Pépite | Caractérisation du rôle de la O-N-acétyl-glucosaminylation dans la structuration sarcomérique du muscle squelettique et de son implication dans certaines pathologies musculaires
309
0
0
Texte intégral
(2) Thèse de Matthias Lambert, Lille 1, 2016. Université de Lille 1 : Sciences et Technologies Ecole Doctorale Biologie-Santé (ED 446). THÈSE de Doctorat d’Université Présentée par. Matthias LAMBERT Pour l’obtention du grade de. Docteur de l’Université de Lille, Sciences et Technologies Spécialité « Physiologie ». Caractérisation du rôle de la O-N-acétyl-glucosaminylation dans la structuration sarcomérique du muscle squelettique et de son implication dans certaines pathologies musculaires. Soutenue publiquement le 27 Septembre 2016 devant le jury composé de : M. Marc BITOUN INSERM - Chargé de recherche, Institut de Myologie, France. Rapporteur. M. Zhenlin LI INSERM - Directeur de Recherche, Université Pierre et Marie Curie, France. Rapporteur. Mme. Laëtitia GUEVEL Enseignant-Chercheur HDR, Université de Nantes, France. Examinatrice. M. Tony LEFEBVRE Professeur, Université de Lille, France. Examinateur. M. Bruno BASTIDE Professeur, Unversité de Lille, France. Directeur. Mme. Caroline CIENIEWSKI-BERNARD Enseignant-Checheur HDR, Université de Lille, France. Co-Directrice. © 2016 Tous droits réservés.. lilliad.univ-lille.fr.
(3) Thèse de Matthias Lambert, Lille 1, 2016. A ma maman, Qui m’a donné les armes pour combattre. « Ne doutez jamais qu’un petit groupe de personnes puisse changer le monde. En fait, c’est toujours ainsi que le monde a changé » - Margaret Mead, anthropologue. © 2016 Tous droits réservés.. lilliad.univ-lille.fr.
(4) Thèse de Matthias Lambert, Lille 1, 2016. Le travail de thèse présenté dans ce manuscrit a été réalisé sous la direction du Professeur Bruno BASTIDE, et sous la co-direction du Docteur Caroline CIENIEWSKI-BERNARD au sein de l’équipe Activité Physique, Muscle, Santé, de l’Unité de Recherche Pluridisciplinaire Sport, Santé, Société (URePSSS - EA7369). Cette thèse a commencé en Octobre 2013 et a bénéficié d’une allocation doctorale handicap du Ministère de la Recherche et de l’Enseignement Supérieur.. © 2016 Tous droits réservés.. lilliad.univ-lille.fr.
(5) Thèse de Matthias Lambert, Lille 1, 2016. Remerciements Je remercie tout d’abord le Pr Serge Berthoin, directeur de l’Unité de Recherche Pluridisciplinaire Sport, Santé, Société (URePSSS) de m’avoir accueilli au sein de son laboratoire me permettant ainsi de réaliser cette thèse. J’ai tout particulièrement apprécié ta disponibilité et ta gentillesse qui sont de grandes qualités à mes yeux. Je tiens à remercier le Pr Bruno Bastide, qui a été mon directeur de thèse pendant ces 3 ans, et qui m’a ouvert les portes du SN4 lors de ma licence 3, il y a un peu plus de 5 ans…Le temps passe vite. Je te remercie de m’avoir permis de réaliser cette thèse dans de bonnes conditions à la fois au SN4 et à Eurasport, mais aussi en dehors lors des différents déplacements en congrès où parfois mes jambes ne pouvaient pas me porter. Je tiens d’ailleurs à m’excuser pour les poignets du fauteuil qui étaient trop basses, j’ai pris note pour la nouvelle commande. Je te remercie donc de m’avoir « poussé » dans tous les sens du terme je crois qu’on peut le dire, et d’avoir cru en moi jusqu’au bout, malgré parfois mon caractère mais qui je pense s’avère nécessaire pour que je maintienne un cap et les ambitions qui sont les miennes. Je tacherai ainsi d’exporter le « made in SN4 » que tu m’as inculqué pour mon futur post-doc, je l’espère de la meilleure des façons possibles. Ensuite, je souhaite énormément remercier le Dr Caroline Cieniewski-Bernard, qui a été mon encadrante et ma co-directrice de thèse. Voilà maintenant un peu plus de 5 ans que je connais grâce à toi le terme « O-GlcNAcylation », et je dois l’avouer, au départ, j’ai eu un peu de mal à le prononcer car je pensais qu’il manquait des voyelles, et je ne comprenais pas pourquoi il y avait tant de majuscules. Depuis, tu m’as véritablement enseigné les fondements de ce champ de recherche que ce soit en terme de connaissances ou de techniques expérimentales. Je te remercie donc beaucoup pour ton enseignement qui est d’une très grande qualité et de m’avoir appris tant de choses. Je te remercie également pour ta disponibilité, ton aide précieuse, ton écoute, ta confiance, et ta gentillesse. Tu m’as permis de travailler sereinement, dans la bonne humeur et avec passion. Comme j’aime à le dire, quand on a un numéro 10 (ou Zidane pour les non connaisseurs de football) au laboratoire, on ne peut que gagner, et je pense que cette thèse a été gagnante grâce à toi. Par ailleurs, je voudrai aussi te remercier de m’avoir nourri pendant ces années, les petits sablés et les délicieux cookies vont me manquer. Enfin Je te remercie encore de m’avoir toujours soutenu et encouragé durant cette thèse, mais aussi pour l’après. J’ai eu à cœur de bien travailler et j’espère m’appliquer aussi bien par la suite grâce à tes enseignements. Mes remerciements vont également au Pr Tony Lefebvre et au Dr Marc Bitoun pour avoir accepté d’être membres de mon Comité de Suivi de Thèse afin d’évaluer mon travail et pour m’avoir apporté de nombreux conseils. Je vous remercie sincèrement pour votre professionnalisme, votre aide, vos encouragements mais aussi pour vos recommandations quant à la recherche d’un post-doctorat. Concernant ce manuscrit et ma future soutenance de thèse, je remercie le Dr Zhenlin Li et le Dr Marc Bitoun pour m’avoir fait l’honneur de faire partie de mon jury afin de juger mon travail de thèse. J’associe à ces remerciements le Dr Laëtitia Guével et le Pr Tony Lefebvre pour avoir accepté d’examiner ce travail. Mes remerciements vont aussi aux personnes qui ont participés à mes travaux de thèse. En premier lieu, le Dr Barbara Deracinois (c’est un peu trop protocolaire, Barbie c’est mieux), je te remercie pour toute ton aide que ce soit en culture ou à la paillasse, mais aussi pour tes conseils. Tu m’as connu au laboratoire pendant mon master et mon début de thèse, je pense que j’ai bien grandi depuis…Et je crois que les enfants tu connais maintenant. Je te remercie sincèrement pour ta gentillesse, ta bonne humeur, et ce. © 2016 Tous droits réservés.. lilliad.univ-lille.fr.
