Table des matières ... vi
Liste des tableaux ... xii
Liste des figures ... xiii
Liste des abréviations ... xv
Remerciements ... xviii
Avant-propos ... xx
Introduction ... 1
Problématique générale ... 1
Anatomie et physiologie cérébrovasculaire ... 3
Polygone de Willis ... 5
Anatomie ... 6
Fonction biologique ... 7
Autorégulation de la circulation et de la pression ... 7
Variations de la configuration ... 8
Physiologie et structure des artères cérébrales ... 9
Adventitia tunica ... 10
Media tunica ... 10
Intima tunica ... 11
Vasa vasorum intracrânien ... 11
Barrière hémato-encéphalique ... 12
Cellules de la BHE et de l’unité neurovasculaire ... 14
Cellules endothéliales ... 14
Péricytes et cellules murales ... 14
Membrane basale ... 15
Astrocytes ... 15
Microglies ... 16
Neurones ... 16
Cellules immunitaires ... 17
Complexes jonctionnels de la BHE ... 17
Occludine ... 18
Claudines ... 19
Protéines d’échafaudage ... 19
Jonctions adhérentes ... 20
Passage à travers la BHE ... 21
Les voies de transport paracellulaire ... 22
Les voies de transport transcellulaire ... 22
Transmigration immunitaire ... 23
Modélisation de la barrière hémato-encéphalique in vitro ... 24
Angiogenèse ... 25
Angiogenèse Physiologique ... 25
Angiogenèse du SNC et développement de la BHE ... 27
Régulateurs de l’angiogenèse ... 28
VEGF ... 28
Angiopoïétines et le récepteur Tie-2 ... 28
PDGF ... 29 FGF ... 29 Signalisation de NOTCH et de WNT ... 29 Intégrines et MMP ... 29 Protéines jonctionnelles ... 30 TGF-β ... 31 Cytokines pro-angiogéniques ... 31
Débalancement angiogénique et cible thérapeutique ... 31
Modélisation de l’angiogenèse in vitro ... 32
Maladies cérébrovasculaires ... 33
Anévrismes Intracrâniens ... 33
Épidémiologie et étiologie ... 34
Pathogenèse ... 35
Présentations cliniques et diagnostic ... 36
Traitements ... 37
Maladie de Moyamoya ... 38
Étiologie et épidémiologie ... 38
Pathogenèse ... 39
Présentations cliniques et diagnostic ... 40
Traitements ... 41
RNF213 et prédispositions génétiques ... 42
Ring finger protein 213 ... 42
RNF213 dans la Maladie de Moyamoya ... 43
Angiogenèse ... 44
Inflammation ... 44
Autres fonctions ... 45
RNF213 dans les anévrismes intracrâniens ... 45
Les autres variants de RNF213 et leur phénotype ... 45
Substituts vasculaires ... 47
Vaisseaux natifs ... 47
Vaisseaux synthétiques ... 47
Vaisseaux reconstruits par génie tissulaire ... 48
Le génie tissulaire et la culture 3D ... 48
Vaisseaux décellularisés ... 50
Polymères synthétiques et biodégradables ... 50
Biopolymères naturels ... 50
Auto-assemblage en génie tissulaire; un outil pour la modélisation 3D de MCV ... 51
Édition du Génome ... 53
Principes de bases ... 53
Nucléases à doigts de zinc ... 54
Nucléases effectrices de type activateur de transcription ... 54
Méganucléases ... 55
CRISPR-Cas9 ... 55
CRISPR: une ligne de défense contre les infections virales ... 55
Adaptation pour l’édition ciblée du génome ... 56
Modifications et applications de CRISPR-Cas9 ... 57
Problématique, hypothèse et objectifs de recherche ... 58
Chapitre 1 : La perte de fonction Le gène de susceptibilité à la maladie de Moyamoya RNF213 régule la fonction de la barrière endothéliale ... 60
1.1. Résumé ... 61
1.2. Abstract ... 63
1.3 Introduction ... 64
1.4 Materials and methods ... 65
1.4.1. Cell culture ... 65
1.4.2. Cell circularity index ... 66
1.4.3. Microarray-based gene expression profiling and bioinformatics analysis ... 66
1.4.4. Western blot analysis ... 66
1.4.5. Detection of apoptosis by flow cytometric ... 67
1.4.6. Transendothelial electrical resistance ... 67
1.4.8. Immune infiltration assay ... 68
1.4.9. Immunofluorescence ... 68
1.4.10. Human cytokine array ... 69
1.4.11. ELISA ... 69
1.4.14. Statistical analysis ... 69
1.5 Results ... 70
1.5.1. RNF213 is predominantly expressed in hCMEC/D3 and endothelial cells ... 70
1.5.2. CRISPR-Cas9-mediated knockout of RNF213 in hCMEC/D3 cell line ... 70
1.5.3. RNF213-mediated knockout altered cerebral endothelial cell morphology ... 73
1.5.4. RNF213 regulates cerebral endothelial cell functions and BBB integrity ... 73
1.5.5. RNF213 deficiency is associated with a downregulation of PECAM-1 expression and an abnormal intercellular junction formation ... 75
1.5.6. CRISPR-Cas9-mediated RNF213 knockout in hCMEC/D3 altered the BBB functions by increasing transmigration and accumulation of immune cells ... 76
1.5.7. Secretion of MMD-associated pro-inflammatory cytokines by brain endothelial RNF213-deficient cells ... 78
1.6 Discussion ... 80
1.7 Conclusion ... 83
1.8 Acknowledgements ... 83
1.9 Supplementary materials and methods ... 84
1.9.1. Cloning of mU6-pSpCas9-2A-Puro plasmid ... 84
1.9.2. Cloning sgRNAs into pSpCas9(BB) vector ... 84
1.9.3. T7 endonuclease I mismatch detection assay ... 84
1.9.4. Cloning of Lentiviral vector ... 85
1.9.5. Production of integrative lentiviral particle ... 85
1.9.6. Generation of hCMEC/D3-RNF213-/- and hCMEC/D3-Cas9n ... 86
1.9.7. Real-time reverse transcription quantitative PCR ... 86
1.9.8. Analysis of membranous PECAM-1 by flow cytometric ... 86
1.9.9. PBMC isolation ... 86
1.9.10. PBMC stimulation ... 87
1.9.11. Immune infiltration assay analysis ... 87
1.10 Supplementary Tables ... 88
1.11 Supplementary Figures ... 91
1.12 Références ... 102
Chapitre 2 : Le gène de susceptibilité à la maladie de Moyamoya RNF213 régule l’angiogenèse des cellules endothéliales de cerveau par la sécrétion de facteurs angiogéniques ... 109
2.1. Résumé ... 110
2.2. Abstract ... 112
2.3 Introduction ... 113
2.4.1. Cell culture and CRISPR-Cas9-mediated RNF213 invalidation ... 114
2.4.2. Cell proliferation assays ... 114
2.4.3. In vitro scratch assays ... 115
2.4.4. Tube formation assay ... 115
2.4.5. 3D capillaries network formation ... 116
2.4.6. Human angiogenesis proteome profiler ... 116
2.4.7. ELISA ... 116 2.4.8. Immunoblotting ... 116 2.4.9. Immunofluorescence ... 117 2.4.10. Microarray ... 117 2.4.11. Statistical analysis ... 118 2.5 Results ... 118
2.5.1. CRISPR-Cas9-mediated RNF213 knockout altered cerebral microvascular endothelial cell morphology 118 2.5.2. RNF213 regulates proliferation and cell-cycle of cerebral endothelial cells ... 119
2.5.3. CRISPR-Cas9-mediated RNF213 knockout drives cellular migration as well as the angiogenic activity of cerebral endothelial cells ... 121
2.5.4. RNF213 controls angiogenesis through secretion of pro-angiogenic factors ... 124
2.5.5. Tridimensional modeling of a cerebral microvasculature to mimic and evaluate the impact of RNF213 invalidation ... 124
2.6 Discussion ... 126
2.7 Acknowledgements ... 128
2.8 Supplementary figures ... 129
2.9 References ... 134
Chapitre 3 : L’ensemencement cellulaire sur des support 3D en polymères traités aux UV-C permet la production rentable de vaisseaux sanguins de petit calibre reconstruits par génie tissulaire ... 138
