UNIVERSITE PAUL SABATIER TOULOUSE III
UFR Sciences
THESE DE DOCTORAT
Discipline : Immunologie
Présentée et soutenue par
Nicolas ESPAGNOLLE
Le 28 septembre 2007
En vue de l’obtention du grade de
DOCTEUR DE L’UNIVERSITE TOULOUSE III
Etude des interactions moléculaires à l’aire de
contact formée entre les lymphocytes T et les cellules
présentatrices d’antigène
JURY
Pr. Joost van MEERWIJK
Président
Dr. Jacques NUNES
Rapporteur
Dr. Emmanuel DONNADIEU
Rapporteur
UNIVERSITE PAUL SABATIER TOULOUSE III
UFR Sciences
THESE DE DOCTORAT
Discipline : Immunologie
Présentée et soutenue par
Nicolas ESPAGNOLLE
Le 28 septembre 2007
En vue de l’obtention du grade de
DOCTEUR DE L’UNIVERSITE TOULOUSE III
Etude des interactions moléculaires à l’aire de
contact formée entre les lymphocytes T et les cellules
présentatrices d’antigène
JURY
Pr. Joost van MEERWIJK
Président
Dr. Jacques NUNES
Rapporteur
Dr. Emmanuel DONNADIEU
Rapporteur
A mon Grand Père,
JEAN ESPAGNOLLE (1901-1994)
Je dédie ce travail
Remerciements
Aucamville, le 30 août 2007
Tandis que la nuit tombe en cette journée banale de fin août, je cherche l’inspiration afin de remercier toutes les personnes qui ont, de près ou de loin, joué un rôle dans ma petite vie de thésard. Un grand merci au Pr. Joost van Meerwijk à l’accent légendaire d’avoir accepté d’être le président du jury en ce jour de jugement final et non dernier … bien heureusement ! Quoi que vu le stress et les antécédents cardiaques dans ma famille… bref, excusez-moi je m’éloigne du sujet.
Je remercie également mes deux rapporteurs Jacques Nunes et Emmanuel Donnadieu qui, tels des métronomes, ont analysé et jugé mon travail. Merci à vous et excusez-moi de vous avoir gâché l’été, même si le temps n’était pas génial il faut l’avouer, et dans un tel contexte, la lecture est un passe-temps très apprécié par les gens de qualité.
Que pourrais-je dire de Salvatore Valitutti ou "il cappo" comme il se fait appeler dans le clan (non des siciliens) mais du laboratoire du CPTP INSERM U563 bât. A deuxième étage. J’ai toujours eu l’impression d’être à bord d’une barque avec toi, quelques fois c’étaient les courants rapides et la tempête, d’autres fois, nous retrouvions les eaux calmes. Heureusement, nous n’avons jamais chaviré. Merci pour cette énergie et passion communicatives qui t’animent et qui t’animeront encore et encore. Sabina, toi l’ancienne championne de planche à voile, si tu décidais de faire le tour du monde en voilier, j’aurais justement un matelot à te conseiller, quoique, bref … louange à dieu pour le don que tu as reçu pour la cuisine italienne. Merci aussi à Mattia pour les petits foots et les flippers, malheureusement trop rares au labo. Ma pensée circule et arrive au pic des deux Eric. Mon premier est mordu de musique classique et raffole de la biochimie qu’il m’a apprise. Le deuxième est mordu de plongée sous-marine et m’a fait goûté à des mastocytes aux sauces humaines et murines. Merci à vous deux, Eric Champagne et Eric Espinosa, mon coéquipier de cordée dans l’ascension du mont Masto. Le sommet est encore loin, surtout ne lâche pas la corde. Viens maintenant la diva du labo, la femme fatale par excellence, qui me faisait tant peur à la fac, et qui me fait tant sourire aujourd’hui. Merci ma petite Cécile pour ta présence et ta bonne humeur au quotidien. Mes souvenirs se portent maintenant sur Olivia Newton-Jones (ma petite Perpignanaise préférée, fraîchement Parisienne). Une grosse absence depuis que tu n’es plus là, les rigolades et les soirées arrosées ainsi que les moments de soutien m’ont manqué, mais tel était le destin du couple de Grease version 2007. J’en viens maintenant à la femme de tous les exploits et de tous les défis (politiques, sportifs, scientifiques), bref la technicienne aux deux Immunity et au multi langage (anglais, espagnol, patois… j’en passe et des meilleurs). J’ai nomme Martine Guiraud. Merci pour ton aide scientifiquement parlant ou pas. Encore un peu de patience pour la retraite et tu pourras faire le Kilimandjaro … avec Cricri que je remercie au passage pour son goût très sûr pour les morilles. Merci aussi à toi Mika pour tes paroles toujours justes. J’ai été très content que tu rejoignes l’équipe après ton DEA, notre complicité dans le boulot et dans les soirées va beaucoup me manquer. Mais nous serons emmenés, j’en suis sûr, à nous revoir. Merci à toi Florie pour ton sourire et ta joie de vivre imperturbables, et à toi Seb, "l’intello-véto" pour les discussions scientifiques ou sur la randonnée pédestre. J’ai également une pensée pour Irish (travailleuse invétérée) qui m’a appris les quelques mots de brésilien qui m’ont permis de jouer au futbal sur la plage de copacabana….. Leandro me vient alors à l’esprit, même si nous n’avons travaillé que quelques mois ensemble, nous avons pu construire les bases d’une amitié naissante, le temps fera la suite en ce qui concerne le reste de la bâtisse. Merci pour l’ouragan de gaieté qui a déferlé sur le labo à deux reprises. Merci aux patriarches du labo, Benoît et Mustapha pour leur passage de témoin. Merci aussi à toi Rossana, car même si tu l’ignores, j’ai beaucoup appris en ta présence.
Merci à l’équipe du colonel breton, Loïc Dupré, et à tous ses acolytes, Ronan le barbare, Gema la guerrière et Julie la princesse. Merci beaucoup Loïc pour ces dialogues où tu nous montres une face cachée de WASP.
Je n’oublierais pas non plus les autres équipes de l’unité, à savoir, l’équipe van Meerwijk et sa cohorte de jolies filles qui en compagnie d’Aurélie avaient le monopole de la presse people du CPTP. Merci à toi Julie R., compagne de DEA, qui a toujours eu un mot gentil dans les bons comme dans les mauvais moments. Ton départ a été douloureux mais je retiendrai tous les excellents moments passés en ta compagnie. Après le départ d’Ingrid, l’éditeur en chef de la presse people qui a succédé à Olive, le maestro à l’humour affûté, il reste encore Julie T. (ex-coach de l’équipe de foot de l’INSERM) et Céline (ou la fille aux yeux azurs) assurant ainsi la descendance du labo. Que dire de toi, John Travolta, si je sais : quel déhanché !!! J’aurai du mal à oublier cette soirée disco !!! Quand à Paola, tes sourires et ta présence à mes lab meeting du matin devant trois personnes me manqueront aussi.
Si je parlais de Jean-Charles Guéry, je pense que je reviendrais sur ce tournoi de foot que nous avons gagné. Je suis heureux d’avoir joué avec toi et non contre toi, vus les tartines que tu envoyais au gardien adverse, un grand moment. Je ne t’oublierai pas non plus Sophie, autre compagne de DEA, à qui je souhaite un bel avenir scientifique car tu le mérites. N’oublie pas Cyril que même si je m’en vais, il faudra bien que tu rendes compte de tes dettes envers le labo Valitutti, tu trouveras d’ailleurs sur mon bureau la facture qui t’ai due. Bye, sans rancune. Quant à toi Karine, finalement quand tu disais que le rose m’allait bien pour le costume de Travolta, tu avais raison … quoique.
J’ai également une pensée pour l’équipe du Pr. Lebouteiller et le harem de Hischam mais aussi l’équipe de Roland Liblau où il était joyeux d’aller chercher la clé de l’animalerie, puisqu’il y avait toujours place à une belle discussion foot-sciences avec Lennart, soirées branchées avec Flo, drague avec la capoeriste Nico C. (n’oublie pas que ce soir, c’est démo en maillot) et dialogue sérieux avec Sabine.
Outrage si j’oubliais le clan des Ramsès qui porte d’ailleurs mal ce nom tant les membres de cette équipe indépendante paraissent jeunes. Merci à toi Fatima, la Tina Turner du plateau cytométrie. Je dois t’avouer que j’ai détesté le LSR car j’appréciais beaucoup plus le temps passé en ta compagnie sur le trieur. Je remercie également l’enthousiasme de Geneviève et Maryse, leur dernière chanson pour la thèse d’Aurélie sera un tube, j’en suis sûr.
Merci à toi petite Aline et petite maman du labo qui avec tes sucettes à l’anis (pardon !!! aux fruits) m’a envoûté et réconforté quand il le fallait. Je ne t’oublierai pas et après tout tu ne te débarrasseras pas de moi comme ça ! Grâce à internet, même si je suis loin, je serai toujours près.
Merci aussi à la fée des commandes, Dominique, qui pas sa dextérité et se gentillesse rend souvent les choses faciles et agréables.
Merci aux personnes de l’ombre, Ivan, Jacques et Nordine, pour les petites discussions sympathiques à chaque rencontre dans les couloirs et pour leur disponibilité souvent non remerciée. Et toi Joël, qui règne sur le magasin avec Marie (mon ange gardien), merci encore une fois pour les rigolades et les services rendus. Si tu continues comme cela Joël, je pense que tu passeras les 20 000 commandes l’année prochaine.