(6) Thèse de Matthias Lambert, Lille 1, 2016. instant de gloire partagé à la télévision (un peu sous l’effet de surprise je te l’accorde, mais j’ai gardé la copie de l’enregistrement). Ensuite, je remercie le Dr Erwan Dupont, pour toute son aide également. Je dirai aussi que les enfants, tu connais, et j’espère que tu as pu voir mon évolution depuis ma 1 ère année de licence où tu m’as eu comme étudiant, jusqu’à maintenant. Je te remercie aussi pour ta disponibilité et tes conseils…Mais aussi pour tes blagues (du style « Paul Hochon », j’ai encore un peu de mal à m’en remettre). Par ailleurs, je remercie Elodie Richard, pour ces innombrables heures passées en microscopie confocale, à chercher et à mesurer toutes ces stries. Ça a été un boulot titanesque mais payant au final, chapeau bas ! Je veux aussi te remercier pour tes conseils et ta grande disponibilité. Je souhaite également remercier le Dr Sophie Duban-Deweer pour les (très) nombreuses analyses en spectrométrie de masse et le Dr Krzewinski pour l’HPLC, où je pense avoir passé 3 mois de ma thèse au C9 ; merci pour cet accueil et ces conseils. Plus généralement, je souhaite adresser mes remerciements chaleureux aux personnels du laboratoire que j’ai côtoyés tout d’abord au SN4 puis à Eurasport. En premier lieu, les techniciennes Valérie, Laëtitia, Estelle qui m’ont vu grandir. Je remercie également Annie pour son aide extrêmement précieuse dans le laboratoire et pour sa très grande gentillesse. Je souhaite aussi remercier les secrétaires, Delphine puis Emilie pour leur aide. Emilie, je retiendrai aussi ces histoires que je nommerai « le boulanger » ou « l’homme du train », je vais en rire encore pendant des années je pense. Je remercie aussi le Pr Laurence Stevens, le Pr Marie-Hèlene Canu, et le Dr Florence Picquet pour leur gentillesse et leur disponibilité. Cela s’adresse également au Dr Luc Dannhoffer, même si Radio campus m’a cassé les oreilles. Enfin, mes remerciements et ma sympathie vont à Fabrice et Jean-Pierre, mais aussi à Arnaud et Jessica pour les TP. Je remercie également les personnes du côté STAPS que j’ai un peu moyen côtoyées certes, mais pour qui j’ai énormément de sympathie et que je commence à connaitre avec cette fusion des équipes à Eurasport. Par ailleurs, je tiens à remercier spécialement le Dr Vincent Tiffreau, avec qui je suis passé du statut « patient » au statut « collègue ». Je pense qu’il y a une découverte qu’on aurait pu rajouter dans cette thèse, celle du gène muté de cette myopathie après 25 ans de recherches. C’est une des belles histoires de cette thèse, ou je dirai plutôt « aventure ». Je te remercie également pour ton engagement dans la sensibilisation du handicap. Je pense que c’est très important en tant que docteur mais aussi chercheur travaillant dans ce domaine d’y être confronté, et de se rendre compte d’un monde qu’on pense connaitre. J’adresse également un énorme merci aux étudiants et doctorants qui sont passés ou qui sont encore au laboratoire et que j’ai pu côtoyer, à savoir François-Xavier, Paul, Julie (la brune), Camille, Giada, Amandine, Julie (la blonde), Samir, le Dr Julien Mysoet, le Dr Semah Tagougui, Baptiste, Mehdi, Agathe,...Désolé pour celles ou ceux que j’ai oubliés, j’ai une fâcheuse tendance à oublier les prénoms (ou alors c’est une simple excuse). Merci pour tous ces moments passés, ils resteront gravés quoi qu’ils en soient, et merci de m’avoir supporté, moi et mes blagues, elles vous manquent déjà j’en suis sûr. En particulier, Julien, merci de m’avoir accueilli dans le bureau des étudiants quand tu étais en master, et de m’avoir permis de parfaitement m’intégrer. Je pense que ça a été un des points clé de la réussite de mon travail, et je t’en remercie. Je te souhaite que du bon pour la suite. Amandine, merci de ton aide au quotidien au labo, de m’avoir supporté (ça c’est un gros boulot), et d’avoir participé à la team O-GlcNAc. Je sais que la chance n’a pas souri mais elle tournera, crois moi, j’en suis sûr. Julie (la brune qui se reconnaitra forcément), depuis ton passage de 4 semaines au labo il y a deux ans, jusqu’à maintenant, il s’en est passé des choses. Je te remercie de m’avoir toujours soutenu et d’avoir cru en moi. Grâce à toi, j’ai dépassé encore un peu plus mes limites, et tu m’as convaincu définitivement d’une chose, c’est d’aller jusqu’au bout de ses idées, même si cette chose parait impossible à atteindre. Je tiens à remercier les deux petites têtes que sont Julie (la blonde cette fois-ci) et Samir, je crois qu’on a lié une belle amitié. Merci pour votre. © 2016 Tous droits réservés.. lilliad.univ-lille.fr.
(7) Thèse de Matthias Lambert, Lille 1, 2016. aide au quotidien, votre bonne humeur, et pour ce vent de fraicheur dans le labo. Vous savez ce qu’il vous reste à faire durant les 2 ans à venir. Samir, je te remercie pour tout le boulot au labo et à la salle de muscu. Oui cette personne a essayé de muscler un myopathe, et ça a fonctionné. Je propose de publier les résultats. En tout cas, désolé de ne pas aller dans les douches collectives, ma religion me l’interdit, mais j’ai pris plaisir à bosser avec toi, partenaire de salle. Enfin, Alex, oui maintenant tu fais partie du labo, je te remercie pour cette belle amitié qui est née en 1ère année de Licence. Maintenant on se retrouve à soutenir notre thèse dans le même labo à 3 jours d’écart. Notre amitié est passée par Los Angeles et Vegas, les tribunes du LOSC, et la rue Solférino (peut-être un peu trop). Je souhaite aussi remercier le reste de mes amis qui ont été le plus présent, à savoir Clémentine, Emeline, Justine, Jérémie, Julie (la rousse maintenant), mais aussi les futurs docteurs de ma promo, à savoir Anthony, Camille, Alban (Je n’oublierai pas toutes ces soirées et ses sorties, avec toi et Alex, merci d’avoir toujours été là, et d’avoir répondu présent). Je remercie spécialement Vanessa (future prof des écoles, j’en suis certain). Je te remercie beaucoup pour m’avoir toujours soutenu, dans les moments drôles et les moments un peu moins drôles. Enfin, je remercie un ami d’enfance qui m’est très cher, Clément, toujours fidèle au poste. Les après-midis Fifa étaient un peu moins présentes durant cette thèse, mais tu as toujours été là. Il y a des personnes que je souhaiterai rajouter au sein de l’équipe URePSSS, à savoir mes coachs sportifs (c’est plus adapté que kiné je pense), qui sont Gilbert (qui m’a connu dès mes premiers jours et jusqu’à maintenant), Magalie, et Guillaume. Sans vous, je n’aurai absolument pas pu réaliser ce que j’ai fait durant cette thèse. Si mes jambes fonctionnent c’est grâce à vous. Ca a été un vrai travail d’équipe et dans l’ombre afin de me permettre d’être à 100% physiquement et moralement au labo. On a parfois repoussé les limites du corps humain et de la science je pense. Guillaume, merci pour ta gentillesse, et les séances de kiné après le labo, parfois douloureuses certes, mais on a également pu tester l’effet de la levure sur la récupération musculaire. C’était de vrai bon moment de vie, et surtout pleins de rigolades. Au passage, je remercie la sécurité sociale, et le contribuable qui m’ont permis d’avoir plus de 5000 séances de kiné à mon actif. Par conséquent, avec ce que je viens d’écrire à l’instant, j’ai beaucoup rigolé pendant 26 ans. Je remercie mes amis et professeurs musiciens, Jérémy, Mathias, Jean-Luc. La musique a toujours été une bouffée d’oxygène pour moi. Merci d’avoir contribué à mon épanouissement. La percu, ça fait maintenant 15 ans que je connais, j’en ferai toujours, peu importe l’endroit ou le moment. Je remercie Lucette, une dame au grand cœur, qui a lancé le Téléthon dans ma ville quand j’étais tout petit et qui m’a nommé mascotte (rôle difficile, car j’étais tout timide. Oui, on a du mal à y croire). C’est extraordinaire de sentir autant soutenu, merci pour tout. Je remercie également, le relais handicap de Lille 1 à savoir Chafika et Sylvie, qui m’ont suivi dès mon arrivée à la fac il y a 8 ans. Du chemin a été parcouru depuis. Je remercie aussi ma famille d’avoir été là, à savoir ma sœur Nathalie, mon beau-frère Tony (oui promis, il y aura un pot à la fin de cette thèse), les nièces et le neveu, Lou-Anne, Hélèna et Yanis. Enfin, celle que je placerai tout en haut de la liste des remerciements, c’est-à-dire ma maman. Je te remercie de m’avoir toujours soutenu, dans mon travail, et mes décisions. On a connu la galère, maintenant cette thèse est un peu une revanche sur la vie. Tu m’as appris à me battre et à ne jamais baisser les bras. Je me rappelle des vieilles VHS de Rocky (la référence) quand j’étais tout petit. Cette thèse, cette vie ça a été un combat de chaque instant, néanmoins tu m’as toujours montré qu’il fallait garder le sourire, et que le meilleur était à venir. On y est. Merci pour tout.. © 2016 Tous droits réservés.. lilliad.univ-lille.fr.