3.1 Résumé ... 139
3.2 Abstract ... 141
3.3 Introduction ... 142
3.4 Experimental Section ... 143
3.4.1 Ethical Statements, Cell Isolation, and Culture ... 143
3.4.2 Surface Treatment of the Substrate ... 143
3.4.3 Wettability of the Substrates ... 144
3.4.4 Fourier Transform Infrared Spectroscopy of Treated Substrates ... 144
3.4.5 Cellular Adhesion and Morphology ... 144
3.4.6 Monolayered Fibroblastic Tubular Conduit Production ... 144
3.4.7 Multilayered Tissue-Engineered Blood Vessel Production ... 146
3.4.8 Mechanical Testing ... 146
3.4.9 Histological Coloration ... 146
3.4.11 Cell Orientation Analysis ... 148
3.4.12 Statistical Analysis ... 148
3.5 Results ... 148
3.5.1 Effects of UV-C Treatment on Plastic Surfaces: Surface Oxidation, Wettability, and Cell Adhesion ... 148
3.5.2 Histological and Mechanical Property Comparisons Between the Tested Tissue-Engineered Fibroblast Conduits Production Methods ... 151
3.5.3 Properties of TEBV Using Stacked Tubular Constructs ... 151
3.5.3.1 Histological Analysis and Mechanical Properties of the Mandrel-Seeded Reconstructed Tissue-Engineered Blood Vessels ... 151
3.5.3.2 Homogeneous Cellular Distribution, and Cell-Driven Reorganization of Smooth Muscle Cells ... 155
3.6 Discussion ... 155
3.7 Conclusion ... 157
3.8 Acknowledgements ... 157
3.9 Supplementary Figures ... 158
3.10 References ... 165
Chapitre 4 : Discussion générale ... 168
4.1 La lignée hCMEC/D3 est idéale pour l’étude de MCV in vitro ... 168
4.2 Génération de hCMEC/D3 déficientes en RNF213par CRISPR-Cas9 ... 170
4.3 RNF213 et la perturbation de la BHE in vitro ... 172
4.3.1 Les protéines du complexe jonctionnel et l’étanchéité de la barrière ... 173
4.3.2 Altération de la transmigration immunitaire et des cytokines inflammatoires ... 175
4.4 RNF213 et l’angiogenèse dans la maladie de Moyamoya ... 178
4.4 Les TEBV: un modèle 3D d’étude de pathologies vasculaires ... 182
Conclusions générales et perspectives ... 186
Références ... 188
Annexe 1 ... 246
Annexe 2 ... 247
Liste des tableaux
Tableau 1.1 – Taille et diamètre externe moyens des artères principales du cercle de Willis ... 6
Tableau 1.2 – Classification des MMP et leur rôle dans l’angiogenèse ... 30
Tableau 1.3 – Diversité des variations génétique de RNF213 dans les MCV ... 46
Tableau 1.4 – Méthodes de reconstruction de vaisseaux par génie tissulaire ... 52
Tableau 1.5 – Comparaison des nucléases les plus utilisées pour l’édition du génome ... 57
Supplementary Table 2.1– Gene expression of endothelial cell markers ... 88
Supplementary Table 2.2 – Cytokines and chemokines genes expression data ... 89
Supplementary Table 2.3 – Primer sequences ... 90
Supplementary Table 2.4 – sgRNAs sequences and characteristics ... 90
Supplementary Table 2.5 – Oligo sequences for sgRNAs cloning ... 90
Supplementary Table 2.6 – Oligo primers for qPCR assays ... 90
Liste des figures
Figure 1.1 – Les principales artères et veines du système cérébrovasculaire ... 4
Figure 1.2 – Le polygone de Willis ... 5
Figure 1.3 – Exemples d’anomalies de la configuration du cercle de Willis ... 9
Figure 1.4 – Les structure majeures des artères intracrâniennes et extracrâniennes ... 10
Figure 1.5 – Les barrières du SNC ... 12
Figure 1.6 – Les CE du SNC et de la vasculature périphérique ... 13
Figure 1.7 – Composition moléculaire des jonctions serrées et jonctions adhérentes ... 18
Figure 1.8 – Les différentes voies de passage à travers la BHE ... 21
Figure 1.9 – Transmigration des leucocytes à travers la barrière hémato-encéphalique ... 24
Figure 1.9 – Représentation schématique de la formation de nouveaux vaisseaux. ... 26
Figure 1.10 – Types d'anévrismes cérébraux et hémorragie sous-arachnoïdienne ... 34
Figure 1.12 – Occlusion artérielle et capillaires compensatoires ... 38
Figure 1.13 – Domaines fonctionnels, variations génétiques et oligomérisation de RNF213 ... 42
Figure 1.14 – Exemples de tissus reconstruit par génie tissulaire avec la méthode d’auto-assemblage ... 49
Figure 1.15 – Schématisation des différentes techniques pour la reconstruction de TEBV par auto-assemblage ... 52
Figure 1.16 – Différentes voies de réparation et d’édition de l’ADN induites par les nucléases ... 53
Figure 1.17 – Diverses applications du système CRISPR-Cas9 ... 56
Figure 2.1 – RNF213 expression in hCMEC/D3 and various cell types. ... 71
Figure 2.2 – CRISPR-Cas9-mediated knockout of RNF213 in hCMEC/D3 cell line. ... 72
Figure 2.3 – RNF213 knockout alters endothelial cell morphology. ... 74
Figure 2.4 – Ingenuity Pathway Analysis generated interactome of genes related to BBB functions and diseases of interest. ... 75
Figure 2.5 – RNF213 deficiency is associated with a downregulation of PECAM-1 expression and an abnormal intercellular junction formation. ... 77
Figure 2.6 – RNF213 deficiency in hCMEC/D3 enhanced lymphocytes transmigration and secretion of pro-inflammatory cytokines and chemokines. ... 79
Supplementary Figure 2.1 – hCMEC/D3s monolayer organization. ... 91
Supplementary Figure 2.2 – Detection of apoptosis in hCMEC/D3s. ... 92
Supplementary Figure 2.3 – Validation of gene expression by RT-qPCR. ... 93
Supplementary Figure 2.4 – Phosphorylation level of β-catenin. ... 94
Supplementary Figure 2.4 – Reduction of PECAM-1 glycosylation in hCMEC/D3-RNF213-/-. ... 95
Supplementary Figure 2.6 – Cell surface expression of PECAM-1. ... 96
Supplementary Figure 2.7 – Flow cytometry characterization of PBMCs for the transmigration assay. ... 97
Supplementary Figure 2.8 – Accumulation of CD3+ lymphocytes on hCMEC/D3 monolayers. ... 98
Supplementary Figure 2.10 – Human cytokines proteome profiler. ... 100
Supplementary Figure 2.11 – Enhanced CCR2 protein expression in hCMEC/D3-RNF213-/-. ... 101
Figure 3.1 – RNF213 knockout in hCMEC/D3 ... 119
Figure 3.2 – RNF213 is associated with cell proliferation and cell-cycle regulation ... 120
Figure 3.3 – RNF213 deficiency promotes cell migration and tube-like formation ... 121
Figure 3.4 – RNF213 invalidation enhanced the secretion of pro-angiogenic factors ... 122
Figure 3.5 – RNF213 deficiency is associated with the formation of moyamoya-like vessels in vitro ... 125
Supplementary Figure 3.1 – Post-filtering and -normalization principal component analysis plots of microarray expression data generated with the Network Analyst platform. ... 129
Supplementary Figure 3.2 – Heat map of the significantly differentially expressed genes between WT and RNF213 -/-cells generated with the Network Analyst platform. ... 131
Supplementary Figure 3.3 – Total secreted angiogenic factors. ... 132
Supplementary Figure 3.4 – Microcapillaries formation in self-assembled constructs. ... 133
Figure 4.1 – Schematic of the techniques used to produce tissue-engineered tubular tissues ... 145