Merci aussi à Sophie Allart (montagnarde chevronnée) qui avec Fatima, permettent à toutes les équipes d’avancer dans leurs travaux en imagerie et cytométrie. Merci à toi Gilles D. pour les repas délires entre midi et deux à la pizzeria où l’espace d’un moment les manips étaient loin.
Je ne saurai oublier David, alias « Tooms » ou homme élastique, ce jour là, j’ai vu jusqu’où pouvait aller le contortionnement du corps humain et la volonté qui forge une amitié. Tes Mojitos me manquent beaucoup, les anglais ne connaissent pas la chance de t’avoir…….En espérant que l’avenir nous rapproche.
Vient le tour de Bruno, le DEA nous a fait connaître, la thèse a construit notre relation. Que pourrais je dire de mon coéquipier de Raid surhumain, qui a toujours eu les bons conseils envers moi, tel un grand frère ……Merci d'être comme tu es, toi la personne avec qui j’ai gravi des cols et avec qui le projet futur que tu connais pourrait prendre forme un jour.
Merci à toi Alex, qui a suivi mon parcours depuis la maternelle. Ça fait un bail maintenant mais notre relation n’a pas bougé. Même si la France nous sépare, on n’a jamais été aussi proche. Une pensée pour toi Dana, qui a chaque fois qu’on se voie a toujours la faculté de me faire rire en toute circonstance. La gloubiboulga reviendra à toulouse, j’en suis sûr et tu en seras certainement l’organisatrice….
Arnaud, Jim, la tangente, bref, autant de surnom pour la personne qui me connaît depuis la fac et ses fameuses excentricités………St Gaudens, mimizan, gala dentaire, des moments inoubliables en ta compagnie. Merci à toi, mon ami qui fait toujours son possible pour entretenir notre relation. Grâce à toi, les moments difficiles de la thèse n’avaient aucune raison d’être. Merci pour ta patience, ton écoute, en espérant que l’avenir continue de nous garder ces moments privilégiés.
J’ai également une pensée pour Kazem, le frère que je n’ai jamais eu et qui est à mes côtés dans les bons comme dans les mauvais moments de la vie. Il y aurait tant de choses à dire…………..tu connais l’essentiel. Merci pour l’appui que tu me fournis dans les passages délicats, tu es plus qu’un ami, un guide………Je rends grâce à Dieu de t’avoir rencontré, il y a maintenant 14 ans sur cette marche du Caousou, c’était un jour d’automne de 1993… Merci pour tout.
La mesure de l’amour est de s’aimer sans mesure. Merci à toi ma petite fleur de me supporter et de m’aimer au quotidien…les mots seraient de trop tant tu sais déjà tout… merci ma ninette ou ma raison d’être aujourd’hui.
Enfin, je tiens à remercier mes parents qui ont toujours fait le meilleur pour moi. Je ne vous remercierai jamais assez et les mots n’y suffiraient certainement pas.
Je sais que j’ai beaucoup de chances…Merci.
La conclusion viendra pour mon grand Père, partit beaucoup trop tôt et qui m’a légué tant de valeurs… Ce travail est pour toi. Tu resteras à jamais un modèle pour moi. Merci…
TABLE DES MATIERES
Index des figures 3
Liste des abréviations 5
Résumé 8
Abstract 9
PARTIE 1 : Etude des interactions moléculaires à la synapse immunologique formée entre les lymphocytes T et les CPAs
INTRODUCTION 11
Avant-propos 12
A) Qui sont les lymphocytes T ? 13
1) Origine et développement 13
2) Le complexe TCR/CD3 15
a) Structure du TCR 15
b) Association du complexe TCR/CD3 16
c) Organisation des gènes codant le TCRαβ 18
3) Fonctions effectrices des lymphocytes T 19
a) Les lymphocytes T CD4+ 19
b) Les lymphocytes T CD8+ 22
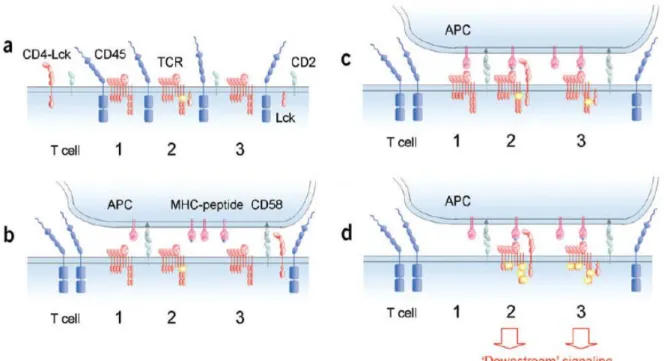
B) La rencontre lymphocyte T/CPA : initiation de la signalisation dans le lymphocyte T 24
1) Haute sensibilité, faible affinité : le paradoxe de la reconnaissance par le TCR 24
a) Problématique 24
b) Les modèles proposés 25
2) Initiation de la signalisation des lymphocytes T : les différents facteurs 29
a) Valence des TCR et rôle des co-récepteurs 29
b) La « kinetic segregation » ou l’organisation de zones privilégiées au site de contact cellule
T/CPA 35
c) Le rôle des microdomaines membranaires dans les deux versants : côté lymphocyte T et
côté CPA 37
d) Changement conformationnel du TCR 39
C) La mise en place des voies de transduction du signal suite à l’engagement du TCR 41
1) Les événements proximaux 41
a) Le complexe CD3 41
b) Les protéines tyrosine kinase de la famille src : Lck et Fyn 43
c) La protéine ZAP-70 46
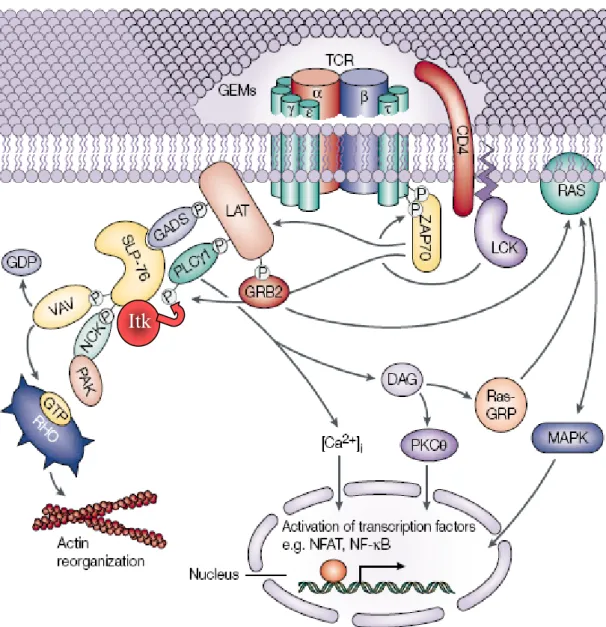
d) Les molécules adaptatrices 49
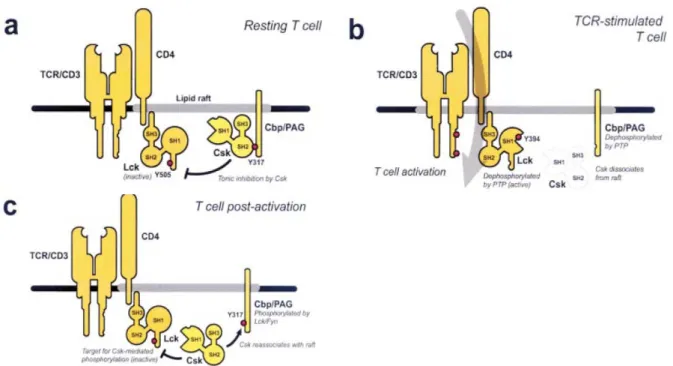
e) Les mécanismes régulateurs de la signalisation 53
2) Le lancement des voies de signalisation 57
a) La voie calcique 57
b) La voie PKCθ/NFκB 68
c) La voie des MAPK 72
d) La voie de la PI3K 77
D) La synapse immunologique ou la communication entre le lymphocyte et la CPA 82
1) Architecture de la synapse immunologique 83
a) La synapse immunologique concentrique 83
b) Les différentes synapses immunologiques 85
c) La molécule CD2 ou le composant de la synapse immunologique aux multiples facettes 88 2) La synapse immunologique, une structure dynamique à l’image des interactions lymphocyte
a) Dynamique de la synapse immunologique 92
b) Dynamique de l’interaction lymphocyte T/CPA 94
3) Mécanismes impliqué dans la formation de la synapse immunologique 96
a) Un exemple de mécanisme passif : « diffusion trapping » 96
b) L’interaction CD2/CD58 : rôle dans l’adhésion et la reconnaissance de l’antigène à la
synapse immunologique 97
c) Un exemple de mécanisme actif : l’implication du cytosquelette d’actine 98
d) Trafic vésiculaire et protéines à domaine PDZ 99
e) Rôle des molécules CD28 et CTLA-4 99
4) Pourquoi créer une synapse immunologique ? 100
a) Rôle dans la « downregulation » des TCR ? 101
b) Rôle du cSMAC dans la polarisation des récepteurs aux cytokines/chimiokines 101
c) Rôle dans la sélection thymique 102
d) Rôle dans la maintenance d’un pool de lymphocytes naïfs périphériques 102
e) Rôle dans la signalisation intracellulaire 103
Objectif du travail 105
RESULTATS 107
CD2 and TCR synergize for the activation of phospholipase Cγ1 /calcium pathway at the
immunological synapse 108
DISCUSSION 113
PERSPECTIVES 119
PARTIE 2 : ETUDE EN COURS
Etude de la capacité des mastocytes à présenter l’antigène
INTRODUCTION 125
A) Les mastocytes 125
1) Origine et développement 125
2) Les différentes modes d’activation des mastocytes 128
3) Reconnaissance des pathogènes et phagocytose 129
4) L’importance de la maturation des mastocytes in vitro 130
B) Mastocytes et lymphocytes T : une relation en plein essor 132
1) Rôle des mastocytes dans le recrutement des lymphocytes T 132
2) Rôle des mastocytes dans la polarisation Th1/Th2 133
3) Mastocytes et lymphocytes T dans l’EAE : exemple d’une pathologie associée 134
4) Le mastocyte : une CPA non conventionnelle ? 135
Objectif du travail 138 MATERIEL ET METHODES 139 RESULTATS PRELIMINAIRES 141 DISCUSSION ET PERSPECTIVES 148 ANNEXE 151 Immunological synapse 152 BIBLIOGRAPHIE 154
INDEX DES FIGURES
Figure 1 : Etapes du développement des thymocytes 14
Figure 2 : Le complexe TCR-CD3 16
Figure 3 : Organisation du complexe TCR-CD3 basée sur l’interaction de résidus transmembranaire 17
Figure 4 : Diversité structurale du TCR 18
Figure 5 : Représentation schématique de l’induction et de la régulation des lymphocytes Th1 et Th2 21 Figure 6 : Orientation de la machinerie lytique au site de contact CTL-CPA 22
Figure 7 : Schéma du Proofreading 26