(8) Thèse de Matthias Lambert, Lille 1, 2016. Résumé L’unité contractile du muscle squelettique, le sarcomère, est douée d’une remarquable organisation, à la fois précise et régulière, essentielle à la genèse d’une force. Le maintien de cette structure « semi-cristalline » est régulé par de nombreuses interactions protéine-protéine au sein d’un réseau complexe et dynamique : le cytosquelette sarcomérique. Cependant, à ce jour, cette régulation reste encore mal comprise. De manière intéressante, de très nombreuses protéines myofibrillaires, sont modifiées par la O-N-acétyl-β-D-glucosaminylation (O-GlcNAcylation), une glycosylation atypique, similaire en certains aspects à la phosphorylation et pouvant interagir avec celle-ci. Comme la phosphorylation, cette glycosylation intervient dans une myriade de processus intracellulaires, dont les interactions protéineprotéine. Cependant, bien que décrite comme un modulateur de l’activité contractile, son rôle au sein de la structuration sarcomérique reste encore inconnu. Lors de cette thèse, le rôle de la O-GlcNAcylation dans la structuration du sarcomère a été étudié dans un modèle cellulaire de cellules C2C12 différentiées en myotubes, s’apparentant à des fibres musculaires squelettiques primitives. Le taux global de O-GlcNAcylation a été modulé par des molécules pharmacologiques afin d’en déterminer la répercussion sur la morphométrie du sarcomère. Nous avons ainsi mis en évidence que la modulation du taux de O-GlcNAcylation est beaucoup plus sensible et dynamique dans la fraction myofilamentaire comparativement au protéome total des myotubes C2C12, et que les paramètres morphométriques du sarcomère (largeur des bandes claires et sombres, longueur de la ligne M et du sarcomère) variaient en fonction du taux global de O-GlcNAc myofilamentaire. Nous avons également montré que des complexes protéiques étaient modulés suite à la variation du taux global de O-GlcNAc. Les protéines constitutives de ces complexes différentiels ont été identifiées, notamment des protéines structurales clés pouvant être impliquées dans ces remaniements structuraux : la desmine, l’αB-cristalline, l’α-actinine, la filamine C, et la moésine. Certaines d’entre elles voyaient leur taux de O-GlcNAc et/ou leur expression modulés au sein de ces complexes, suggérant une altération de certaines interactions protéine-protéine. Enfin, nous avons mis en évidence que l’interaction entre la desmine et son chaperon moléculaire, l’αB-cristalline était modulée suite à la variation du taux de OGlcNAc. Afin de mieux comprendre l’implication de la O-GlcNAcylation et de la phosphorylation dans la modulation de l’interaction entre la desmine et l’αB-cristalline, nous avons modulé le taux de phosphorylation de la desmine par l’utilisation d’activateurs spécifiques de certaines protéines kinases. Nos résultats préliminaires montrent que le taux de O-GlcNAcylation et certaines propriétés de la desmine se sont avérés être modulés, associés à l’altération de son interaction avec l’αB-cristalline, suggérant qu’elle puisse être régulée de manière complexe et dynamique par le dialogue entre ces deux modifications. Enfin, nous avons identifié de nouvelles protéines structurales O-GlcNAcylées, telle que la titine, mettant en évidence que la O-GlcNAcylation pourrait jouer un rôle prépondérant dans la structuration du sarcomère. Nous avons également localisé certains sites de O-GlcNAcylation sur des protéines d’intérêt ; en particulier, un site a été identifié au niveau de la desmine, connu comme étant muté dans les desminopathies. L’ensemble de ces résultats suggèrent que la O-GlcNAcylation est impliquée dans la régulation de la structuration du sarcomère, notamment par la modulation d’interactions protéine-protéine. Ces travaux fournissent également de nouvelles données pour la compréhension des mécanismes sous-jacents de certaines pathologies musculaires telles que les myopathies myofibrillaires, caractérisées par une déstructuration marquée de l’organisation sarcomérique.. © 2016 Tous droits réservés.. lilliad.univ-lille.fr.
(9) Thèse de Matthias Lambert, Lille 1, 2016. Abstract The sarcomere is the functional unit of skeletal muscle, presenting an accurate striated organization essential for force generation. The maintenance of this paracrystalline structure is the result of many protein-protein interactions forming a dynamic and intricate network, named the sarcomeric cytoskeleton. However, to date, its regulation is still unclear, while its dysregulation is associated with some cases of muscle atrophy and many muscular diseases. Interestingly, it has been shown that many myofilament proteins are modified by the O-N-acetyl-β-D-glucosaminylation (O-GlcNAcylation), an atypical glycosylation that is similar in some aspects to phosphorylation and may exhibit a dynamic interplay with it. Akin to phosphorylation, O-GlcNAcylation is involved in numerous intracellular processes, including proteinprotein interactions. Although it has been described as a modulator of contractile activity, the role of this glycosylation in sarcomeric organization remains to be considered. In my Ph.D, the role of O-GlcNAcylation in the sarcomeric structure has been studied within C2C12 cells differentiated in myotubes, a precursor cell system of skeletal muscle fibers. The global O-GlcNAc level has been modulated by pharmacological molecules to determine the effects on the sarcomeric morphometry. Firstly, we demonstrated that the modulation of the O-GlcNAc level was more sensitive and dynamic in the myofilament-enriched fraction than the total proteome of the C2C12 myotubes. Secondly, we pointed out that some sarcomeric morphometry parameters (dark band, I band, and the M line widths, as well as the sarcomere length) varied according to the global O-GlcNAc level of the myofilament. We have also shown that some protein complexes were modulated through the variation of the OGlcNAc level. Some proteins within these differential complexes have been identified, including key structural proteins may be involved in these structural changes : desmin, αB-crystallin, α-actinin, filamin-C, and moesin. Some of them displayed a modulation of its O-GlcNAc level and/or expression within these differential complexes, suggesting a disruption of some protein-protein interactions. In this regard, we have highlighted that the interaction between desmin and its molecular chaperone, the αB-crystallin, was modulated through the variation of O-GlcNAc level. In a quest to better understand the involvement of the O-GlcNAcylation and phosphorylation in the interaction between desmin and αB-crystallin, we modulated the phosphorylation rate of by using specific activators of some kinases. Our preliminary results indicated that the O-GlcNAc level and some properties of the desmin have been modulated, associated to the disruption of its interaction with αB-crystallin, suggesting that it may be regulated by an extensive crosstalk and interplay between these posttranslational modifications. Finally, we have identified new O-GlcNAc-modified proteins, such as titin, suggesting that O-GlcNAc may play a meaningful role in the sarcomeric structuration. We have also located some O-GlcNAc sites on proteins of interest ; in particular, an O-GlcNAc site has been identified on desmin, known as a mutated site in desminopathy. Taken together, these Ph.D results suggested that O-GlcNAcylation is involved in sarcomeric structure, protein-protein interactions and in sarcomere remodeling. This work may provide new insights in the understanding of molecular mechanisms of diseases such as myofibrillar myopathies where sarcomere is disorganized, and many protein-protein interactions are disrupted.. © 2016 Tous droits réservés.. lilliad.univ-lille.fr.