Figure 4.2 – Ultraviolet (UV-C) irradiation of PETG radically changes surface characteristics to allow for proper cell adhesion ... 150
Figure 4.3 – The novel mandrel-seeded approach led to the generation of mechanically superior tissue-engineered tubular constructs ... 152
Figure 4.4 – Multilayered Tissue Engineered Blood Vessels (TEBV) exhibit uniform cellular distribution, collagen-dense ECM deposition, and robust mechanical properties ... 153
Figure 4.5 – Cellular distribution, TEBV endothelization, and cell orientation ... 154
Supplementary Figure 4.1 - Cell adhesion analysis on all tested plastic polymers. ... 158
Supplementary Figure 4.2 - Cellular morphology on UV-C treated plastics. ... 159
Supplementary Figure 4.3 - Effects of UV-C treatment on cell adhesion to PETG. ... 160
Supplementary Figure 4.4 – Recorded Infrared spectra for all tested plastics. ... 161
Supplementary Figure 4.5 - Impact of failure load on different TEBVs and native umbilical artery. ... 162
Supplementary Figure 4.6 – Cellular distribution and extracellular matrix deposition within umbilical cord native artery. ... 163
Supplementary Figure 4.7 – Relative expression of type 1 collagen in TEBV and native artery detected by immunochemical analysis. ... 164
Liste des abréviations
Abréviation Signification
2D Bidimensionnelle 3D Tridimensionnel
AAA+ ATPase associée à diverses activités cellulaires
ADN Acide désoxyribonucléique ADNc ADN complémentaire ADP Adénosine diphosphate AIC Anévrisme intracrânien AIT Attaque ischémique transitoire AMPc AMP cyclique
ANG-1 Angiopoïétine-1 ANG-2 Angiopoïétine-2 ApoE Apolipoprotéine E
ARM Angiographie par résonance magnétique ARN Acide ribonucléique
ARNi ARN interférents ARNm ARN messager ATP Adénosine triphosphate AVC Accident vasculaire cérébral BHE Barrière hémato-encéphalique CAM Molécules d’adhésion cellulaires Cas Protéines associées à CRISPR Cas9n Cas9 nickase
CCR2 Récepteur de cytokines C-C de type 2 CD Cluster of differentiation
CDB Cassures double brin CE Cellule endothéliale CEF Cellule de front CES Cellules de soutien
CMEC Cellule endothéliale microvasculaire cérébrale CML Cellule musculaire lisse
crARN ARN CRISPR
CRISPR Clustered Regularly Interspaced Repeats
CSP Codon stop prématuré dCas9 Cas9 déficiente en nucléase DLL4 Delta-Like 4
DRV Dirésidues à répétition variable DSC Débit sanguin cérébral EEG Électroencéphalographie
e-NOS Synthase endothéliale de monoxyde d’azote ERK Extracellular signal-regulated kinases
FGF Facteur de croissance des fibroblastes
FLAIR Imagerie de récupération d'inversion atténuée par fluide FTIR Spectroscopie infrarouge à transformée de Fourier G-CSF Facteur stimulant les colonies de granulocytes
GLUT-1 Transporteur-1 de glucose
GM-CSF Facteur stimulant les colonies de granulocytes/macrophages HCAEC Cellule endothéliale d’artère coronarienne humaine
hCMEC Cellule endothéliale microvasculaire humaine de cerveau HGF Facteur de croissance des hépatocytes
HSA Hémorragie sous-arachnoïdienne HSPG Héparines sulfate protéoglycanes
HUVEC Cellule endothéliale de cordon ombilical humain ICAM-1 Molécules d’adhésion intercellulaire 1
IFN𝛾 Interféron gamma IL-1 Interleukine 1 IL-6 Interleukine 6 IL-8 Interleukine-8
Indels Introduction de petites mutations d’insertion ou de délétion iPSC Cellules souches pluripotentes induites
iPSEC CE vasculaires différenciées d’iPSC IRM Imagerie par résonance magnétique JA Jonctions adhérentes
JAM Molécules d’adhérence jonctionnelles JENH Jonction des extrémités non-homologues JS Jonctions serrées
LCR Liquide céphalo-rachidien LEE Lame élastique externe LEI Lame élastique interne
LFA-1 Antigène-1 associé aux fonctions lymphocytaires MAD2 Protéine déficiente en arrêt mitotique 2 MAPK Mitogen-activated protein kinase
MCAM Molécule d'adhésion au mélanome MCP-1 Protéine-1 chimiotactique des monocytes MCV Maladie cérébrovasculaire
MEC Matrice extracellulaire
MEK Mitogen-activated protein kinase kinase
MMM Maladie de Moyamoya MMP Métalloprotéinases matricielles MOI Multiplicité d’infection
NF1 Neurofibromatose de type 1
NF-κB Activateur de chaîne légère kappa du facteur nucléaire des cellules B PAM Protospacer-adjacent motif
PBMC Cellules mononucléées du sang périphérique
PC Polycarbonate
PDGFβ Facteur de croissance β dérivé des plaquettes PDGFRβ Récepteur au PDGFβ
PECAM-1 Platelet endothelial cell adhesion molecule-1
PETG Polyéthylène téréphtalate glycol
PG Plexiglas
PHc Pression hydrostatique capillaire PI3K Phosphoionositide 3-kinase
PlGF Facteur de croissance placentaire PKA Protéine kinase A
POCS Pression osmotique colloïdale du sang PPC Pression de perfusion cérébrale
PS Polystyrène
PW Polygone de Willis
RDH Réparation dirigée par homologie RET Résistance électrique transendothéliale RNF213 Ring finger protein 213
SaCas9 Cas9 de S. aureus SCV Système cérébrovasculaire SNC Système nerveux central sgARN ARN guide syntétique
Shh Sonic hedgehog
SOAIC Sténose/occlusion de l’artère principale intracrânienne SpCas9 Cas9 de S. pyogenes
TALE Effecteurs de type activateur de transcription
TALEN Nucléases effectrices de type activateur de transcription TDM Tomodensitométrie
TEBV Vaisseau sanguin reconstruit par génie tissulaire TEVA Tissue-engineered vessel adventitia
TEVM Tissue-engineered vessel media
TEVMA Tissue-engineered vessel media-adventitia
TGFβ Facteur de croissance transformant β TGFR2 Récepteur 2 au TGF-β
TIMP Inhibiteurs des MMP
TNF⍺ Facteur de nécrose tumoral alpha tracrARN ARN CRISPR trans-activant UV-C Rayons ultraviolet C
VE Vascular endothelial
VEGF Vascular endothelial growth factor
VEGFR-2 Récepteur 2 au VEGF
VCAM-1 Molécules d’adhésion cellulaire vasculaire 1 VLA-4 Antigène-4 très tardif
VMM Vaisseaux Moyamoya VWF Facteur de von Willebrand
WT Wild type
ZFN Nucléases à doigts de zinc
Remerciements
Je voudrais tout d’abord grandement remercier mon directeur de recherche, Dr François Gros-Louis, pour m’avoir accueilli dans son équipe et m’avoirconfié ces sujets de recherche pour lequels j’ai beaucoup d’intérêt. Merci de m’avoir fait autant confiance pour la progression des différents projets. Je suis très fier des avancements que j’ai pu apporter aux projets sur RNF213 dans les maladies cérébrovasculaires et sur le développement de modèles 3D pour l’étude de la Neurofibromatose de type 1. Je te remercie également de m’avoir permis de découvrir des endroits magnifiques dans le monde grâce aux nombreux congrès scientifiques auxquels j’ai pu participer. Je voudrais finalement te remercier de m’avoir offert un emploi dans ton équipe et de pouvoir rester au LOEX, ça fait chaud au cœur! Je suis très choyé de pouvoir continuer à travailler sur mes projets de recherche, de relever de nouveaux défis et de pouvoir amener mon expertise au sein de l’équipe.