Figure 8 : Schéma du « serial triggering » : un simple complexe CMH/peptide peut engager et activer
plusieurs TCR durant une interaction cellule T/CPA prolongée 28
Figure 9 : Schéma du modèle d’hétérodimérisation 31
Figure 10 : Le modèle de pseudo-dimérisation 32
Figure 11 : Les TCR multivalents permettent la dissémination du signal à travers les multimères 34
Figure 12 : Le modèle de kinetic segregation 36
Figure 13 : Les structures des bicouches lipidiques 37
Figure 14 : Modèles pour l’organisation de la membrane plasmique 38
Figure 15 : Représentation schématique de p56Lck 43
Figure 16 : Régulation de l’activité catalytique des src kinases 44
Figure 17 : Recrutement et activation de src kinases durant la stimulation canonique du TCR 46
Figure 18 : Représentation schématique de ZAP-70 47
Figure 19 : De multiples évènements sont requis pour l’activation de ZAP-70 48 Figure 20 : Contexte intracellulaire de la régulation et de la fonction de ZAP-70 49 Figure 21 : Structure de différents adaptateurs de la signalisation du lymphocyte T 50 Figure 22 : Régulation positive de la signalisation du TCR par les molécules adaptatrices 52
Figure 23 : Régulation des src kinases par le complexe PAG/Csk 55
Figure 24 : Les voies de signalisation en aval du TCR 57
Figure 25 : Représentation schématique de la structure de la Tec kinase, Itk 58 Figure 26 : Représentation schématique de la structure de la PLCγ1 60 Figure 27 : Contexte intracellulaire où s’effectuent la régulation et la fonction de la PLCγ1 62
Figure 28 : Vue d’ensemble des voies de signalisation calcique 64
Figure 29 : Les trois modèles de communication entre la vidange des stocks calciques et l’ouverture
Figure 30 : La rencontre entre lymphocyte T et DC, le « stop signal » 66
Figure 31 : Représentation schématique de la structure de la PKCθ 68
Figure 32 : Le concept de costimulation pour l’activation de la cellule T 70 Figure 33 : Résumé schématique de la voie d’activation NFκB induite par TCR/CD28 dans le
lymphocyte T 71
Figure 34 : Représentation schématique du cycle des GTPases 73
Figure 35 : Uprégulation de l’expression des gènes JNK durant l’activation des cellules T CD4+ 76
Figure 36 : La production d’IFNγ par les cellules Th1 requiert l’activation des p38 MAPK 77 Figure 37 : Activation et signalisation de la PI3K dans les lymphocytes T 79 Figure 38 : Localisation de la synapse immunologique entre un lymphocyte T et un lymphocyte B 83 Figure 39 : Représentation schématique de la synapse immunologique mature 84 Figure 40 : Vue de face (côté lymphocyte T) de la synapse immunologique mature 84 Figure 41 : Diversité des zones d’interaction moléculaire de la synapse immunologique 86
Figure 42 : Le concept de la synapse immunologique dynamique 88
Figure 43 : Représentation schématique de la structure du CD2 89
Figure 44 : Morphologie et phases d’interaction entre une cellule T et une CPA 94 Figure 45 : Dimensions des molécules impliquées dans la formation de la SI 97 Figure 46 : Caractéristiques majeures des deux sous types de mastocyte chez le rongeur 126 Figure 47 : Principales classes de médiateurs relargués par les mastocytes 127
Figure 48 : Comparatif BMMC/PCMC 131
LISTE DES ABREVIATIONS
ADN / ARN : acide désoxyribonucléique / acide ribonucléique AP-1 : Activator protein-1
BMMC : Bone Marow-derived-Mast Cell Btk : Bruton's tyrosine kinase
Cbl : Casitas B lineage lymphoma CD : cluster of differenciation
CDR : complementarity-determining regions CD2AP : CD2-associated protein
CD2BP : CD2-binding protein
CMH : complexe majeur d’histocompatibilité CPA : cellule présentatrice d’antigène
CR : Complement Receptor
CRAC : Ca2+-released-activated Ca2+channels Csk : C-terminal Src Kinase
CTL : cytotoxic T lymphocyte
CTLA-4 : cytotoxic-T lymphocyte-associated protein 4 DAG : (1,2) Diacylglycerol
DC : cellules dendritiques
EAE : Experimental autoimmune encephalomyelitis Erk : Extracellular signal-regulated protein kinases ERM : Ezrine-Radixine-Moésine
FRAP : Fluorescence recovery after photobleaching FRET : Fluorescence resonance energy transfer
Fyb / SLAP-130 / ADAP: Fyn SH2-binding protein / SLP-76 associated phosphoprotein of 130 kDa / adhesion and degranulation-promoting adaptor protein
Gads : Grb2 related adaptor downstream of Shc GAP : GTPases activating protein
GEF : guanine nucleotideexchange factor GFP : Green Fluorescent protein
GPI : glycophosphatidylinositol Grap : Grb2-related adaptor protein
GSK3 : Glycogène synthase kinase 3 GTP : guanosine 5′-triphosphate ICOS : Inducible T-cell co-stimulator IFN : interferon
Ig : Immunoglobuline IKK : Inhibitor of κB kinase IL : Interleukine
IP3 : inositol (1,4,5) trisphosphate
ITAM : Immunoreceptor tyrosine-based activation motifs
Itk / Emt / Tsk: IL-2-inducible T-cell Kinase / Expressed in mast cells / T cell-specific-kinase JNK : c-Jun NH2-terminal kinases
LAT : Linker for activation of T cells
LAX : Linker for activation of X (unknown) cells Lck : Lymphocyte-specific protein tyrosine kinase LFA-1 : Lymphocyte function-associated antigen-1 LPS : Lipopolysaccharide
MAPK : Mitogen-Activated Protein Kinase
MEK : Mitogen-activated or extracellular signal-regulated protein kinase MOG : Myelin oligodendrocyte glycoprotein
MTOC : Microtubule organizing centre NFAT : Nuclear factor of activated T cells NFκB : Nuclear factor-kappa B
NK cells : natural killer cells
PAG / Cbp(phosphoprotein associated with glycosphingolipid-enrichedmicrodomains) / (Csk-binding protein)
PAK 1 : p21-activated kinase 1
PDK1 : Phosphoinositide-dependent protein kinase 1 PH domain: pleckstrin homology domain
PiP2 : Phosphatidylinositol 4,5-bisphosphate
PI3K : phosphatidylinositol-3-OH kinase PKCθ : Protein kinase Cθ PI(4.5)P2 : phosphatidylinositol 4.5 bisphosphate
PI(3,4,5)P3 : Phosphatidylinositol (3,4,5) trisphosphate PKB : Protein Kinase B
PLCγ1 : Phospholipase Cγ1 PMA : Phorbol myristate acetate PR : prolin rich region
PTP : protéine tyrosine phosphatase PYK-2 : proline rich tyrosine kinase-2 Ras : Rous avian sarcoma
RasGRP1 : Ras Guanyl nucleotide-releasing protein Rlk : Resting lymphocyte kinase
RyR : récepteurs ryanodine SCF : Stem Cell Factor
SH domain : Src homology domain SHP-1 : Src homology phosphatase-1 SI : Synapse immunologique
SLP-76 : SH2-domain-containing leucocyte protein ok 76 kDa SMAC : Supra-molecular activation cluster
Sos : Son of sevenless Syk : spleen tyrosine kinase TCR : T cell Receptor
Tec : Tyrosine kinase expressed in hepatocellular carcinoma TLR : Toll-like receptor
TNF : Tumor Necrosis Factor TRIM : TCR-interacting molecule
VCAM : Vascular cell adhesion molecule
WASP : Wiscott-Aldrich immunodeficiency syndrome protein ZAP-70 : Zeta chain-associated protein of 70 kDaltons
Nicolas Espagnolle
Etude des interactions moléculaires à l’aire de contact formée entre les lymphocytes T et les cellules présentatrices d’antigène
Directeur de thèse : Salvatore Valitutti Toulouse, le 28 Septembre 2007
Résumé
L’interaction entre la cellule T et la cellule présentatrice d’antigène (CPA) est l’événement central de la réponse immunitaire adaptative. Cette rencontre entraîne la réorganisation de plusieurs molécules de surface et de composants de la signalisation intracellulaire au niveau de l’aire de contact cellule-cellule. Cette réorganisation moléculaire est connue sous le nom de synapse immunologique (SI).