(10) Thèse de Matthias Lambert, Lille 1, 2016. Travaux scientifiques. © 2016 Tous droits réservés.. lilliad.univ-lille.fr.
(11) Thèse de Matthias Lambert, Lille 1, 2016. Publications scientifiques Deracinois B, Camoin L, Lambert M, Audebert S, Dupont E, Bastide B, Cieniewski-Bernard C. (2016 – en cours de rédaction) O-GlcNAcylation site mapping by (azide-alkyne) click chemistry and mass spectrometry following intensive fractionation. Molecular and Cellular Proteomics Lambert M, Richard E, Duban-Deweer S, Krzewinski F, Deracinois B, Dupont E, Bastide B, CieniewskiBernard C. (2016) O-GlcNAcylation is a key modulator of skeletal muscle sarcomeric morphometry associated to modulation of protein-protein interactions. Biochim Biophys Acta. 1860(9):2017-2030 Cieniewski-Bernard C, Lambert M, Dupont E, Montel V, Stevens L, Bastide B. (2014) O-GlcNAcylation, contractile protein modifications and calcium affinity in skeletal muscle. Frontiers in physiology. 30;5:421 Cieniewski-Bernard C, Dupont E, Deracinois B, Lambert M, Bastide B. (2014) Multiplexed detection of O-GlcNAcome, phosphoproteome, and whole proteome within the same gel. Frontiers in endocrinology. Oct 28;5:184. Communications orales Septembre 2016 : 45th European Muscle Conference, Montpellier, France. Lambert M, Deracinois B, Camoin L, Audebert S, Girard A, Bastide B, Cieniewski-Bernard C. O-GlcNAcylation is a key modulator of cytoskeletal interactome involved in the skeletal muscle sarcomeric organization. Septembre 2015 : 15ème journée André Verbert – Congrès des doctorants, Lille, France. Lambert M, Richard E, Duban-Deweer S, Krzewinski F, Deracinois B, Dupont E, Bastide B, Cieniewski-Bernard C. O-GlcNAcylation in skeletal muscle: a key regulator of the sarcomeric structure? Septembre 2014 : 43th European Muscle Conference, Salzbourg, Autriche. Lambert M, Richard E, Duban-Deweer S, Krzewinski F, Deracinois B, Dupont E, Bastide B, Cieniewski-Bernard C. OGlcNAcylation modulates sarcomeric organization and protein-protein interactions in C2C12 myotubes. Résumé soumis à publication : Kawai M, Lamb G, Galler S. Editorial on EMC 2014 special issue. J Muscle Res Cell Motil. (2015). 36(1):1-3.. Communications écrites Juillet 2016 : New directions in biology and disease of skeletal muscle conference, Orlando, Floride, Etats-Unis. Lambert M, Richard E, Duban-Deweer S, Krzewinski F, Deracinois B, Dupont E, Bastide B, Cieniewski-Bernard C. O-GlcNAcylation in skeletal muscle: a key regulator of the sarcomeric structure? Mars 2016 : 5th international congress of myology, Lyon, France. Lambert M, Richard E, DubanDeweer S, Krzewinski F, Deracinois B, Dupont E, Bastide B, Cieniewski-Bernard C. O-GlcNAcylation in skeletal muscle: a key regulator of the sarcomeric structure? Septembre 2014 : 43th European Muscle Conference, Salzbourg, Autriche. Deracinois B, Dupont B, Richard E, Lambert M, Bastide B, Cieniewski-Bernard C. Phosphorylation/O-GlcNAcylation interplay in the modulation of protein synthesis: focus on the IGF-1 pathway in differentiated C2C12 myotubes. Résumé soumis à publication : Kawai M, Lamb G, Galler S. Editorial on EMC 2014 special issue. J Muscle Res Cell Motil. (2015). 36(1):1-3. Octobre 2013 : EuPA 2013 Scientific Meeting, Saint Malo, France. Deracinois B, Dupont E, DubanDeweer S, Camoin L, Richard E, Lambert M, Bastide B, Cieniewski-Bernard C. Study of the phosphorylation/O-GlcNAcylation interplay in differentiated C2C12 myotubes: focus on the PI3K/AKT/mTORpathway.. © 2016 Tous droits réservés.. lilliad.univ-lille.fr.
(12) Thèse de Matthias Lambert, Lille 1, 2016. Index et abréviations. © 2016 Tous droits réservés.. lilliad.univ-lille.fr.
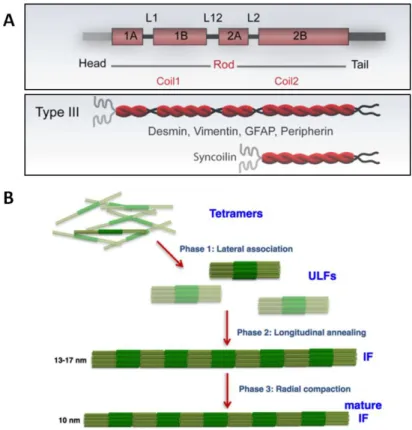
(13) Thèse de Matthias Lambert, Lille 1, 2016. Index des figures Figure 1 :. Présentation succincte des différents types de muscle dans l’organisme.. Figure 2 :. Organisation du muscle strié squelettique – Du muscle, jusqu’à son unité contractile, le sarcomère.. Figure 3 :. Coupe transversale d’un muscle squelettique après coloration à l’hématoxyline-éosine (microscopie en champ clair, x400, image C.Picken).. Figure 4 :. Coupe transversale d’un muscle squelettique après précipitation du produit de la réaction ATPase des chaines lourdes de myosines, suite à la pré-incubation des fibres musculaires dans un milieu acide à pH 4,3, et à leur coloration.. Figure 5 :. Anatomie fonctionnelle d’une fibre musculaire.. Figure 6 :. Striation et organisation du sarcomère.. Figure 7 :. Organisation et structure des filaments épais de myosine.. Figure 8 :. Structure des filaments fins d’actine.. Figure 9 :. Organisation du complexe des troponines sur la molécule de tropomyosine.. Figure 10 :. Les différentes étapes moléculaires de l’interaction entre l’actine et la myosine, menant à la contraction et au raccourcissement du sarcomère.. Figure 11 :. Théorie de glissement des filaments fins d’actine le long des myofilaments épais de myosine, synonyme de contraction.. Figure 12 :. Liste non exhaustive et localisation des protéines appartenant au cytosquelette sarcomérique.. Figure 13 :. Schématisation de l’organisation stricte et précise du sarcomère à l’origine de sa stabilité au repos, pendant la contraction, et lors du relâchement.. Figure 14 :. Localisation des protéines géantes au sein du sarcomère.. Figure 15 :. Structure de la titine qui s’étend sur la moitié d’un sarcomère, de la strie Z à la ligne M.. Figure 16 :. Localisation des domaines d’interaction de la titine avec ses protéines partenaires à la strie Z (A), à la bande I (B), à la bande A et à la ligne M (C).. Figure 17 :. Structure de l’obscurine et localisation des domaines d’interactions avec ses protéines partenaires.. Figure 18 :. Structure de la nébuline et localisation des domaines d’interaction avec ses protéines partenaires.. Figure 19 :. Pontage des filaments fins d’actine au niveau de la strie Z par l’alpha-actinine.. Figure 20 :. Structure de l’homodimère d’alpha-actinine.. Figure 21 :. Interactome de l’alpha-actinine.. Figure 22 :. Modèle d’organisation du cytosquelette sarcomérique au niveau de la strie Z, afin d’y assurer le maintien de son intégrité, son organisation, et la communication physique avec le sarcolemme.. Figure 23 :. Structure et assemblage de la desmine.. © 2016 Tous droits réservés.. lilliad.univ-lille.fr.