Merci à mon co-directeur, Dr François A. Auger, également directeur du laboratoire et du groupe vasculaire, de m’avoir accueilli dans le groupe, de m’avoir donné de bons conseils de vie et d’avoir corrigé mon français. Merci pour l’évaluation de cette thèse et le temps que vous m’avez accordé.
Merci aux évaluateurs, Dr Guérin, Dr Dion et Dr ElAli d’avoir accepté de réviser cette thèse. Je vous suis très reconnaissant de prendre le temps et l’énergie nécessaire pour l’évaluation de ma thèse, qui est assez longue. Merci pour votre intérêt, vos commentaires et vos critiques, qui permettront d'améliorer et de compléter le contenu scientifique de ce document.
Je voudrais remercier les membres de notre équipe Lydia Touzel Deschênes, Alyssa Brodeur, Vincent Clément et Rémy Lamontagne. Merci à Lydia pour avoir réalisé plusieurs expériences et d’avoir fait certains de mes changements de milieu, tu m’as grandement aidé et as permis d’avancer davantage mes projets, c’est très apprécié. Merci également d’avoir été mon épaule supplémentaire lorsque ça allait moins bien, je ne te remercierai jamais assez. Tu m’as beaucoup aidé à passer à travers des moments plus difficiles de ma vie au laboratoire et en général. Je sais que tu sais de quoi je parle. Merci à Alyssa pour le coup de main important afin de finaliser toutes les expériences qui se retrouvent dans cette thèse. Tu as été d’une aide généreuse et je t’en remercie. Maintenant c’est à mon tour de t’aider, et toi aussi Vincent. Merci à Rémy pour les avancements importants que tu as apportés au le projet NF1, tant fondamental que clinique. Merci à Todd Galbraith de m’avoir permis de participer au projet de vaisseaus sanguins reconstruits par génie tissulaire. Grâce à notre collaboration, nous avons publié un superbe papier dans un bon journal. Merci aussi de m’avoir aidé tout au long de mon doctorat pour certaines expériences et l’avancement de mes projets. Merci à Dominic Jauvin pour son aide et ses conseils précieux pour le clonage et les RT-qPCR. Merci à Sébastien Larochelle pour son aide avec le cytomètre en flux. Merci à Marie-Josée Beaudet pour ses conseils précieux, autant en sciences que pour la vie en générale. Merci à Rémy Pépin pour son aide importante avec le cytomètre en flux, pour la conception des expériences, pour l’analyse des résultats et pour l’écriture de mes articles. Merci à Sergio Cortez Ghio pour son aide précieuse pour l’analyse des biopuces d’ARN, pour la génération de figures et pour l’écriture de mes articles. Merci à Dr François
Berthod et Dre Lucie Germain d’avoir révisé mes articles. Merci à Kim Santerre et Dre Stéphanie Proulx pour leur aide et leurs conseils avec l’analyse de résistance électrique transendothéliale. Merci à Gaëtan Le-Bel et Dr Sylvain Guérin pour la conception et l’analyse des biopuces d’ARN.
Je voudrais remercier tous les assistants de recherche qui préparent nos milieux de culture, sérums, additifs et autres. Vous facilitez énormément les tâches de chacun. Merci à tous mes amis, amies et collègues du LOEX. Je vous remercie d’apporter une joie quotidienne dans la journée de tous et chacun. J’adore l’ambiance actuelle du laboratoire et je suis fier de travailler à vos côtés. D’un autre sens, vous êtes quand même privilégiés de pouvoir travailler avec moi, évidemment. Je suis également très content d’avoir pu passer une fin de semaine mémorable tous ensemble dans un chalet avant que nos vies normales soient chamboulées par ce satané virus. En espérant que tout le monde va bien et que personne ne soit affecté directement ou indirectement. Merci aux professeurs qui m’ont motivé à poursuivre mes études toujours plus loin. Merci à Dr Robert M. Tanguay, mon directeur de recherche de maîtrise, de m’avoir inculqué une rigueur scientifique qui m’a grandement aidée lors de mon doctorat.
Finalement, un grand merci à mes parents, Nathalie et Martin, de m’avoir fortement encouragé et soutenu tout au long de mon parcours académique. Je vous aime énormément. Merci à tous mes amis qui m’ont enduré et qui continueront d’endurer mon petit caractère. Merci de m’avoir encouragé dans ce processus, d’avoir démontré un réel intérêt envers mes travaux de recherche et de m’avoir écouté et conseillé dans les moments un peu plus durs.
Avant-propos
Cette thèse comporte l’insertion de trois articles scientifiques séparée en trois différents chapitres. Le premier article (chapitre 1) a été récemment soumis dans la revue Stroke et le deuxième (chapitre 2) sera soumis éventuellement dans la revue Scientific Reports. Le troisième article (chapitres 3) a été publié dans la revue scientifique Biotechnology Journal et la version manuscrite de cet article est présentée en version intégrale. Seulement les modifications du format et de la mise en page ont été apportées afin de se conformer correctement au style de l’ouvrage déposé. Ma contribution pour chaque manuscrit est détaillée dans la section Avant-propos au début de chaque article. Deux autres manuscrits et un article de revue auquel j’ai participé à la conception, l’expérimentation et la rédaction dans le cadre de mon doctorat ont été ajoutés en annexe pour consultation. Un article de revue de la littérature a été publié dans le journal BioMed Research
International (annexe 1). Un article scientifique a été soumis dans le journal Biotechnology Journal(annexe 2)et un dernier article est également en voie d’être soumis dans la revue Scientific Reports (annexe 3).
Introduction
Problématique générale
Les maladies cérébrovasculaires (MCV) sont parmi les principales causes de mortalité et d’invalidités permanentes mondialement (Zhou et al. 2016; Benjamin et al. 2019). Elles désignent un vaste regroupement de conditions et désordres affectant les vaisseaux sanguins et l’irrigation du cerveau et du système nerveux central (SNC). Si un blocage, une malformation ou une hémorragie empêche les cellules cérébrales de recevoir suffisamment d'oxygène, des lésions cérébrales importantes peuvent en résulter. Les MCV sont de causes multifactorielles et sont influencées par des facteurs de risque vasculaires, des facteurs environnementaux et des prédispositions génétiques. Elles comprennent les accidents vasculaires cérébraux (AVC), les attaques ischémiques transitoires (AIT), les anévrismes intracrâniens (AIC) et les malformations vasculaires (Chandra et al. 2017). Des interactions interfactorielles incomprises ajoutent également un degré de complexité à la compréhension des processus pathogéniques. Les artères cérébrales et la microvasculature irriguant le SNC possèdent des caractéristiques uniques leur permettant de réguler précisément le passage des ions, des molécules et des cellules immunitaires entre la circulation sanguine et le cerveau. En effet, la barrière hémato-encéphalique (BHE) favorise le bon fonctionnement des neurones et protège le cerveau des toxines et des pathogènes (Abbott et al. 2006). L’altération de cette homéostasie peut mener progressivement au développement de diverses pathologies et maladies neurologiques.
Plusieurs études de génomique ont démontré que des facteurs génétiques peuvent avoir des effets uniques sur le développement des MCV (Della-Morte et al. 2016). Les effets phénotypiques de variations génétiques peuvent varier selon l’âge et le sexe et peuvent avoir une plus grande prédisposition chez différents groupes ethniques. Des gènes candidats ont été associés au développement d’AIC, dont certains interviennent dans la dégradation de la matrice extracellulaire, l’angiogenèse ainsi que dans la réponse inflammatoire (Zhang & Claterbuck 2008). Récemment, des variations génétiques ont été identifiées dans le gène RNF213 dans une population canadienne-française avec des histoires familiales d’AIC (Zhou et al. 2016). Toutefois, aucune caractérisation biologique n’a été concrètement réalisée. Le gène RNF213 a été précédemment associé à la maladie de Moyamoya (MMM) dans des populations de l’Asie de l’Est (Liu et al. 2011). Actuellement, les études in vitro et in vivo ne sont pas en mesure de reproduire correctement la pathologie, sauf pour certaines exceptions (Lin & Sheng 2018). Les modèles cellulaires utilisés sont moins représentatifs pour l’étude de pathologies cérébrovasculaires. Des évidences ont récemment démontré que les AIC et la MMM sont en partie des désordres de l’endothélium et des cellules endothéliales (CE) (Bang et al. 2016; Sheinberg et al. 2019). Une perturbation de la BHE serait en cause de l’initiation de la pathogenèse, mais les processus biologiques et moléculaires exacts ne sont toujours pas connus (Ivanidze et al. 2018; Narducci et al. 2019).