A ce jour, les fonctions de la SI ne sont pas encore pleinement élucidées. Afin de comprendre la raison d’être d’une telle structure, dans une première étude, nous avons recherché l’impact que l’engagement de la molécule CD2, avec son ligand CD58 à la SI, a sur la signalisation calcique du lymphocyte T humain stimulé par l’antigène. Ainsi, nous avons montré que l’engagement du CD2 permet le recrutement et l’activation optimale de la PLCγ1 ainsi que l’amplification et le maintien de la mobilisation calcique dans les lymphocytes T stimulés par l’antigène. Sans être intrinsèquement capable d’activer la voie calcique, l’interaction CD2/CD58 permet la modulation de cette voie de signalisation induite par le TCR permettant l’activation totale du lymphocyte T.
Dans une seconde étude, encore en cours, nous avons pour but d’étudier la SI formée entre les lymphocytes T et des CPAs non conventionnelles : les mastocytes. Pour cela, dans un premier temps, nous avons mis au point la méthodologie qui permet de dériver des mastocytes à partir de cellules de la moelle osseuse et dans un deuxième temps, nous étudions, à présent, leur capacité à présenter l’antigène aux cellules T.
Mots clés : synapse immunologique, interaction CD2/CD58, signalisation intracellulaire, cellule
présentatrice d’antigène, mastocytes.
Discipline : Immunologie
Nicolas Espagnolle
Study of molecular interactions at contact area formed between T lymphocytes and antigen presenting cells
Thesis supervisor : Salvatore Valitutti Toulouse, the 28th September 2007
Abstract
T cell/APC interaction is the central event in adaptive immune response. This encounter allows the reorganization of several surface molecules and intracellular signaling components at the cell-cell contact area. This molecular reorganization is named the
immunological synapse (IS).
Actually, IS functions are not completely understood. To understand the « raison d’être » of this structure, in a first study, we searched the impact of CD2 molecule engagement with its ligand CD58 at IS, on calcium signaling in antigen stimulated human T cells. Therefore, we have shown that CD2 engagement permits optimal recruitment and activation of PLCγ1 and plays a key role in sustaining [Ca2+]i increase in antigen-stimulated T cells. Without being intrinsically able to trigger the calcium pathway, CD2/CD58 interaction modulates this TCR induced signaling pathway allowing to full T lymphocyte activation.
In a second study, still going on, we aim to study IS formed between T cells and unconventional APCs : mast cells. For that, in a first time, we developped methods allowing to derive mast cells from bone marrow cells and, in a second time, we are now studying their capacity to present antigen to T cells.
Key words : immunological synapse, CD2/CD58 interaction, intracellular signaling, antigen
presenting cell, mast cells.
Subject : Immunology
PARTIE 1
Etude des interactions moléculaires à la synapse
immunologique formée entre les lymphocytes T et les
AVANT-PROPOS
Des agents infectieux comme les bactéries, les virus ou les parasites d’une part, et l’apparition de cellules néoplasiques d’autre part, constituent des menaces potentielles pour l’organisme des vertébrés. Pour riposter contre l’envahisseur, l’organisme utilise plusieurs mécanismes de défense.
Ainsi le système immunitaire se compose de deux concepts : le système immunitaire inné et le système immunitaire adaptatif. Dans le premier cas, il s’agit d’une réaction rapide, sans mémoire et indépendante de l’antigène, principalement basée sur la reconnaisance de motifs moléculaires associés aux organismes pathogènes ou PAMP (Pathogen-associated molecular pattern) par les PRR (Pattern Recognition Receptor), qui sont des récepteurs distribués à la surface de diverses cellules telles que les cellules dendritiques et les macrophages. Dans le deuxième cas, la réponse adaptative concerne des phénomènes liés aux antigènes, et consiste en la sélection de clones de lymphocytes, capables de cibler ce qui est perçu comme une menace. Cette réponse adaptative est lente, strictement dépendante des antigènes, et possède une mémoire immunitaire.
A ces deux concepts, il est important d’incorporer les différents types de réactions immunitaires : les réactions de type humoral ou cellulaire. Ainsi, l’immunité humorale innée implique des facteurs solubles tels que les anticorps naturels, le complément et les produits dérivés de l’inflammation (leucotriènes, prostaglandines, facteurs tissulaires). Quant à l’immunité humorale adaptative, elle est supportée par la présence d’anticorps circulants (IgM, IgG, IgE, IgA) produits par des lymphocytes B différenciés en plasmocytes. En ce qui concerne l’immunité cellulaire innée, elle implique des cellules qui sont capables de réagir à un phénomène sans éducation préalable et de manière indépendante de l’antigène (cellules NK, macrophages, cellules dendritiques). Enfin, l’immunité cellulaire adaptative met en jeu les lymphocytes T et B. Alors que les lymphocytes B reconnaissent l’antigène sous sa forme native par leur récepteur BCR (B cell receptor), le TCR (T cell receptor) des lymphocytes T reconnaît seulement les fragments antigéniques associés aux molécules du CMH (complexe majeur d’histocompatibilité) disposés à la surface des CPA (cellules présentatrices d’antigène).
L’immunité à médiation humorale (médiée par les anticorps) et cellulaire (médiée par les lymphocytes T) sont les deux principales lignes de défense mises en place par les organismes supérieurs dans la lutte contre les pathogènes microbiens. Alors que les anticorps reconnaissent des antigènes entiers, les lymphocytes T distinguent les éléments étrangers sous la forme de petits fragments peptidiques, dans le contexte d’une présentation par des molécules du CMH (complexe majeur d’histocompatibilité) à la surface de CPA (cellule présentatrice d’antigène).
Il existe deux types de lymphocytes T : les lymphocytes Tαβ très fortement majoritaires et les lymphocytes Tγδ (5% du total des lymphocytes T seulement).
Durant mon travail de thèse, nous nous sommes exclusivement intéressés aux lymphocytes Tαβ.
A) Qui sont les lymphocytes T ?
1) Origine et développement
Les lymphocytes T dérivent d’une cellule souche pluripotente provenant de la mœlle osseuse. Les progéniteurs hématopoïétiques colonisent ensuite le thymus (organe lymphoïde primaire), où ils subissent une intense prolifération, permettant ainsi la constitution d’un grand nombre de thymocytes immatures. Le développement des thymocytes est caractérisé par l’expression ordonnée de plusieurs molécules de surface, incluant les co-récepteurs cluster of differenciation CD4 et CD8.
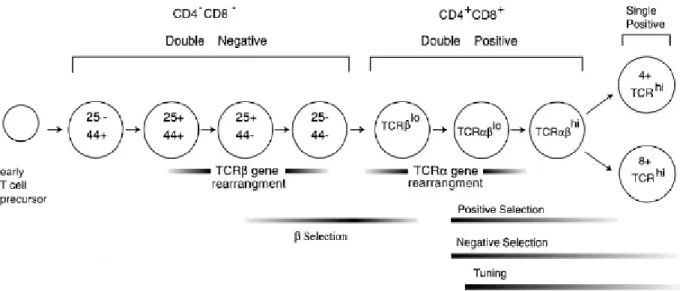
La maturation des thymocytes peut être divisée en trois grandes parties : La première étape de cette maturation est caractérisée par la présence de cellules double négative (DN) CD4-CD8-. Suite à ceci, la deuxième étape est symbolisée par le développement de cellules double positive (DP) CD4+CD8+ et enfin, la dernière étape constitue la formation de cellules matures simple positive (SP) CD4+CD8- ou CD8+CD4- (Figure 1). Toutes ces étapes sont sous la direction d’un paramètre prédominant : le TCR (T cell receptor). En effet, le récepteur des cellules T est le facteur qui va permettre de sélectionner les thymocytes suivant leur spécificité. Le TCR est constitué de deux chaînes α et β. Dans un premier temps, à l’étape double négatif (DN), les thymocytes immatures (constituant approximativement 1-5% des thymocytes totaux) réarrangent le locus codant la chaîne β du TCR, emmenant ainsi à l’expression de cette même chaîne β qui en association avec un substitut de la chaîne α (pTα) à la surface des thymocytes formera le complexe pré-TCR. Ce dernier entraînera la
prolifération et l’expansion de ces thymocytes ; un processus nommé la sélection β (Saint-Ruf
et al. 1994), (von Boehmer et al. 1997).
Figure 1 : Etapes du développement des thymocytes. Au cours de leur maturation, les thymocytes vont
réarranger les gènes du récepteur T afin de produire un TCR αβ fonctionnel. Sa restriction et sa spécificité seront par la suite testées par deux mécanismes de sélection : la sélection positive et négative. Modèle murin d’après (Sebzda et al. 1999)
Les signaux transmis par le complexe pré-TCR non seulement induisent l’expansion, mais aussi le réarrangement du gène du TCRα, et de manière parallèle l’expression des co-récepteurs CD4 et CD8. Les thymocytes, à l’étape double positif CD4+CD8+, représentent 80-90% des thymocytes totaux. Après avoir acquis l’expression d’un TCRαβ mature, les thymocytes CD4+CD8+ seront sélectionnés pour leur capacité à reconnaître des antigènes présentés par les molécules du CMH du soi. Ainsi, cette étape du développement est appelée
la sélection positive et a lieu dans une région périphérique du thymus, le cortex. Les
thymocytes entrent donc en interaction avec des cellules stromales thymiques qui leur serviront de juges telles que les cellules épithéliales corticales (cTec) (Benoist et al. 1989), (Marrack et al. 1997). Ces interactions ont d’ailleurs été illustrées par microscopie bi-photonique (Bousso et al. 2002).