(14) Thèse de Matthias Lambert, Lille 1, 2016. Figure 24 :. Schématisation du réseau de desmine au sein de la fibre musculaire et de ses différentes interactions avec des protéines des myofibrilles, des mitochondries, du noyau et du sarcolemme.. Figure 25 :. La desmine au cœur d’un vaste interactome.. Figure 26 :. Organisation de la ligne M.. Figure 27 :. La myomésine, le composant principal des ponts M.. Figure 28 :. Interactome de la strie Z et de la ligne M, points nodaux du sarcomère et carrefours de différents mécanismes et voies intracellulaires.. Figure 29 :. Schématisation du système de dégradation protéique ubiquitine-protéasome (UPS).. Figure 30 :. Implication des protéines chaperonnes dans l’homéostasie protéique.. Figure 31 :. Organisation des différentes protéines au sein de la ligne M du sarcomère.. Figure 32 :. Etiologie de pathologies liées à une désorganisation du sarcomère : les liens de causes à effets.. Figure 33 :. Phénotype cellulaire du muscle strié squelettique dans le cas d’une desminopathie.. Figure 34 :. Les modifications post-traductionnelles, un mécanisme de complexification du génome et du protéome offrant une diversité fonctionnelle accrue à la cellule.. Figure 35 :. Mécanisme de la O-GlcNAcylation.. Figure 36 :. La voie de biosynthèse des hexosamines menant à la synthèse de l’UDP-GlcNAc est nécessaire à la O-GlcNAcylation.. Figure 37 :. Structure de l’OGT.. Figure 38 :. Structure de l’OGA.. Figure 39 :. Les différentes possibilités de dialogue entre la phosphorylation et la O-GlcNAcylation au sein d’une même protéine.. Figure 40 :. Principales fonctions cellulaires régulées par la O-GlcNAcylation.. Figure 41 :. La O-GlcNAcylation et son impact sur l’homéostasie physiologique et le développement de pathologies.. Figure 42 :. Cartographie des protéines pouvant être O-GlcNAcylées au niveau du sarcomère.. Figure 43 :. Localisation de l’OGT au l’OGA au niveau du sarcomère.. Figure 44 :. Effets de la O-GlcNAcylation sur les paramètres d’activation calcique des fibres striées squelettiques.. Figure 45 :. Localisation des enzymes régulatrices de la phosphorylation de la MLC2 et régulant la OGlcNAcylation au sein du sarcomère.. Figure 46 :. Mécanismes sous-jacents des effets d’une augmentation de la O-GlcNAcylation par le Thiamet-G et la dexaméthasone menant à l’atrophie des myotubes de type C2C12.. Figure 47 :. Schématisation de la culture cellulaire de type C2C12.. Figure 48 :. Schématisation simplifiée de la myogenèse du muscle strié squelettique.. Figure 49 :. Cinétique d’expression des principales protéines contractiles et structurales.. © 2016 Tous droits réservés.. lilliad.univ-lille.fr.
(15) Thèse de Matthias Lambert, Lille 1, 2016. Figure 50 :. Cinétique de formation des sarcomères par marquage immunofluorescent des chaines lourdes de myosines (MHC).. Figure 51 :. Cinétique de formation des myotubes par une coloration au May-Grünwald Giemsa.. Figure 52 :. Effet des traitements sur l’état de maturation des myotubes.. Figure 53 :. Mesure de la cytotoxicité des traitements sur les myotubes via le test MTT.. Figure 54 :. Mesures préliminaires du taux global de O-GlcNAcylation de la fraction myo-filamentaire.. Figure 55 :. Chromatogrammes représentatifs de la séparation de l’UDP-GlcNAc et de l’UDP-GalNAc par HPLC sur colonne échangeuse d’anions.. Figure 56 :. Mesure du taux d’UDP-GlcNAc par HPLC.. Figure 57 :. Quantification de l’expression de la GFAT au sein de l’extrait protéique total par western blot.. Figure 58 :. Dosage de l’activité enzymatique de l’OGA par hydrolyse du pNP-GlcNAc.. Figure 59 :. Quantification de l’expression de la O-GlcNAcase (OGA) au sein de l’extrait protéique total par western blot.. Figure 60 :. Quantification de l’expression de la O-GlcNAc Transférase (OGT) au sein de l’extrait protéique total par western blot.. Figure 61 :. Quantification par western blot du taux global de O-GlcNAcylation au sein de l’extrait protéique total et dans la fraction myofilamentaire.. Figure 62 :. Spécificité du marquage des chaines lourdes de myosines par l’anticorps My32.. Figure 63 :. Visualisation par microscopie inversée à fluorescence de l’optimisation du marquage immunofluorescent des chaines lourdes de myosines.. Figure 64 :. Modulation de la morphométrie du sarcomère en fonction des variations du taux global de OGlcNAcylation mesurées au sein de la fraction myofilamentaire des myotubes C2C12.. Figure 65 :. Démarche scientifique dans le cadre de la mise en évidence de l’implication de la OGlcNAcylation dans les interactions protéine-protéine suite à la modulation du taux global de OGlcNAcylation des myotubes C2C12.. Figure 66 :. Comparaison entre une électrophorèse en conditions dénaturantes de type SDS-PAGE, et une électrophorèse en conditions natives de type Blue-Native PAGE.. Figure 67 :. Principe de la 2D-BN-PAGE/SDS-PAGE.. Figure 68 :. Résultat de l’optimisation du tampon d’extraction protéique, et de la composition du gel de polyacrylamide en vue d’une séparation électrophorétique en conditions non dénaturantes.. Figure 69 :. Comparaison de la séparation de l’extrait protéique total et de la fraction myofilamentaire par Blue-Native PAGE.. Figure 70 :. Amélioration de la solubilisation des protéines de la fraction myofilamentaire par l’utilisation préalable de différents détergents en vue d’une électrophorèse en conditions non dénaturantes.. Figure 71 :. Préservation des interactions protéine-protéine lors de l’extraction différentielle de la fraction myofilamentaire : le choix du pontage chimique.. Figure 72 :. Comparaison des différents profils protéiques de la fraction myofilamentaire et totale ayant subi ou non un pontage chimique des protéines.. © 2016 Tous droits réservés.. lilliad.univ-lille.fr.