Le développement de modèles vasculaires in vitro robustes faciliterait la compréhension des évènements initiaux de la pathogenèse associée au gène RNF213. Au cours des dernières années, le génie tissulaire et les systèmes de culture cellulaire tridimensionnelle (3D) complexes ont acquis une grande popularité comparativement aux modèles in vivo (Kapałczyńska et al. 2018). En effet, ils reproduisent avec précision le microenvironnement et permettent un criblage de médicaments à haut débit (Li et al. 2018). La méthode d'auto-assemblage constitue désormais l'un des modèles les plus prometteurs en médecine de précision. Les substituts vasculaires de petits calibres reconstruits par auto-assemblage (TEBV) possèdent des propriétés biologiques et mécaniques similaires aux vaisseaux sanguins natifs (L’Heureux et al. 2006; Gauvin et al. 2010). Toutefois, le modèle actuel comporte certaines limitations, telles que la variabilité, la reproductibilité et le temps de production. De plus, des problèmes d’infiltration et de délamination sont régulièrement rencontrés. Pour être utilisé comme modèle pathologique et/ou pharmacologique, le vaisseau reconstruit doit être complet, avec un adventitia tunica composé de fibroblastes, une media tunica composée de cellules musculaires lisses (CML) et un intima tunica formé d’une monocouche de CE. L’optimisation du modèle permettra de générer des TEBV plus uniformes qui pourront également être utilisés comme modèle 3D in vitro pour étudier des pathologies vasculaires. De plus, le développement d’un modèle de BHE in vitro avec un système 3D sera bénéfique afin de comprendre l’impact phénotypique des variants de RNF213 sur les CE cérébrales et ainsi déterminer l’étanchéité de la barrière endothéliale en culture. Puisque l’angiogenèse et les facteurs angiogéniques semblent avoir un rôle important dans la pathogenèse des AIC et de la MMM, le développement de modèles 3D servira à caractériser l’implication de RNF213 dans le processus. La prévascularisation de substituts de fibroblastes épineuraux reconstruit par génie tissulaire avec des CE en culture 3D représenterait le modèle idéal pour l’étude de l’angiogenèse dans un environnement 3D complexe.
Le système Clustered Regularly Interspaced Short Palindromic Repeats et la protéine associée Cas9 (CRISPR-Cas9) ont été adaptés de la bactérie à l’édition ciblée du génome des cellules eucaryotes (Jinek et al. 2012). Cette méthode présente plusieurs avantages comparativement aux autres approches d’édition et semble prometteuse pour une caractérisation adéquate du gène RNF213 dans la prédisposition au développement de la MMM et la formation d’AIC. En effet, le système CRISPR-Cas9 est régulièrement utilisé pour l’invalidation de gènes d’intérêts, l’insertion de mutations ponctuelles spécifiques ou la régulation de la transcription (Araki and Ishii 2014). La génération de CE cérébrales stables n’exprimant pas la protéine RNF213 (RNF213-/-) par CRISPR-Cas9 permettra de combler la difficulté à obtenir des CE pathologiques
dérivées de patients porteurs d’une variation génétique causale. Le développement de modèles vasculaires 3D avec des CE cérébrales éditées génétiquement sera bénéfique à la fois pour la compréhension de la pathogénicité cérébrovasculaire associée aux variants de RNF213 et pour la mise au point d’éventuelles thérapies préventives en médecin de précision.
Anatomie et physiologie cérébrovasculaire
Système cérébrovasculaire
Le cerveau est l’un des organes les plus vascularisés du corps humain et nécessite des besoins énergétiques variant constamment entre les différentes régions (Chandra et al. 2017; Shah & Jeyaretna 2018). Le système cérébrovasculaire (SCV) constitue l’ensemble des vaisseaux sanguins transportant le sang à travers le cerveau et le SNC. Il est composé d’artères, d’artérioles, d’une vaste microvasculature, de veinules et de veines. Le cœur est au centre du système cardiovasculaire et pompe le sang chargé en oxygène et en nutriments vers le SCV par l’aorte ascendante, puis par deux groupes de vaisseaux: les artères carotides internes et les artères vertébrales (Figure 1.1A). Les artères carotides internes passent par le devant du cou et les artères vertébrales longent la moelle épinière. Le sang est alors distribué à travers l’ensemble des méninges par un système d’artères communicantes, appelé le Polygone de Willis (PW) (section 1.2.2).
Le PW se sépare en trois paires d’artères principales, les artères cérébrales antérieures, moyennes et postérieures, qui se divisent en artères et artérioles de plus en plus petites pénétrant les tissus cérébraux pour alimenter en sang le cortex cérébral. Alors que le cerveau représente seulement 2% de la masse corporelle totale, il est l’organe le plus énergivore et utilise pratiquement 50% du glucose (Fehm et al. 2006). L’hyperhémie fonctionnelle est le mécanisme contrôlant le SCV de façon sophistiquée et s’assurant que l’apport sanguin concorde parfaitement avec la demande métabolique des différentes régions du cerveau (Iadecola & Nedergaard 2007). Les neurones, les CE, les cellules murales et les cellules gliales établissent des interactions étroitement régulées afin de répondre rapidement à une augmentation locale de l’activité neuronale et ainsi élever le flux sanguin (Iadecola 2004). La vasoréactivité cérébrale s’assure de conserver un débit sanguin cérébral (DSC) relativement constant lors d’une augmentation indésirable de la pression sanguine (Dahl et al. 1992; Silvestrini et al. 2000).
L’oxygénation des tissus et l’échange fluidique se produisent majoritairement dans les capillaires. La microvasculature cérébrale diffère grandement des capillaires extracrâniens. En effet, la BHE permet de contrôler sélectivement et précisément l’homéostasie entre la circulation sanguine et le SNC (section 1.3) (Abbott et al. 2010). Les molécules et les cellules immunitaires empruntent différentes voies de transport qui leur sont propres afin de pénétrer la BHE (section 1.3.3). Généralement, les échanges de fluides nécessitent l’intervention de deux mécanismes dépendants de la pression: la filtration et la réabsorption. La filtration nette se produit près de l’extrémité artérielle des capillaires, car la pression hydrostatique capillaire (PHc) est supérieure à la pression osmotique colloïdale du sang (POCS). Puisque la PHc est égale à la POCS, il n’y a pas de mouvement fluidique au point médian des capillaires. La réabsorption nette a lieu près de l’extrémité veineuse, là où la POCS est devenue supérieure à la PHc.
Le sang concentré en dioxyde de carbone et en déchets cellulaires revient dans la circulation cardiovasculaire par un système de veines et sinus intracrâniens (Figure 1.1B) (Kılıç & Akakın 2007). Les veines cérébrales sont dépourvues de valve et d’une paroi musculaire. Deux groupes de veines s’occupent du drainage cérébral: les veines corticales superficielles et profondes. Les veines corticales superficielles sont situées dans la pie-mère à la surface et drainent le cortex cérébral et la substance blanche sous-corticale. Les veines profondes se sous-divisent en veines sous-épendymales, cérébrales internes, basales et en grande veine de Galen. Ces veines drainent la matière blanche et grise profonde à l’intérieur du cerveau pour ensuite s’anastomoser avec les veines corticales et se vider dans le sinus sagittal supérieur. Par la suite, les veines profondes et le sinus sagittal supérieur se connectent aux sinus sigmoïdes et veines jugulaires. Le cervelet est principalement drainé par les veines cérébelleuses inférieures et les sinus occipitaux et le tronc cérébral est drainé par des veines se terminant par les sinus pétreux inférieurs et transversaux. Un dysfonctionnement ou une anomalie dans le système cérébrovasculaire peut mener au développement de maladies cérébrovascualires (section 1.5).