Par la suite, les thymocytes sélectionnés positivement vont connaître une nouvelle sélection qui dépendra de leur faculté à reconnaître des antigènes du soi, synonyme de danger potentiel pour l’organisme. En effet, les thymocytes possédant un TCR auto réactif seront éliminés, c’est la sélection négative. Les thymocytes migrent alors vers un autre territoire thymique plus central, qui sera le siège de cette sélection, la medulla. Afin d’étayer ceci, il a été montré par les travaux de Bravo et coll. que des souris déficientes pour RelB, présentant des désordres de l’organisation thymique accompagnés d’une atrophie de la medulla,
présentent un défaut de la sélection négative et par conséquent une auto-immunité en périphérie (Weih et al. 1995). Deux types cellulaires interviennent dans la sélection négative des lymphocytes T : les cellules épithéliales médullaires (mTec) et des CPA de la medulla (majoritairement des cellules dendritiques, DC). Les mTec présentent des antigènes issus de la synthèse de protéines normalement synthétisées en périphérie. Cette expression ectopique est sous la dépendance de la protéine AIRE (autoimmune regulator) (Anderson et al. 2002), (Villasenor et al. 2005). Les thymocytes reconnaissant les antigènes présentés par les mTec seront rendus fonctionnellement inactifs (anergiques). Parallèlement à ceci, les thymocytes reconnaissant les antigènes à la surface des CPA seront, quant à eux, délétés (Page et al. 1996). La délétion clonale et l’anergie des thymocytes font partie de ce qui est appelé la tolérance centrale. Malgré ces étapes de sélection, certains lymphocytes auto réactifs parviennent à être exportés hors du thymus. Ils seront alors contrôlés par les mécanismes de la tolérance dite périphérique, impliquant notamment les lymphocytes T régulateurs CD4+CD25+.
Deux types de lymphocytes T dits « naïfs » sortiront du thymus : les lymphocytes T D4+
) Le complexe TCR/CD3
C restreints aux antigènes présentés exclusivement par le CMH II (Berg et al. 1989), (Kaye et al. 1989) et le lignage des lymphocytes T CD8+ restreints aux antigènes présentés exclusivement dans le contexte du CMH I (Teh et al. 1988), (Sha et al. 1988).
2
ans la réponse immunitaire à médiation cellulaire est la
) Structure du TCR
L’évènement primaire d
reconnaissance de l’antigène par le récepteur des cellules T, le TCR. a
mposé de deux chaînes polypeptidiques glycosylées (α et β) appartenant Le TCR est co
à la super famille des immunoglobulines (Ig). Chaque chaîne est composée d’un domaine variable (V) amino terminal et d’un domaine constant (C) carboxyterminal. Les deux chaînes (α et β) sont reliées entre elles par un pont di-sulfure (Garcia et al. 1996), (Schumacher 2002) (Figure 2). L’association de la partie variable des deux chaînes du TCR est le siège de reconnaissance du complexe CMH/antigène. Plus précisément, ces sites de liaison antigénique ont été prédits comme analogue aux CDR (complementarity-determining regions) des immunoglobulines (Chothia et al. 1988). Les CDR1 et 2 se lient préférentiellement au CMH
alors que le CDR3 se lierait préférentiellement au peptide antigénique (Jorgensen et al. 1992), (Sant'Angelo et al. 1996).
b) Association du complexe TCR/CD3
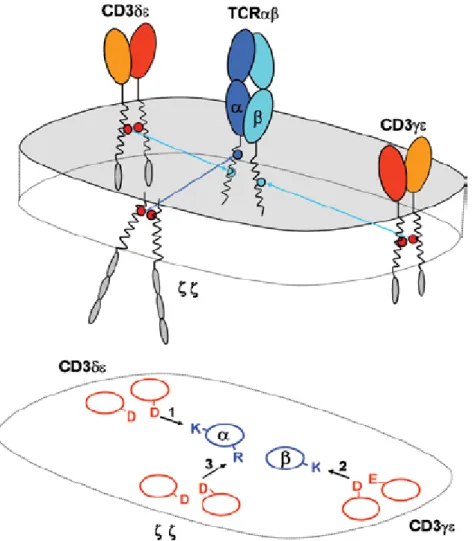
Figure 2 : Le complexe TCR-CD3. D’après (Schumacher 2002).
Des expériences utilisant des anticorps dirigés contre le récepteur ont eu pour conséquence l’identification de quatre polypeptides distincts qui étaient associés de façon non covalente avec l’hétérodimère du TCR : les chaînes CD3γ, CD3δ et CD3ε et l’homodimère de la chaîne ζ (Samelson et al. 1985), (Clevers et al. 1988). Les chaînes CD3γ, CD3δ et CD3ε sont constituées d’un domaine extracellulaire aminoterminal de type Ig, d’un domaine transmembranaire chargé négativement qui permet aux chaînes CD3 d’interagir avec le TCR et enfin d’un domaine intracytoplasmique carboxyterminal. La chaîne CD3ζ quant à elle possède un très court domaine extracellulaire mais un long domaine intracytoplasmique. Il est bon de noter que le TCR ne possède qu’un court domaine cytoplasmique dépourvu de toute activité enzymatique intrinsèque. La fonction de signalisation sera donc assurée par le complexe CD3. En effet, les domaines intracytoplasmiques contiennent des séquences renfermant des motifs ITAM (Immunoreceptor tyrosine-based activation motifs : YxxL(X)6-8YxxL). Après avoir été phosphorylés, ces ITAMs servent de points d’ancrage pour des protéines importantes de la signalisation (Samelson et al. 1985).
La stoechiométrie du complexe TCR/CD3, la séquence de son assemblage aussi bien que la nature des interactions entre les sous unités ont longtemps été énigmatiques. Il semble résolu que le complexe TCR/CD3 soit constitué d’hétérodimères TCRαβ, CD3εγ, CD3εδ et de l’homodimère CD3ζζ (Call et al. 2002), (Call et al. 2005) et Figure 3.
Figure 3 : Organisation du complexe TCR-CD3 basée sur l’interaction de résidus transmembranaire.
Chaque étape d’assemblage résulte dans la formation d’une interface à trois hélices dans la membrane impliquant un résidu basique du TCR (bleu) et une paire de résidus acides à partir des dimères CD3. D’après
(Call et al. 2005)
La formation du complexe se réalise dans le réticulum endoplasmique selon des mécanismes rigoureusement contrôlés. La chaîne α du TCR va tout d’abord s’associer avec l’hétérodimère CD3εδ, tandis que la chaîne β s’associera avec le CD3εγ. Les deux trimères ainsi formés s’associeront via un pont disulfure entre les chaînes α et β pour former l’hexamère TCRαβCD3εγCD3εδ. Après association avec l’homodimère ζζ, le complexe
TCR/CD3 fonctionnel sera exporté à la surface de la cellule (Alarcon et al. 1988), (Huppa et al. 1997).
Non seulement le complexe CD3 est nécessaire pour la signalisation en aval du TCR mais il permet aussi l’expression stable du TCR en surface.
c) Organisation des gènes codant le TCRαβ : la prévention du risque
Etant donné que l’organisme ne peut pas prédire quels seront les antigènes dérivés de pathogène qui seront rencontrés, le système immunitaire s’appuie sur la génération et la maintenance d’un répertoire très diversifié pour le TCR. Ainsi, la taille et la diversité d’un répertoire de cellules T naïves sont cruciales dans l’élaboration d’une réponse immunitaire pour un antigène donné.
Concrètement et sur la base du modèle de réarrangement des gènes des Ig, les chaînes α et β du TCR sont générées via la recombinaison de segments dits variable (V), joining (J),
diversity (D) ajoutés à un segment de la région constante (C) pour le locus β et une
association des segments V-J-C seulement pour le locus α (Nikolich-Zugich et al. 2004) (Figure 4).
Figure 4 : Diversité structurale du TCR. La chaine β est composée à partir d’une recombinaison des segments
V(variable), D(diversity), J(joining) and C(constant), alors que la chaine α est composée des segments V-J-C. La diversité recombinatoire est de plus amplifiée par l’addition ou la délétion de nucleotides (N) aux jonctions
entre les segments. D’après (Nikolich-Zugich et al. 2004).
La destruction ciblée de l’enzyme TdT (terminal deoxyribonucleotidyl transferase) entraîne une réduction de 10 fois dans la diversité du TCR (Gilfillan et al. 1993). De ce fait, la TdT est essentielle pour l’addition efficace de nucleotides au niveau des régions CDR3 pendant la recombinaison V(D)J (Cabaniols et al. 2001).
« En même temps qu'elle nous révèle la particularité de chacun, la biologie nous rappelle la fraternité de tous. »
Jean ROSTAND
3) Fonctions effectrices des lymphocytes T
Suite à leur maturation dans le thymus, les lymphocytes T CD4+ ou CD8+ naïfs migrent vers les organes lymphoïdes secondaires (ganglions lymphatiques, amygdales…) où ils rencontreront des cellules dendritiques leur présentant des complexes CMH/peptide. Ces interactions entraînent l’activation, la prolifération et la différenciation des lymphocytes T. Ainsi, les cellules T CD4+ reconnaissant les peptides dans le contexte du CMH II se différencieront en lymphocytes T de deux types : les T helpers 1 (Th1) et les Th2.