(16) Thèse de Matthias Lambert, Lille 1, 2016. Figure 73 :. Comparaison entre la Blue-Native PAGE et la Red-Native PAGE.. Figure 74 :. Modulation de certains complexes protéiques déterminée par la Red-Native PAGE.. Figure 75 :. Diagramme représentant la proportion des différents types de protéines identifiées par spectrométrie de masse de type Nano-LC MS/MS, au sein des complexes différentiels mis en évidence par la Red-Native PAGE.. Figure 76 :. Les protéines « candidates » appartiennent aux complexes différentiels dans lesquels elles ont été identifiées, suite à la variation du taux global de O-GlcNAcylation.. Figure 77 :. Quantification par western blot du taux global de O-GlcNAcylation des complexes protéiques différentiels.. Figure 78 :. Quantification du taux de O-GlcNAcylation immunoprécipitation.. Figure 79 :. Les interactions protéine-protéine de certains candidats sont modulées suite à la variation du taux global de O-GlcNAcylation.. Figure 80 :. Les interactions protéine-protéine entre la desmine et l’alpha-B cristalline sont modulées suite à la variation du taux global de O-GlcNAcylation.. Figure 81 :. Schéma des différentes techniques d’enrichissement des protéines/peptides O-GlcNAcylés afin de les identifier et de localiser leurs sites de O-GlcNAcylation par spectrométrie de masse.. Figure 82 :. Marquage métabolique in cellulo des protéines par la GlcNAz.. Figure 83 :. Comparaison des profils protéiques de la fraction myofilamentaire précipitée de manière séquentielle par différents pourcentages de sulfate d’ammonium.. Figure 84 :. Diagramme des différents types de protéines O-GlcNAcylées identifiées par spectrométrie de masse suite à l’enrichissement par marquage chimio-enzymatique in vitro, suivi de la clickchemistry, à partir de 4mg échantillon protéique total de myotubes matures de type C2C12.. Figure 85 :. Méthodologie de localisation des sites O-GlcNAcylés après identification des peptides bétaéliminés : l’exemple de BAG3.. Figure 86 :. Expression totale de la desmine.. Figure 87 :. Effets du TPA et du Thiamet-G sur la modulation des différentes formes phosphorylées de la desmine.. Figure 88 :. Effets du TPA et du Thiamet-G sur la phosphorylation globale de la desmine.. Figure 89 :. Effets du TPA et du Thiamet-G sur le taux de O-GlcNAcylation de la desmine.. Figure 90 :. Effets du TPA et du Thiamet-G sur les propriétés lectiniques de la desmine.. Figure 91 :. Effets du TPA et Thiamet-G sur la modulation de l’interaction entre la desmine et l’αB-cristalline.. Figure 92 :. Visualisation d’une relocalisation de l’OGT et l’OGA au sein du sarcomère du muscle soléaire du rat, avant et après une période d’hypodynamie-hypokinésie.. Figure 93 :. Représentation schématique de la modulation de l’organisation du sarcomère suite à la variation du taux global de O-GlcNAcylation de la fraction myofilamentaire.. Figure 94 :. Schéma des effets d’une non-utilisation du muscle sur la structure du sarcomère et sur ses propriétés fonctionnelles.. Figure 95 :. Résultat préliminaire du marquage immunofluorescent de l’α-actinine des myotubes C2C12.. © 2016 Tous droits réservés.. de certaines protéines candidates par. lilliad.univ-lille.fr.
(17) Thèse de Matthias Lambert, Lille 1, 2016. Figure 96 :. Relation longueur du sarcomère et tension développée.. Figure 97 :. Régulation potentiellement complexe de la O-GlcNAcylation et de la phosphorylation de la desmine.. Figure 98 :. Interactome simplifié des « protéines candidates » qui pourraient expliquer les changements d’organisation du sarcomère suite à des variations de O-GlcNAcylation dans les myotubes C2C12.. Figure 99 :. Schéma de l’influence de l’αB-cristalline sur les filaments de desmine.. Figure 100 : Modèle d’interaction proposé entre les filamines et les filaments d’actine. Figure 101 : Mise à jour de la cartographie des protéines identifiées comme pouvant être O-GlcNAcylées au niveau du sarcomère. Figure 102 : Mutations de la desmine et physiopathologie des desminopathies. Figure 103 : Localisation du domaine Ig-like 123 de la titine. Figure 104 : Interactome de la desmine. Figure 105 : Visualisation par microscopie inversée de coupes longitudinales de muscle soléaire de rat marquées par immunofluorescence au niveau de l’α-actinine et de la desmine. Figure 106 : Diagramme montrant les différents effets et implications de la O-GlcNAcylation dans le muscle squelettique.. © 2016 Tous droits réservés.. lilliad.univ-lille.fr.
(18) Thèse de Matthias Lambert, Lille 1, 2016. Index des tableaux Tableau 1 :. Principales caractéristiques structurales, fonctionnelles et métaboliques des fibres musculaires squelettiques.. Tableau 2 :. Paramètres de mise au point des traitements pharmacologiques afin de moduler le taux de OGlcNAcylation des myotubes C2C12.. Tableau 3 :. Sélection des traitements pharmacologiques suite aux premières mises au point.. Tableau 4 :. Paramètres d’optimisation du marquage immunofluorescent des chaines lourdes de myosines.. Tableau 5 :. Tableau récapitulatif de la morphométrie du sarcomère en fonction des traitements et du taux global de O-GlcNAcylation myofilamentaire.. Tableau 6 :. Liste des différentes optimisations concernant l’électrophorèse en conditions natives sur l’échantillon protéique total et la fraction myofilamentaire. Tableau 7 : Optimisation du gel de séparation de polyacrylamide et du tampon lyse cellulaire en vue d’une électrophorèse en conditions non dénaturantes.. Tableau 8 :. Liste restreinte des protéines identifiées par spectrométrie de masse de type Nano-LC MS/MS, au sein des complexes différentiels modulés par le taux global de O-GlcNAcylation et déterminés par la Red-Native PAGE : protéines candidates pouvant expliquer la modulation de la structure du sarcomère liée aux variations du taux global de O-GlcNAcylation.. Tableau 9 :. Liste restreinte des protéines identifiées comme étant O-GlcNAcylées et dont certains de leurs sites de O-GlcNAcylation ont été localisés : les protéines d’intérêt du maintien de l’organisation sarcomérique.. Tableau 10 : Tableau récapitulatif des effets du TPA et du Thiamet-G sur la modulation de la desmine.. © 2016 Tous droits réservés.. lilliad.univ-lille.fr.
(19) Thèse de Matthias Lambert, Lille 1, 2016. Abréviations ABD : Actin binding actin (domaine de liaison à l’actine) Ac4-GlcNAz : N-azido-acétyl-glucosamine peracétylée ACN : Acétronitrile ADN : Acide désoxyribonucléique ADP : Adénosine diphosphate AMPK : AMP-activated protein kinase APS : Ammonium persulfate ARN : Acide ribonucléique ATP : Adénosine triphosphate BAG3 : BCL2 Associated Athanogene 3 BSA : Bovine serum Albumine CAMD : Calmodulin binding domain (domaine de liaison à la calmoduline) CaMKII : Calcium/calmodulin-dependent protein kinase II CK : Creatine Kinase DAPI : 4',6-diamidino-2-phenylindole DMSO : Diméthylsulfoxide DON : 6-Diazo-Oxo-L-Norleucine DTT : Dithiothreitol EDL : extensor digitorum longus Fn : Fibronectin GFAT: glutamine fructose-6-phosphate amidotransférase GalNAc : N-acétyl-D-galactosamine GlcNAc : N-acétyl-D-glucosamine HPLC : « high performance liquid chromatography » (chromatographie liquide haute performance) HRP-WGA: « horse radish peroxidase » (WGA couplée à la peroxydase) HSP : beat shock protein ; protéine de choc thermique IEF : isoélectrofocalisation Ig : Immunoglobuline MALDI-TOF MS : « matrix-assisted laser desorptionlionisation-time of flight mass spectrometry » (spectrométrie de masse par désorption/ionisation par tir laser assisté par matrice et par mesure du temps de vol) MARP : Muscle ankyrin repeat protein MHC : myosin heavy chain (chaîne lourde de myosine) MLC : myosin light chain (chaîne légère de myosine) MS/MS : spectrométrie de masse en tandem MTT : méthylthiazoletétrazolium MuRF : Muscle-specific RING finger O-GlcNAc : 0-N-acétyl-glucosaminylation OGA = O-GicNAcase : N-acétyl-f3-D-glucosaminidase OGT = O-GlcNAc transférase: uridine diphospho-N-acétyl-glucosaminyl transférase PBS : Phosphate Buffer Saline PFA : paraformaldéhyde PKA : Protéine Kinase A PKC : Protéine Kinase C PLA : Proximity Ligation Assay PNGase F : peptide N-glycosidase F PNP : para-nitrophénol PNP-GlcNAc : para-nitrophényl-N-acétylglucosamine PTM : Modification post-traductionnelle PUGNAc : 0-(2-acétamido-2-déoxy-D-glucopyranosilidène )amino-N-phénylcarbamate RL-2 : anticorps anti-0-GlcNAc. © 2016 Tous droits réservés.. lilliad.univ-lille.fr.