Figure 1.1 – Les principales artères et veines du système cérébrovasculaire
A) Distribution des artères cérébrales approvisionnant en sang la tête et le cou. L’artère carotide commune devient l’artères carotides
internes, qui irrigue les sinus et le crâne, et l’artère carotide externe, qui reste superficielle et se sépare en de nombreuses artères vascularisant la tête. Les artères vertébrales se ramifie à partir de l'artère sous-clavière, traverse le foramen transversal où les vertèbres cervicales et entre à la base du crâne où le foramen vertébral. B) Distribution des veines cérébrales drainant le sang usé de la tête et du cou. (Adaptée de (Gordon et al. 2013))
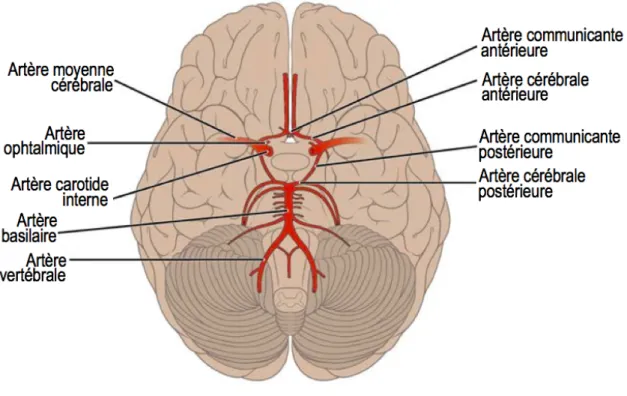
Polygone de Willis
Le PW, aussi appelé le cercle de Willis, est un circuit hexagonal d’artères anastomosées se situant à la base du cerveau et permettant l’approvisionnement adéquat en sang pour le fonctionnement de ce dernier et des structures avoisinantes. Il y a approximativement 400 ans, une description de l’anatomie de ce cercle artérielle a été détaillée par le médecin Tomas Willis (Lo & Ellis 2010). Il a aussi décrit sa fonction de mécanisme compensatoire en cas d’occlusion ou de sténose des artères carotides internes ou des artères vertébrales. Le cercle de Willis est composé des artères carotides internes, cérébrales antérieures, communicante antérieure, postérieures communicantes, cérébrales postérieures et basilaires (Figure 1.2). En effet, cette configuration permet une distribution du sang entrant de l’artère carotide interne ou de l’artère vertébralevers n’importe quelle partie des deux hémisphères et les flux sanguins se rejoignent à l'artère communicante postérieure. À ce point, la pression est équivalente et le sang ne se mélange pas.
Figure 1.2 – Le polygone de Willis
Schéma en vue inférieure illustrant les artères desservant le cerveau et le SNC en sang. La structure est un circuit hexagonal d’artères anastomosées qui se situe à la base du cerveau. Cette configuration permet une distribution adéquate du sang qui entre par les artères carotides internes ou des artères vertébrales vers les deux hémisphères et les flux sanguins se rejoignent aux artères communicantes postérieures. (Adaptée de (Gordon et al. 2013))
Anatomie
Les branchements terminaux de la circulation antérieure et postérieure s’anastomosent pour former la structure du cercle de Willis à la base du cerveau (Liebeskind & Caplan 2017). Le PW est composé de cinq artères principales: les artères carotides internes, les artères cérébrales antérieures, l’artère communicante antérieure, les artères cérébrales postérieures et les artères communicantes postérieures (Fig. 1.2). Il est important de noter que l’anatomie du PW n’est pas le même chez tous les individus (Tableau 1.1). En effet, des anomalies anatomiques sont observées chez près de 50% de la population (section 1.2.2.4). Du côté de la circulation antérieure, les artères carotides internes sont le prolongement de l’artère carotide commune et chaque artère carotide interne fournit environ 40% du total de la perfusion cérébrale. Les artères carotides internes gauche et droite se prolongent pour former les artères cérébrales moyennes et se développent en artères olfactives primitives et en artères cérébrales antérieures. Les artères cérébrales moyennes fournissent le sang dans la plus grande partie de la circulation intracrânienne. Les branchements des artères cérébrales moyennes perfusent le frontal, le pariétal et une certaine partie des cortex temporaux et occipitaux. Les artères olfactives primitives de chaque côté forment un plexus dans la ligne médiane qui donne naissance à l’artère communicante antérieure. Les artères cérébrales antérieures s’étendent entre le chiasma optique et la substance perforée antérieure pour se rejoindre et s’anastomoser par le biais de l’artère communicante antérieure. Du côté de la circulation postérieure, les artères vertébrales sont le prolongement des artères sous-clavières qui s’anastomosent pour former l’artère basilaire. Le plus grand affluent des artères vertébrales est l’artère cérébelleuse inférieure postérieure, qui apparait un peu avant la jonction vertébrobasilaire. L’artère basilaire se divise en plusieurs branchements tels que l’artère cérébelleuse inférieure antérieure, les artères pontines et l’artère cérébelleuse supérieure. L’artère basilaire finit par se connecter au PW par les artères cérébrales postérieures et les artères communicantes postérieures. Les artères cérébrales postérieures se forment de la fusion de plusieurs vaisseaux provenant de la partie terminale des artères communicantes postérieures. Finalement, les artères communicantes postérieures se connectent aux artères carotides internes avant la bifurcation terminale entre l’artère cérébrale antérieure et l’artère cérébrale moyenne pour ainsi fermer le cercle.
Tableau 1.1 – Taille et diamètre externe moyens des artères principales du cercle de Willis
Nom de l’artère
Taille (mm) Diamètre (mm) Gauche Droite Gauche Droite
Artère carotide interne 4,7 ± 1,5 4,8 ± 1,5 4,2 ± 0,9 4,2 ± 0,9 Artère cérébrale antérieure 13,8 ± 2,7 14,7 ± 3,0 2,4 ± 0,5 2,2 ± 0,6 Artère communicante antérieure 2,5 ± 1,8 1,9 ± 0,9
Artère cérébrale postérieure 6,9 ± 3,1 6,8 ± 2,7 2,2 ± 0,6 2,1 ± 0,7 Artère communicante
postérieure 13,3 ± 3,3 13,5 ± 2,7 1,4 ± 0,7 1,5 ± 0,7 (Adapté de (Kamath 1981))
Fonction biologique
Le PW a pour fonction théorique de maintenir le débit sanguin cérébral en cas d’occlusion ou de sténose des artères carotides internes ou des artères vertébrales (Nornes 1973; Edelman et al. 1990). En cas de sténose progressive d’une artère cérébrale principale, les artères communicantes s’adaptent avec le temps et s’élargissent afin de maintenir la perfusion des deux hémisphères cérébraux. En conditions pathologiques, le PW agit à titre de valve de sécurité permettant une circulation collatérale du sang si le débit est réduit dans une zone du cerveau. En conditions physiologiques, le cercle de Willis est stabilisé et ne fonctionne théoriquement pas. Certains ont une école de pensées différentes et croient que la théorie actuelle de la fonction compensatoire du PW semble être incorrecte d’un point de vue évolutif (Vrselja et al. 2014). En effet, l'évolution produit des organismes avec des spécificités structurelles qui sont adaptées à des environnements spécifiques. L'évolution conduit à une structure anatomique spécifique qui entrainera des pathologies spécifiques et non l'inverse. Par conséquent, le processus pathologique ne forcerait pas les espèces à s'adapter; cela conduirait plutôt à son extinction. Il est plus probable que les contraintes et les charges physiques agissent comme une pression sélective dans certaines circonstances. Bien évidemment, les deux théories ont du sens, mais un débat plus approfondit permetterait d’avoir une meilleure compréhension.