Les lymphocytes Th1 produiront des cytokines pro inflammatoires, comme l’interleukine-2 (IL-2), l’interferon γ (IFNγ) et le Tumor Necrosis Factor α (TNFα) impliquées dans la réponse contre les pathogènes intracellulaires (virus, bactéries…) en agissant entre autres sur la CPA, alors que les lymphocytes Th2 sécrétant de l’4, 5, IL-10 et IL-13 sont plutôt impliqués dans la lutte contre les pathogènes extracellulaires (helminthes …). Ajouté à ceci, les Th2 auront un rôle dans l’activation des éosinophiles par l’intermédiaire de l’IL-5 et 13 mais aussi dans l’inhibition des réponses inflammatoires (IL-4,10 et 13).
Récemment, il a été découvert un nouveau lignage, les Th17 produisant de l’IL-17 et de l’IL6, présentant entre autres des similarités avec les cellules T régulatrices (Weaver et al. 2006).
De manière parallèle, les cellules T CD8+ reconnaissant les antigènes présentés par le CMH I se différencieront en lymphocytes T cytotoxiques (CTL) qui auront pour but d’éliminer les cellules infectées par un virus ou les cellules cancéreuses.
a) les lymphocytes T CD4+ : l’histoire
Les lymphocytes Th1/Th2 : les frères ennemis
Au début des années 1970, il était généralement accepté que les cellules T pouvaient être divisées en deux groupes distincts sur la base de leurs marqueurs de surface : CD4 et CD8. Cependant une évidence accumulait les preuves comme quoi les cellules T CD4+ étaient hétérogènes (Marrack et al. 1975) et il a été formellement montré en 1986 que les cellules CD4+ pouvaient être divisées en deux groupes, désignés Th1 et Th2 sur la base de leur profil
de production de cytokines (Mosmann et al. 1986). Cette conclusion a ensuite été étendue à l’Homme (Parronchi et al. 1991). En tant que cellules « helpers » (aide aux lymphocytes B), les Th1 et Th2 agissent sur la commutation de classe des Igs produites par les lymphocytes B. En effet, les Th1, par le biais de l’IFNγ, dirigent la production d’IgG2a alors que les lymphocytes Th2 permettent l’augmentation de la synthèse d’IgE et d’IgG1. A partir des années 1990, l’utilisation de clones spécifiques pour des bactéries intracellulaires comme
Mycobacterium Tuberculosis ou pour des helminthes extracellulaires (Toxocara canis)
ajoutait des pierres à l’édifice (Del Prete et al. 1991). Ainsi, il a été montré que des clones spécifiques pour Mycobacterium produisaient de l’IFNγ alors que les clones spécifiques de Toxocara étaient principalement producteurs d’IL-4/5.
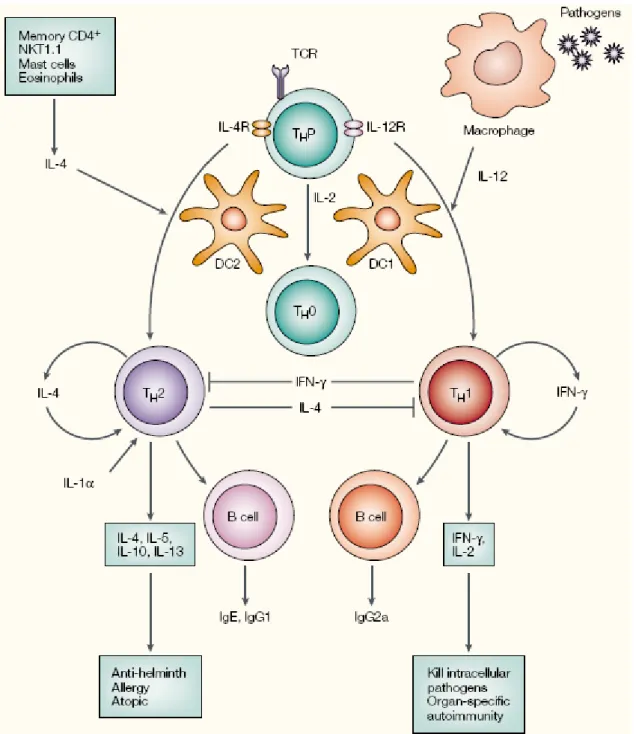
Plusieurs facteurs jouent un rôle prépondérant dans le développement Th1/Th2. Tout d’abord il a été montré dans les années 1990 que l’IL-4 était essentielle pour l’induction de cellules Th2 à partir de précurseurs naïfs (Le Gros et al. 1990). Puis en 1993, Hsieh et coll. montraient que l’IL-12 était requise pour l’induction de cellules Th1 (Hsieh et al. 1993). Ces pensées établissaient que les cellules Th1 et Th2 possédaient un précurseur commun et que le micro-environnement cytokinique était le facteur déterminant dans le développement du lignage Th (Figure 5).
Cependant, des travaux mettaient en valeur que plusieurs autres facteurs pouvaient influencer le développement sélectif des cellules T Th1 et Th2. Ceux-ci incluaient la dose d’antigène (Parish et al. 1972) (Constant et al. 1995), l’affinité pour les antigènes, l’haplotype du CMH (Constant et al. 1995) et les facteurs co-stimulateurs. En accord avec cette dernière idée, une pensée émergeante prône qu’il existerait 2 types distincts de cellules dendritiques (DC) : les DC1 et les DC2 induisant réciproquement les lymphocytes T Th1 et Th2 (Rissoan et al. 1999). Ainsi, les facteurs de co-stimulation liés à la membrane, tels que B7-1 (CD80)/B7-2 (CD86) (Kuchroo et al. 1995) et ICOS (inducible T-cell co-stimulator) (Coyle et al. 2000) ont aussi été montrés comme facteurs d’influence du développement Th1/Th2.
L’IL-12 et l’IL-4 ont pour principale origine la réponse immunitaire innée. Les macrophages (Gazzinelli et al. 1993) et les DC (Macatonia et al. 1995) sont les principaux producteurs d’IL-12. Il est beaucoup moins bien établi quelles sont les sources initiales de l’IL-4 ainsi que ses mécanismes d’induction. Plusieurs candidats sont toutefois cités : les cellules NKT (natural killer T cells) (Yoshimoto et al. 1994), les mastocytes (Plaut et al. 1989), les basophiles (Seder et al. 1991), ainsi que des lymphocytes T CD4+ matures (Schmitz et al. 1994).
Pour conclure ce chapitre, il est important de noter que l’IFNγ et l’IL-4 produits respectivement par les lymphocytes T Th1 et Th2 peuvent agir comme facteurs de croissance autocrine pour le groupe d’intérêt mais aussi comme facteurs inhibiteurs pour le groupe opposé (Fernandez-Botran et al. 1988) (Figure 5).
Figure 5 : Représentation schématique de l’induction et de la régulation des lymphocytes Th1 et Th2.
Fonctionnellement, les cellules T Th1 médient la destruction de pathogènes intracellulaires et l’autoimmunité spécifique d’organe via l’activation de cellules cibles par l’IFNγ. Les Th2, pour leur part, sont anti-helminthes et
b) les lymphocytes T CD8+ : les « killers »
Suite à la rencontre des lymphocytes T CD8+ naïfs avec une CPA présentant un complexe CMH/peptide agoniste, une activation suivie d’une différenciation du lymphocyte T CD8+ intervient. Grâce à l’aide apportée par les lymphocytes Th1 producteurs d’IL2, les cellules T CD8+ deviendront des CTL.
Comme mentionné plus haut, les CTL auront pour but de débarrasser l’organisme de cellules infectées par un virus ou cancéreuses. Pour cela les CTL utilisent plusieurs mécanismes. Tout d’abord, les cytokines (comme le TNFα et l’IFNγ ayant une action cytotoxique) sont sécrétées à proximité des cellules cibles entraînant ainsi leur mort.
Le mécanisme cytotoxique de référence est la voie perforine/granzyme (voie dominante dans les CTL). Suite à la rencontre avec l’antigène, les CTL polarisent leur machinerie lytique vers le site de contact CTL/cellule cible (Yannelli et al. 1986) (Figure 6).
CTL CPA
Figure 6 : Orientation de la machinerie lytique au site de contact CTL-CPA. La Perforine (bleu) et le LTR
(Lyso Tracker red) (rouge), marqueurs des lysosomes sécrétoires sont orientés au site de contact CTL/CPA chargé en peptide antigénique, vu par microscopie confocale. D’après (Faroudi et al. 2003)
Suite à cette polarisation, la perforine relarguée par les CTL induit la mort des cellules cibles en formant des pores dans la membrane plasmique (Henkart et al. 1982). En plus d’une destruction membranaire, la lyse est accompagnée d’une détérioration cellulaire qui est, quant à elle, médiée par des sérines protéases appelées granzymes (Lowin et al. 1995). Ces protéases induisent une apoptose rapide des cellules cibles (Heusel et al. 1994) après leur entrée dans ces mêmes cellules facilitée par les pores membranaires formés par la perforine (Peters et al. 1991). Les granzymes prédominants dans la lyse médiée par les CTL sont les granzymes A et B. Les granzymes B induisent l’apoptose de la cellule cible via une voie dépendante des caspases (Sutton et al. 2003) tandis que les granzymes A emmènent à un programme de mort cellulaire indépendant des caspases (Beresford et al. 2001).