(20) Thèse de Matthias Lambert, Lille 1, 2016. Rpm : rotation par minute SDS-PAGE: «sodium dodécyl sulfate - polyacrylamide gel électrophoresis » (électrophorèse en gel de polyacrylamide en présence de sodium dodécyl sulfate) SR : Spectrin repeat TBST : Tris buffer saline - Tween 20 TEMED : Tetramethylethylenediamine TFA : Acide trifluoroacétique Tm : tropomyosine Tn : troponine TPA : 12-O-tétradécanylphobol-13-acétate TPR: tetratricopeptide repeat UDP-GlcNAc : uridine diphospho-N-acétylglucosamine WGA : wheat germ agglutinin (lectine isolée du germe de blé) ZASP : Z-band alternatively spliced PDZ-motif protein 2D : électrophorèse bidimensionnelle. © 2016 Tous droits réservés.. lilliad.univ-lille.fr.
(21) Thèse de Matthias Lambert, Lille 1, 2016. Sommaire. © 2016 Tous droits réservés.. lilliad.univ-lille.fr.
(22) Thèse de Matthias Lambert, Lille 1, 2016. SOMMAIRE INTRODUCTION Partie 1 : L’organisation du muscle squelettique et la structure de son sarcomère I.. Préambule ...................................................................................................................................... 1. II.. L’organisation générale du muscle squelettique .............................................................................. 3 1.. Du muscle dans son ensemble jusqu’aux fibres musculaires................................................................ 4 A.. Les caractéristiques des différentes fibres musculaires .................................................................... 4. B.. L’anatomie des fibres musculaires .................................................................................................... 6. 2.. L’unité contractile du muscle strié squelettique : le sarcomère ........................................................... 7 A.. Son organisation ................................................................................................................................ 7. B.. La structure des myofilaments .......................................................................................................... 8. C.. Mécanismes moléculaires de la contraction ................................................................................... 11. III. La structure du sarcomère : un cytosquelette atypique .................................................................. 14 1.. Notion de cytosquelette sarcomérique ............................................................................................... 14. 2.. Les protéines géantes du sarcomère ................................................................................................... 16 A.. La titine : le squelette du sarcomère ............................................................................................... 17. B.. L’obscurine : le lien existant entre le sarcomère et le réticulum sarcoplasmique .......................... 19. C.. La nébuline : l’empreinte des filaments fins d’actine...................................................................... 20. 3.. Les protéines structurales de la strie Z ................................................................................................ 20 A.. L’alpha-actinine ............................................................................................................................... 21. B.. La téléthonine (ou Tcap) .................................................................................................................. 22. C.. La myotiline ..................................................................................................................................... 23. D.. La myopalladine............................................................................................................................... 23. E.. La filamine C .................................................................................................................................... 23. F.. La protéine MLP (Muscle LIM Protein) ............................................................................................ 24. G.. ZASP (Z-band Alternatively Spliced PDZ-motif protein) .................................................................. 24. 4.. Les filaments intermédiaires ............................................................................................................... 25 A.. La desmine : filament intermédiaire majoritaire de la fibre musculaire mature ............................ 26. B.. Interactome de la desmine.............................................................................................................. 28. C.. Fonctions structurales et mécaniques de la desmine ..................................................................... 30. 5.. La structure de la ligne M .................................................................................................................... 31 A.. La ligne M au service de la stabilité du sarcomère.......................................................................... 32. B.. La myomésine : acteur majeur de l’intégrité de la ligne M ............................................................. 32. IV. Le cytosquelette sarcomérique : une plateforme dynamique pour le sarcomère et la cellule .......... 34. © 2016 Tous droits réservés.. lilliad.univ-lille.fr.
(23) Thèse de Matthias Lambert, Lille 1, 2016. 1.. Le maintien du sarcomère passe par la dégradation locale de ses constituants ................................ 35. 2.. Les protéines chaperonnes : le service assistance du sarcomère ....................................................... 36. 3. Des protéines du métabolisme ancrées au sarcomère sous forme de métabolon : production d’une source locale d’énergie indispensable ........................................................................................................ 38 4. V.. Le sarcomère : carrefour pour de nombreuses voies de signalisation ............................................... 40 Quand la structure du sarcomère est mise à mal… ......................................................................... 42. Partie 2 : La O-N-acétyl-β-D-glucosaminylation I.. Avant-propos : Intérêt de l’étude des modifications post-traductionnelles ..................................... 45. II.. Une modification post-traductionnelle à part entière : la O-GlcNAcylation ..................................... 47 1.. Une glycosylation atypique ................................................................................................................. 47. 2.. La O-GlcNAcylation est étroitement liée à la voie de biosynthèse des hexosamines ......................... 48 A.. L’UDP-GlcNAc : le produit final de la voie de biosynthèse des hexosamines ................................. 48. B.. LA GFAT : l’enzyme limitante de la voie de biosynthèse des hexosamines .................................... 50. 3.. Les enzymes régulatrices de la O-GlcNAcylation................................................................................. 51 A.. La O-GlcNAc Transférase (ou OGT).................................................................................................. 51. B.. La O-GlcNAcase (ou OGA) ................................................................................................................ 55. III. La balance O-GlcNAcylation/phosphorylation : un mécanisme complexe ....................................... 56 IV. La découverte des fonctions attribuées à la O-GlcNAcylation ......................................................... 60 V.. Les dérégulations de la O-GlcNAcylation et son implication dans certaines pathologies .................. 62. VI. Implications de la O-GlcNAcylation dans la physiologie du muscle squelettique ............................. 63 1.. La O-GlcNAcylation : modulateur de l’activité contractile .................................................................. 65 A.. Rôle de la O-GlcNAcylation dans la sensibilité calcique .................................................................. 65. B. La MLC2 candidate pour expliquer l’implication de la O-GlcNAcylation dans la modulation de la sensibilité calcique................................................................................................................................... 67 2.. L’exercice musculaire module la O-GlcNAcylation du muscle squelettique ....................................... 70. 3.. La O-GlcNAcylation et les dysfonctions musculaires ........................................................................... 71 A.. La O-GlcNAcylation est associée aux phénomènes d’atrophie musculaire .................................... 71. B.. O-GlcNAcylation et pathologies du muscle strié squelettique ........................................................ 74. PROBLEMATIQUE………………………………………………………………………………………… ………72. © 2016 Tous droits réservés.. lilliad.univ-lille.fr.