Le PW fonctionne aussi comme un mécanisme d’absorption de la pression afin d’empêcher des lésions de la microvasculature intracrânienne et de la BHE (Kontos et al. 1978; Armstead 2016). Lors d’une forte augmentation de la pression sanguine, les plus grandes artères cérébrales subissent plus de pression afin de protéger la microcirculation. Les bifurcations des artères intracrâniennes ont une structure plus perpendiculaire comparativement aux artères extracrâniennes de taille similaire afin d’assurer une réduction de la vélocité sanguine, un meilleur transfert de l’énergie cinétique et une augmentation de la PHc (Donnelly et al. 2016). De plus, les artères cérébrales se ramifient plus progressivement en plus petits vaisseaux que les artères systémiques afin d’obtenir une résistance cérébrovasculaires plus puissante. Au repos, la résistance se concentre aux artérioles en aval des artères cérébrales moyennes et, lors d’un exercice, la résistance se déplace vers les plus grosses artères du PW (Kontos et al. 1978; Willie & Ainslie 2011).
Autorégulation de la circulation et de la pression
Le débit sanguin cérébral (DSC) est un processus biologique régulé de façon très stricte. La moyenne normale du DSC de l’entièreté du cerveau est d’environ 50 ml/100g/min (Lassen 1985). La perturbation du débit sanguin affecte directement le métabolisme cérébral. Un DSC inférieur à 35 ml/100g/min est accompagné d’une diminution de la synthèse protéique (Hossmann 1994). Toutefois, les fonctions neurologiques restent inaffectées (Marshall et al. 2001). Des déficits neurologiques deviennent évidents lorsque le DSC est sous les 27 ml/100g/min et s’il demeure sous les 15 ml/100g/min pendant près d’une heure, le patient subira
inévitablement un infarctus du myocarde permanent (Pulsinelli 1992; Marshall et al. 2001). Lorsque le DSC demeure inférieur aux valeurs physiologiques normales, des dommages ischémiques pouvant être irréversibles se produisent. Le débit peut être influencé par plusieurs facteurs tels la pression artérielle moyenne (MAP), le métabolisme neurologique, les facteurs chimiques et les entrées synaptiques (Vavilala et al. 2002). Malgré tous les changements dynamiques que peut subir le cerveau, le DSC est maintenu constant par le processus d’autorégulation cérébrale. L’autorégulation cérébrale contrôle, entre autres la pression de perfusion cérébrale (PPC) pour chaque région. La PPC est le gradient de pression net dirigeant le flux sanguin vers le cerveau. L’autorégulation compense les variations de la PPC par la vasoconstriction et la vasodilatation des artères pour modifier la résistance des vaisseaux en aval et ainsi maintenir le DSC. Ce processus demeure fonctionnel lorsque la MAP varie entre 50 et 150 mmHg (Lassen 1959). Par exemple, une MCV occlusive peut entrainer une sténose obstruant partiellement ou totalement d’un vaisseau, réduisant ainsi la PPC. L'autorégulation entraine alors une dilatation des artères en aval afin de rétablir un DSC suffisant pour maintenir la fonction métabolique des cellules neuronales.
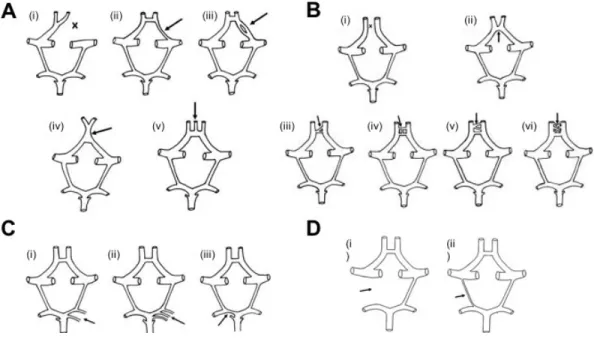
Variations de la configuration
Des changements de la morphologie du PW sont régulièrement retrouvés dans la population. Ces variations anatomiques sont généralement d’origine génétique, se développent au stade embryonnaire précoce et persistent dans la vie post-natale (Milenković et al. 1985). D’autres modificateurs, comme les facteurs environnementaux, le stress hémodynamique et l’amplitude des mouvements du cou, peuvent également être en cause (Hillen 1987). Les changements de la configuration du PW se manifestent par l’absence, de l’hypoplasticité, une duplication, une triplication ou même du pleximorphisme d’une artère cérébrale (Figure 1.3) (Kapoor et al. 2008). Chez certains individus, il n’est pas rare de retrouver une combinaison de deux ou plusieurs types de variations. L’âge et le sexe n’ont aucun effet sur l’incidence des variations. Le taux d’anomalies est significativement plus élevé chez les patients atteints de troubles mentaux ou de pathologies cérébrovasculaires, tels qu’un AVC ou un AIC (Iqbal 2013). Dans une étude menée sur des patients avec un ou plusieurs anévrismes, des variations de la configuration de la vasculature intracrânienne ont été identifiées dans 57,8% des cas (Orakdogen et al. 2016). Dans une autre étude, 40,7% des patients avec un anévrisme avaient aussi des anomalies du PW (Lazzaro et al. 2012). De plus, ils ont démontré que les variations sont plus fréquentes dans des AIC rompus (46,9%) comparativement aux patients avec des AIC non rompus (29,6%). En effet, il est clair qu’une configuration anormale du cercle de Willis joue un rôle dans la manifestation des symptômes cliniques. Par ailleurs, il a été démontré que le DSC est déterminé par l’anatomie du PW (Hartkamp et al. 1999). Une connaissance détaillée des variations des patients serait bénéfique aux chirurgiens afin de planifier adéquatement les stratégies à adopter lors des opérations vasculaires. Une telle démarche permettrait de prévenir les traumatismes vasculaires involontaires.
Figure 1.3 – Exemples d’anomalies de la configuration du cercle de Willis
A) Illustrations schématisées des anomalies retrouvées de l’artère cérébrale antérieure: (i) absence; (ii) hypoplasticité; (iii) scission;
(iv) fusion; (v) triple artère cérébrale antérieure. B) Illustrations schématisées des variations de la configuration retrouvées dans l’artère communicante antérieure: (i-ii) absence; (iii) duplication; (iv-v) triplication; (vi) plexiforme. C) Illustrations schématisées des anomalies du segment proximal de l'artère cérébrale postérieure: (i) duplication; (ii) triplication; (iii) hypoplasticité. D) Illustrations schématisées des variations de la configuration retrouvées dans l’artère communicante postérieure: (i) absence; (ii) hypoplasticité. (Adaptée de (Kapoor et al. 2008))
Physiologie et structure des artères cérébrales
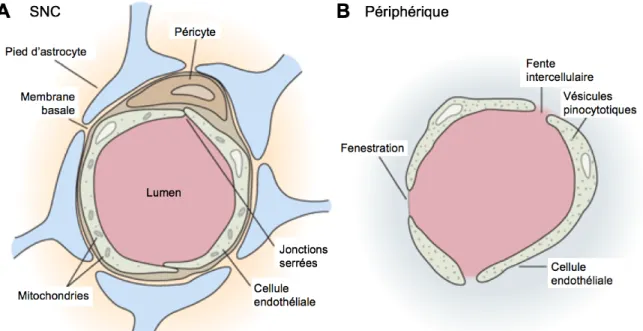
La structure des artères intracrâniennes diffère des artères extracrâniennes sur plusieurs aspects physiologiques (Figure 1.4). Les artères cérébrales sont de type musculaire et possèdent une media tunica et une adventicia tunica généralement plus mince (Moossy 1966; Velican 1970). De plus, la media tunica contient peu de fibres élastiques comparativement aux artères périphériques. Les artères intracrâniennes ne possèdent pas de lame élastique externe (LEE), ont une lame élastique interne (LEI) beaucoup plus épaisse et dense et n’ont généralement pas de vasa vasorum, sauf pour certaines exceptions (Portanova et al. 2013). Elles sont entourées de liquide céphalo-rachidien (LCR) qui est très riche en nutriment. Les CE se distinguent par leur composition et leur enrichissement en jonctions intercellulaires et par la réduction importante de la perméabilité vasculaire, qui sont des caractéristiques particulières contrôlées par la BHE (Hladky and Barrand 2016). Les artères cérébrales ont également un mécanisme plus performant pour se protéger contre les dommages provoqués par le stress oxydatif et sont relativement insensibles aux stimulations sympathomimétiques et à l’histamine (Edvinsson & Owman 1975; Napoli et al. 1999). Le DSC est également hautement régulé et dicté en fonction de l’anatomie du PW (Hartkamp et al. 1999).