Les CTL peuvent également utiliser une autre voie pour tuer leurs cibles : la voie Fas/Fas-Ligand (Ostergaard et al. 1987). La stimulation du récepteur de mort Fas (membre de la famille du Tumor Necrosis Factor, TNF) situé sur la cellule cible permet l’activation de la caspase 8 (Scaffidi et al. 1998). Il a été suggéré que Fas-Ligand soit stocké dans les mêmes granules que ceux contenant la perforine et les granzymes, ainsi le relargage de Fas-Ligand sur la cellule cible favoriserait l’interaction Fas/Fas-Ligand, puis par conséquence, la mort des cellules cibles (Bossi et al. 1999). Cela dit, il existe très peu de Fas-Ligand stocké dans les CTL, par conséquent une cytotoxicité optimale par cette voie ne pourra avoir lieu que lorsque de nouveaux ligands auront été synthétisés, soit quelques heures après la stimulation du TCR.
Les CTL possèdent deux modes d’induction de mort contre les cellules dangereuses pour l’organisme. Cependant, l’utilisation de la voie Perforine/Granzyme est prépondérante due à sa rapidité et à son efficacité d’exécution.
B) La rencontre lymphocyte T/CPA : initiation de la signalisation dans le
lymphocyte T
1) Haute sensibilité, faible affinité : le paradoxe de la reconnaissance par le TCR
a) Problématique
La reconnaissance du complexe CMH/peptide antigénique par le TCR est l’évènement central de la réponse immunitaire adaptative. Comprendre quels sont les paramètres qui dirigent cette interaction est donc d’un grand intérêt. Depuis des années, des chercheurs ont décrit, analysé et tenté d’expliquer les différentes facettes de cette interaction.
Les premières mesures d’affinité du TCR pour les complexes CMH/peptide ont été faites par Matsui et coll. (Matsui et al. 1991) et Weber et coll. (Weber et al. 1992). L’affinité du TCR pour son ligand se révélait être curieusement faible (Kd approximativement entre 10-7 et 10-4 M).
Par ailleurs, le développement de la technologie BIACORE® (utilisant la résonnace plasmonique de surface) a permis de connaître la demi-vie d’association du complexe TCR/CMH/peptide, soit quelques secondes (Matsui et al. 1994) (Davis et al. 1998). L’interaction TCR/CMH/peptide montre alors des paramètres cinétiques particuliers ; un taux d’association lent et un taux de dissociation rapide. Tous ces résultats emmènent à une question : Comment une si faible interaction peut elle engendrer une transduction de signal ?
Un autre point qui emmène à réflexion est le fait que le nombre de ligands spécifiques du TCR, présentés à la surface de la CPA est très faible. En effet, les complexes CMH/Peptide spécifiques sont « noyés » dans une mer de complexe CMH/peptide du soi provenant de la dégradation et présentation successive des protéines intra et extracellulaires de l’environnement. Cependant, 60-200 molécules de complexe CMH/Peptide spécifiques sur la CPA sont suffisantes pour entraîner une réponse lymphocytaire T ; ceci représente seulement 0.03% des molécules du CMH sur la CPA (Demotz et al. 1990). Les lymphocytes T doivent donc être très sensibles et très spécifiques pour pouvoir engendrer un signal à partir de petites concentrations de ligands spécifiques parmi une marée de ligands non spécifiques.
b) Les modèles proposés
Le modèle du « Kinetic Proofreading » (McKeithan 1995)
Initialement, le kinetic proofreading a été développé par Hopfield (Hopfield 1974) afin d’expliquer la remarquable exactitude de la réplication de l’ADN et de la synthèse protéique. Ainsi, il y était décrit et cité que « plusieurs évènements indépendants dans la reconnaissance de substrat combineraient pour augmenter la fidélité de réponse résultant d’une très petite différence dans l’énergie de liaison entre une interaction correcte et incorrecte ». S’appuyant sur ces idées, en 1995, McKeithan transpose ce modèle à la transduction de signal induite par le TCR (McKeithan 1995). Il souligne, tout d’abord, qu’un délai dans le temps sépare la liaison initiale de la production d’un signal majeur d’activation, incluant plusieurs étapes enzymatiques avant la génération de seconds messagers. Une conséquence directe à ceci est le fait que des complexes non spécifiques ayant un taux de dissociation rapide n’arrivent pas à produire un signal avant leur dissociation, alors que, dans les mêmes concentrations, un complexe spécifique le génère.
L’hypothèse du modèle est la suivante (Figure 7) : 1) Le complexe récepteur-ligand spécifique ou non spécifique C0 est converti via une série d’intermédiaires Ci en un complexe actif CN. Chacune de ces étapes requiert de l’énergie et implique la phosphorylation sur tyrosines. 2) La dissociation du complexe emmène à la réversion des modifications, par le biais de phosphatases par exemple. 3) Le taux de dissociation de complexes non spécifiques est suffisamment haut, afin que la dissociation intervienne avant que ce complexe génère des signaux.
Ainsi, ce modèle se base sur le fait qu’il y a un laps de temps nécessaire pour compléter la cascade des intermédiaires, entre l’engagement du TCR et l’initiation de la signalisation. Depuis des années, l’utilisation de variants peptidiques induisant soit l’inhibition d’un signal (antagoniste) ou n’induisant que l’activation partielle du lymphocyte T (agoniste partiel) aide à comprendre les mécanismes de l’engagement du TCR. Dans le cas des peptides altérés (APL : altered peptide ligand), agonistes partiels ou antagonistes, ayant une durée d’interaction trop courte avec le TCR, la cascade des intermédiaires ne peut pas arriver jusqu’à son but final et ainsi n’engendre que des activations partielles du lymphocyte T engagé (Lyons et al. 1996).
Figure 7 : Schéma du Proofreading. Des complexes initiaux C0 formés entre le TCR (T) et le complexe
CMH/peptide (Mx) doivent subir N modifications avant de générer un complexe actif C
N. A chaque étape, le
complexe peut se dissocier emmenant à une complète réversion des sous unités à leur état basal. Pour une totale activation, les signaux doivent être générés à partir du complexe final CN. D’après (McKeithan 1995).
En 1996, une étude de Rabinowitz et coll. élargissait la vision du kinetic proofreading en proposant que, suivant la stimulation par un antagoniste, l’activation incomplète du récepteur, autrement dit une cascade d’intermédiaires incomplète, est inhibitrice pour la réponse de la cellule T (Rabinowitz et al. 1996). En d’autres termes, l’utilisation de façon stérile de l’énergie et du recrutement de molécules expliquerait le pouvoir inhibiteur qu’un antagoniste peut avoir sur un agoniste.
Le modèle du kinetic proofreading peut expliquer la fascinante habilité discriminatoire du TCR, en prédisant que la signalisation du lymphocyte T induite par une faible concentration de complexes CMH/peptide spécifiques sera plus forte que celle induite par une grande concentration de complexes CMH/peptide non spécifiques à la surface d’une CPA.
Ce modèle pourrait aussi expliquer certains mécanismes intervenant dans la sélection thymique mettant en jeu des différences dans l’affinité des interactions TCR/complexe CMH/peptide (Alam et al. 1996).
Enfin, pour illustrer ce qui pourrait être un intermédiaire de la chaîne d’activation décrite dans le schéma du kinetic proofreading, Risueno et coll. ont récemment observé qu’un changement conformationnel avait lieu au niveau de la chaîne CD3ε suite à une stimulation par un agoniste et non par un antagoniste (Risueno et al. 2005).
Le modèle du « Serial Triggering » (Valitutti et al. 1997)
Comme nous l’avons vu plus haut, l’interaction TCR/CMH/peptide est caractérisée par un fort taux de dissociation avec une demi-vie d’association allant de quelques secondes à quelques minutes. Ces résultats apparaissaient alors paradoxaux : il était difficile d’envisager comment des TCR de faibles affinités pouvaient permettre aux cellules T de reconnaître un
petit nombre de complexes CMH/peptide distribués sur la CPA (Demotz et al. 1990). En 1995, Valitutti et coll. s’appuyant sur le fait que l’internalisation et le recyclage du TCR est une conséquence de la liaison spécifique avec son ligand (Padovan et al. 1993) (Krangel 1987), expliquait le paradoxe de la haute sensibilité-faible affinité du TCR par le « serial triggering » (Valitutti et al. 1995). Ce dernier expliquait qu’un petit nombre de complexes CMH/peptide peuvent engager des associations en série avec un très grand nombre de TCR emmenant à un signal soutenu et amplifié. Le fort taux de dissociation du TCR est un instrument de ce mécanisme puisqu’il permet à un simple complexe CMH/peptide d’engager plusieurs TCR par étapes successives de liaison-engagement-dissociation. Ainsi, dans des conditions physiologiques les lymphocytes T peuvent être activés par l’engagement monovalent et séquentiel de plusieurs TCR par un petit nombre de ligands. En accord avec ce modèle, un temps minimum de liaison est requis pour entraîner la transduction de signal et l’internalisation après engagement du TCR. Le modèle de « serial triggering » est illustré par la figure 8.
Le modèle assume que le processus « serial triggering » prend place dans une aire limitée du contact cellule T/CPA et sera par conséquent dépendant de trois facteurs : 1) L’adhérence de la cellule T facilitant l’interaction TCR-CMH-peptide ; 2) Un approvisionnement continu de composants de la signalisation au niveau des TCRs engagés ; 3) Un approvisionnement de nouveaux TCR afin de remplacer ceux qui sont internalisés après engagement.