(24) Thèse de Matthias Lambert, Lille 1, 2016. MATERIEL ET METHODES I.. Culture cellulaire de type C2C12 .................................................................................................... 78 1.. Prolifération des myoblastes et différenciation des myotubes .......................................................... 78. 2.. Traitements pharmacologiques ........................................................................................................... 78 A.. Modulation des taux de O-GlcNAcylation ....................................................................................... 78. B.. Modulation du taux de phosphorylation de la desmine ................................................................. 79. 3. II.. Test de viabilité cellulaire au méthylthiazoletétrazolium (MTT)......................................................... 79 Marquage par immunofluorescence du sarcomère ........................................................................ 80. 1.. Détermination des paramètres morphométriques ............................................................................. 80. 2.. Index de fusion .................................................................................................................................... 80. III. Extractions protéiques .................................................................................................................. 81 1.. Extraction du protéome total .............................................................................................................. 81. 2.. Extraction fractionnée du protéome ................................................................................................... 81. 3.. Extraction du protéome en condition non dénaturante (condition native) ....................................... 81. IV. Dosage de l’activité enzymatique de la O-GlcNAcase (OGA) ........................................................... 81 V.. Dosage du contenu en UDP-GlcNAc de la cellule ............................................................................ 82. VI. Techniques d’enrichissement des protéines : immunoprécipitation, co-immunoprécipitation et chromatographie d’affinité .................................................................................................................. 82 1.. Expériences d’immunoprécipitation des protéines O-GlcNAcylées .................................................... 82. 2.. Détermination des propriétés lectiniques de certaines protéines par chromatographie d’affinité ... 83. 3.. Enrichissement du protéome phosphorylé ......................................................................................... 83. VII. Expériences de co-immunoprécipitation en vue de l’analyse des interactions protéine-protéine..... 84 VIII. Les différentes techniques d’électrophorèse utilisées .................................................................... 84 1.. Electrophorèse en conditions dénaturantes SDS-PAGE et western blot ............................................ 84. 2. Electrophorèse SDS-PAGE en présence de Phostag : séparation des différentes formes phosphorylées ............................................................................................................................................. 85 3. Electrophorèses en conditions non dénaturantes : la Blue Native-PAGE (BN-PAGE) et la Red NativePAGE (RN-PAGE) .......................................................................................................................................... 85 IX. Préparation des échantillons en vue de l’identification des constituants des complexes protéiques par spectrométrie de masse ................................................................................................................. 86 1.. Digestion « en gel » des protéines ...................................................................................................... 86. 2. Analyse par spectrométrie de masse de type nano-LC-MS (nano liquid chromatography – mass spectrometry) .............................................................................................................................................. 86 X.. Identification des sites de O-GlcNAcylation ................................................................................... 87 1.. Fractionnement de l’extrait cytosolique ............................................................................................. 87 A.. © 2016 Tous droits réservés.. Précipitation au sulfate d’ammonium ............................................................................................. 87. lilliad.univ-lille.fr.
(25) Thèse de Matthias Lambert, Lille 1, 2016. B. 2.. IEF en phase liquide ......................................................................................................................... 88 Marquage des protéines O-GlcNAcylées par incorporation de la fonction azide ............................... 88. A.. Stratégie de marquage métabolique in cellulo................................................................................ 88. B.. Stratégie de marquage chimio-enzymatique in vitro ...................................................................... 88. 3.. Etape de déglycosylation ..................................................................................................................... 89 A.. Traitement à la PNGase F de l’extrait marqué in cellulo ................................................................. 89. B.. Traitement à la béta-hexosaminidase de l’extrait marqué in vitro ................................................. 89. 4.. Stratégie d’enrichissement des protéines O-GlcNAcylées par Click-chemistry .................................. 89. 5.. Spectrométrie de masse de type Orbitrap LC-MS/MS ........................................................................ 90. XI. Analyse statistique ....................................................................................................................... 90. RESULTATS PREAMBULE…………………………………………………………………………………………………………………………………………91. Chapitre 1 : Modulation du taux global de O-GlcNAcylation dans les myotubes C2C12 I.. Objectif ........................................................................................................................................ 95. II.. Sélection des traitements pharmacologiques ................................................................................ 96 1.. Effet des traitements sur la maturation générale des myotubes et des sarcomères ......................... 96. 2.. Evaluation de la cytotoxicité des traitements ..................................................................................... 97. 3.. Mesures préliminaires du taux global de O-GlcNAcylation ................................................................. 98. III. Efficacité des traitements pharmacologiques sélectionnés ............................................................. 99 1.. Effet des traitements sur le taux d’UDP-GlcNAc ................................................................................. 99. 2.. Effet des traitements sur l’activité de la O-GlcNAcase (OGA) ........................................................... 102. 3. Effets des traitements pharmacologiques sur la modulation du taux global de O-GlcNAcylation dans l’extrait protéique total et la fraction myofilamentaire des myotubes C2C12 matures ........................... 106 IV. Conclusion .................................................................................................................................. 107. Chapitre 2 : Détermination d'une modulation de la structure du sarcomère suite à la variation du taux global de O-GlcNAcylation myofilamentaire : la morphométrie du sarcomère I.. Objectif ...................................................................................................................................... 108. II.. Optimisation de la technique ...................................................................................................... 108. III. Mesure de la morphométrie en fonction du taux global de O-GlcNAcylation myofilamentaire ...... 110. © 2016 Tous droits réservés.. lilliad.univ-lille.fr.
(26) Thèse de Matthias Lambert, Lille 1, 2016. IV. Conclusion .................................................................................................................................. 112. Chapitre 3 : Mise en évidence de l'implication de la O-GlcNAcylation dans les interactions protéine-protéine I.. Objectif ...................................................................................................................................... 113. II. Identification globale sans a priori des complexes protéiques modulés suite à la variation du taux global de O-GlcNAcylation grâce à l’électrophorèse en conditions non dénaturantes ........................... 114 1.. Pourquoi l’électrophorèse en conditions non dénaturantes ? ......................................................... 114. 2.. Optimisation de la Blue-Native PAGE pour l’échantillon total .......................................................... 116. 3.. Optimisation de la Blue-Native PAGE pour la fraction myofibrillaire ............................................... 119. 4.. Utilisation du Ponceau S, une alternative au Coomassie G-250 : la Red-Native PAGE ..................... 123. III. Mise en évidence de la modulation de certains complexes protéiques de l’extrait total par RedNative PAGE ...................................................................................................................................... 124 IV. Identification des protéines constitutives des complexes modulés mis en évidence par Red-Native PAGE ................................................................................................................................................. 125 V. Modulation des interactions protéine-protéine due aux variations du taux de O-GlcNAcylation : focus sur les protéines “candidates” ................................................................................................... 128 VI. Conclusion .................................................................................................................................. 133. Chapitre 4 : Identification des sites de O-GlcNAcylation des protéines d'intérêt I.. Objectif ...................................................................................................................................... 134. II.. Technique .................................................................................................................................. 134. III. Etude de la fraction myofilamentaire .......................................................................................... 138 IV. Etude de la fraction protéique totale ........................................................................................... 139 V.. Conclusion .................................................................................................................................. 142. Chapitre 5 : Modulation de la phosphorylation de la desmine, et analyse de la balance phosphorylation/O-GlcNAcylation dans l'interaction desmine - αB-cristalline I.. Etude de la modulation de la phosphorylation de la desmine....................................................... 143. II.. Etude de la modulation du taux de O-GlcNAcylation et des propriétés lectiniques de la desmine .. 146. III. Etude de la modulation de l’interaction desmine – αB-cristalline ................................................. 147 IV. Conclusion .................................................................................................................................. 148. © 2016 Tous droits réservés.. lilliad.univ-lille.fr.
Figure




+7





Outline
L’alpha-actinine
Fonctions structurales et mécaniques de la desmine
La balance O-GlcNAcylation/phosphorylation : un mécanisme complexe
Implications de la O-GlcNAcylation dans la physiologie du muscle squelettique
La O-GlcNAcylation est associée aux phénomènes d’atrophie musculaire
Choix du modèle cellulaire, le myotube C2C
Effet des traitements sur le taux d’UDP-GlcNAc
Effet des traitements sur l’activité de la O-GlcNAcase (OGA)
heure à température ambiante ou la nuit à 4°C
Quelles seraient les conséquences fonctionnelles du changement de morphométrie sarcomérique observé ?
Documents relatifs