Figure 1.4 – Les structure majeures des artères intracrâniennes et extracrâniennes
Les artères intracrâniennes ont une media tunica et une adventitia tunica plus minces, peu de fibres élastiques et une LEI plus épaisse. Elles n’ont pas de LEE et généralement pas de vasa vasorum. De plus, les jonctions intercellulaires des CE de l’intima tunica sont beaucoup plus importantes dans les artères intracrâniennes. Les CE ont également une composition distincte en glycocalyx du côté luminal de leur membrane (Adaptée de (Ritz et al. 2014))
Adventitia tunica
L’adventicia tunica est la couche la plus externe des artères et est majoritairement composée de fibroblastes empaquetés dans un réseau dense de fibres d’élastine et de collagène (Lee 1995). Généralement, les fibroblastes et les fibres de la matrice extracellulaire (MEC) sont alignés de façon longitudinale à l’axe du vaisseau (Smith et al. 1981). L’adventitia tunica sert de couche protectrice et permet de limiter la déformation de la paroi artérielle. La surface externe des artères intracrâniennes est en contact direct avec le LCR environnant, ce qui facilite les échanges avec la circulation sanguine (Sakka et al. 2011). Le tissu conjonctif de l’adventitia protège les nerfs autonomes, qui ont également des connexions avec la media et avec les CE de l’endothélium (Bleys et al. 1996).
Media tunica
La media tunica est principalement constituée de cellules musculaires lisses (CML), de fibres d’élastine, de collagène et de proteoglycane (Mérei et al. 1980). L’abondance en fibres élastiques varie d’un type de vaisseau à l’autre, mais sont moins exprimées par les cellules des artères cérébrales. Les CML sont liées entre elles par des jonctions intercellulaires et sont pratiquement alignées circonférentiellement autour de la lumière des vaisseaux (Wagenseil and Mecham 2009). L’organisation de la MEC et des CML permet une répartition plus uniforme des contraintes mécaniques et une meilleure résistance à une forte pression
sanguine. Les fibres musculaires de la media sont responsables de la modulation locale du diamètre artériel en contrôlant la vasodilatation et la vascoconstriction (Brozovich et al. 2016). L’organisation circonférentielle des CML sert à limiter les besoins énergétiques nécessaires au processus de contraction. Les terminaisons nerveuses de la media sont en contact avec les CML et sécrètent de l’épinéphrine agissant sur les récepteurs β2afin de contrôle ce processus (Zhang et al. 1990; Purkayastha & Raven 2011). Les CML de la media tunica et les CE de l’intima tunica sont en interaction dynamique, ce qui permet de réguler plus rapidement les processus physiologiques associés au système circulatoire (Qiu et al. 2014; Li et al. 2018).
Intima tunica
L’intima tunica est formée d’une monocouche de CE vasculaires qui sont supportées par une épaisse LEI (Wagenseil & Mecham 2009). Les CE sont directement en contact avec le sang du côté apical et reposent sur une membrane basale (section 1.3.1.3). Les CE contribuent à la formation de la LEI par la production d’élastine en réponse aux signaux envoyés par les CML de la media (Cantor et al. 1980; Mecham et al. 1983). De plus, les CE sécrètent de nombreux facteurs influençant les CML, afin de contrôler indirectement la vasodilatation et la vasoconstriction des artères (Félétou & Vanhoutte 2009). Les CE de l’endothélium artériel sont généralement de forme allongée et sont également alignées dans le sens de la circulation sanguine (Eskin et al. 1984). La membrane plasmique de la région apicale des CE cérébrales se distingue grandement par sa composition en glycoprotéines, glycolipides et en métabolites (Kutuzov et al. 2018).Cette particularité est aussi connue pour influencer le métabolisme de la paroi vasculaire. Comparativement à l'endothélium des artères périphériques, l'endothélium cérébral a moins de cavéoles et un transport vésiculaire nettement réduit (Fung et al. 2018). Finalement, les CE cérébrales sont liées entre elles par d’étanches jonctions serrées (JS) qui leur permettent de contrôler sélectivement la perméabilité vasculaire (Wolburg & Lippoldt 2002).
Vasa vasorum intracrânien
La vaste majorité des artères extracrâniennes sont aussi irriguées par un réseau de micro-capillaires sanguins, appelé le vasa vasorum. Par contre, l’existence d’un tel réseau dans les artères intracrâniennes demeure encore nébuleuse. Une étude réalisée sur des artères cérébrales majeures post-mortem a permis de conclure qu’un vasa vasorum est seulement présent chez certains individus et que sa distribution peut aussi varier (Takaba et al. 1998). En effet, la microvasculature peut être localisée dans la tunica adventitia ou même plus rarement dans la tunica media. De plus, l'existence d’un vasa vasorum est plus courante dans les artères proximales que dans les artères distales. Le développement d’un tel réseau dans les artères cérébrales est plus fréquent chez les personnes âgées et corrèle avec le développement de MCV et d’athérosclérose sévère (Portanova et al. 2013). Par ailleurs, le développement de vasa vasorum dans la tunica media peut être associé avec certains changements pathologiques des artères cérébrales.
Barrière hémato-encéphalique
Le SNC est principalement constitué du cerveau et de la moelle épinière et régule les réponses du corps à des stimuli internes et externes. Les neurones sont au centre de cette fonction et nécessitent un contrôle étroit des signaux électrophysilogiques et chimiques pour un maximum d’efficacité. La régulation précise du microenvironnement neuronal est essentielle pour le maintien de l’homéostasie du SNC, de l’intégrité des neurones et d’un signal synaptique optimal. Il existe 3 barrières cellulaires qui forment une interface entre la circulation sanguine et les tissus neuronaux: la barrière sang-liquide céphalo-rachidien (BSLCR), la barrière arachnoïdienne et la barrière hémato-encéphalique (BHE) (Figure 1.5). La barrière sang-liquide céphalo-rachidien est formée de cellules épithéliales du plexus choroïde (Abbott et al. 2006). Cet épithélium sécrète le LCR, qui remplit les espaces cérébral et spinal sous-arachnoïdiens et les ventricules afin de protéger le cerveau, et réguler le débit sanguin et les échanges. La barrière arachnoïdienne est formée de l’épithélium arachnoïdien et sert de scellant entre le LCR et les autres fluides extracellulaires (Abbott et al. 2010).
Figure 1.5 – Les barrières du SNC
A) La BHE est située à l'endothélium qui tapisse la microvasculature cérébrale et qui constitue le principal site d'échange moléculaire
entre le sang et le cerveau. B) La barrière sang-liquide céphalo-rachidien se retrouve le long de l’épithélium du plexus choroïde et permet de séparer le LCR sécrété par les cellules épithéliales du reste de la vasculature cérébrale et des tissus neuronaux. La barrière arachnoïdienne, qui n’est pas illustrée ci-dessous, est formée par la couche arachnoïde des méninges qui entoure complètement le cerveau. Pour chaque barrière du SNC, des complexes de jonctions serrées aident à limiter la perméabilité paracellulaire. Les jonctions serrées de la BHE forment la barrière la plus serrée et la plus régulée. (Adaptée de (Deczkowska et al. 2016))