Dans le cas des peptides altérés, il a été établi que des complexes CMH/agonistes partiels ou encore CMH/antagonistes associés à des TCR avaient des taux de dissociation plus fort comparés aux peptides agonistes (Lyons et al. 1996). De ce fait, la dissociation prématurée du TCR avant une pleine activation reflèterait le comportement du TCR à l’égard des peptides altérés. Ainsi, il est possible que la liaison pour un temps court pourrait résulter dans une activation partielle, alors qu’une liaison encore plus courte pourrait utiliser des TCR de façon stérile sans signal détectable. L’importance de la demi-vie du complexe TCR-CMH/peptide doit donc être optimale (Kalergis et al. 2001).
De plus le modèle de « serial triggering » explique deux caractéristiques clés de la reconnaissance antigénique : la sensibilité et la spécificité. La sensibilité est expliquée par le fait qu’un petit nombre d’agonistes avec des cinétiques de liaison optimales peuvent engagés beaucoup de TCR et emmener ainsi à l’activation de la cellule T. Le fait que de petites déviations dans ces cinétiques optimales peuvent dramatiquement inhiber la capacité
d’engager en série les TCR explique le haut niveau de spécificité de la reconnaissance antigénique (Valitutti et al. 1997).
Figure 8 : Schéma du « serial triggering » : un simple complexe CMH/peptide peut engager et activer plusieurs TCR durant une intéraction cellule T/CPA prolongée. a) Engagement d’un TCR par un complexe
CMH/peptide pour un temps suffisant pour initier la signalisation. b) Le TCR engagé se dissocie et le ligand CMH/peptide est laissé libre d’engager un autre TCR. c) Les TCR engagés transduisent du signal puis sont
internalisés. D’après (Valitutti et al. 1997).
Hudrisier et coll. ont montré que la réponse fonctionnelle des CTL dépend de la fréquence de l’engagement en série des TCR car une liaison covalente du TCR avec le complexe CMH/peptide, autrement dit un engagement des TCR permanent, résulte dans l’abolition de la signalisation soutenue dans le lymphocyte T (Hudrisier et al. 1998).
Des modèles mathématiques ont été développés afin d’estimer les rôles de chacun des deux modèles dans l’internalisation du TCR. Il en résultait l’apparition d’un nouvel état de
TCR (activé, dissocié à partir du complexe CMH/peptide mais encore prêt à être internalisé) non présumé par les deux modèles précédents. De plus le modèle prédit aussi que la fenêtre de demi-vie de liaison TCR-CMH/peptide dépend de la concentration et de la localisation de molécules de signalisation intracellulaires (Coombs et al. 2002).
En résumé, nous pourrions dire que le « kinetic proofreading » se focalise au niveau du récepteur individuel. En effet, pour devenir activé, un TCR lié doit compléter une série de modifications biochimiques (phosphorylations des ITAMs, association puis activation de la ZAP-70 (Zeta chain-associated protein of 70 kDa), recrutement et phosphorylation d’autres molécules de signalisation additionnelles), avant de se dissocier avec le complexe CMH/peptide). Par contre, le modèle du « serial triggering » porte à discuter au niveau cellulaire. Puisque, dans des conditions physiologiques où la densité de CMH/peptide spécifique sur la CPA est faible, la demi-vie de liaison du complexe TCR/CMH/peptide doit être assez courte pour permettre à un seul peptide d’engager en série de multiples TCR requis pour l’activation de la cellule T.
2) Initiation de la signalisation du lymphocyte T : les différents facteurs
a) Valence des TCR et rôle des co-récepteurs
La reconnaissance antigénique par la cellule T montre des parallèles avec le système nerveux (Steinman 2004), à l’image des cellules de la rétine chez les rongeurs pouvant détecter un seul et simple photon, il est suspecté que les lymphocytes T sont capables de détecter un seul et simple ligand agoniste. Mais il existe des seuils différents afin d’emmener à une réponse irréversible (Krogsgaard et al. 2005).
Il a été montré, depuis plusieurs années, que le pontage des complexes TCR par des anticorps induit l’activation de la cellule T (Kappler et al. 1983) (Kolanus et al. 1993). Cette agrégation des TCR, requis pour l’activation, est confortée par des études de stimulation par des multimères de complexes CMH/peptide qui se révèlent être de meilleurs stimuli comparés aux monomères et dimères des mêmes complexes (Boniface et al. 1998). Cependant, il ne faut pas perdre à l’esprit plusieurs contradictions à ce principe, et notamment le fait que les complexes CMH/peptide agonistes sont monovalents et exprimés à de faibles concentrations à la surface des CPA. De plus, en 1996 les travaux de Sykulev ont pu montrer qu’un seul complexe CMH/peptide était suffisant pour induire la cytotoxicité par les CTL (Sykulev et al.
1996). En ce qui concerne les lymphocytes T CD4, Irvine et coll. montraient, en 2002, que les cellules T CD4 mobilisaient les stocks de calcium intracellulaire en réponse à un seul complexe CMH/peptide, décrivant du même fait un rôle du co-récepteur dans cette haute sensibilité (Irvine et al. 2002). En 1998, pour la première fois, il était montré que l’hétérodimérisation du TCR et de son co-récepteur CD8 avec leur ligand physiologique, un monomère de CMH chargé avec le peptide approprié, engendre une cascade de signalisation emmenant à une forte réponse calcique à des niveaux similaires de ceux induits par des anticorps anti-CD3 (Delon et al. 1998).
Ainsi, trois modèles ont été présentés s’appuyant sur le rôle des co-récepteurs et sur la valence des TCR : le modèle d’hétérodimérisation, le modèle de pseudodimérisation et le modèle des TCR multivalents.
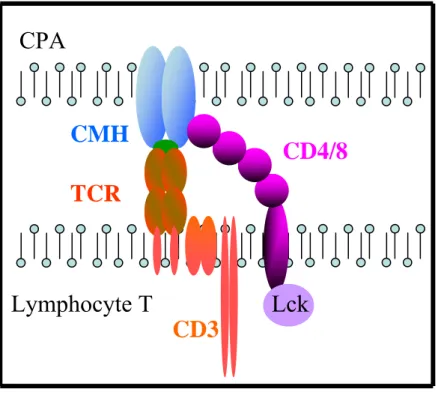
Le modèle d’hétérodimérisation
Le modèle d’hétérodimérisation a été initialement proposé à la fois pour les cellules T CD4+ et CD8+ (Salter et al. 1990) (Madrenas et al. 1996). Mais le support expérimental en faveur de ce modèle a été obtenu avec des cellules T CD8+ (Delon et al. 1998). Ce modèle postule que la signalisation du TCR peut être initiée dans les cellules T CD8+ par l’hétérodimérisation du TCR avec son co-récepteur CD8 associé avec la src-kinase Lck (Lymphocyte-specific protein tyrosine kinase), emmenant ainsi à l’initiation de la transduction de signal. Un tel modèle est en accord avec d’anciens résultats qui montraient que le pontage du CD3 accompagné de celui du co-récepteur (CD4 ou CD8) avait un fort potentiel de stimulation (Anderson et al. 1987). Il a d’ailleurs été remarqué que l’interaction TCR-CMH I est augmentée de façon marquante par le CD8 (Luescher et al. 1995).
Figure 9 : Schéma du modèle d’hétérodimérisation. L’initiation de la signalisation du lymphocyte T s’opère
après hétérodimérisation du TCR et de son co-récepteur (CD4 ou CD8).
Ce modèle s’appuie néanmoins sur un facteur majeur, l’adhérence. En effet, des lymphocytes T adhérents répondent aux complexes CMH/peptide monomériques alors que ces mêmes cellules en suspension ne répondent pas (Randriamampita et al. 2003). Cet effet est observé lorsque les cellules T adhèrent à différents substrats tels que la fibronectine, des anticorps immobilisés et dirigés contre le CMH I, CD11 et CD18 ou encore lorsque ces cellules sont en contact avec des DC (Randriamampita et al. 2003). Ceci explique les résultats apparemment divergents obtenus : tout d’abord le travail initial de Delon et coll. mettait en oeuvre des CTL qui adhéraient à leurs cellules cibles (Delon et al. 1998). De manière parallèle, à la synapse immunologique, un simple comlexe CMH/peptide pouvait engager une réponse calcique de la cellule T en contact avec la CPA (Irvine et al. 2002) ; alors que les lymphocytes qui ne répondaient pas aux complexes monomériques CMH/peptide (Boniface et al. 1998) étaient en suspension.
Il est important de noter que l’adhérence de la cellule T provoque une augmentation dans la quantité de calcium des réservoirs intracellulaires mais aussi dans la quantité de PIP2 (Phosphatidylinositol 4,5-bisphosphate) à la membrane plasmique (Randriamampita et al. 2003). De plus, cette adhérence emmène à la phosphorylation de deux tyrosines kinases,
TCR
CMH
Lymphocyte T
CPA
CD4/8
Lck
CD3
PYK-2 (proline rich tyrosine kinase-2) et Lck ainsi que d’une molécule adaptatrice associée au cytosquelette, la paxilline (Doucey et al. 2003).
Ainsi, le seuil de détection pour la signalisation du TCR est significativement plus bas dans une cellule T « pré-activée » par adhérence et de ce fait sensible au complexe CMH/peptide monomérique.
Le modèle de pseudodimérisation
Le modèle de pseudodimérisation a été proposé dans le but d’expliquer des résultats obtenus sur cellules T CD4+. Récemment, l’équipe de M.M Davis, entre autres, a trouvé un recrutement substantiel de molécules CMH-peptide endogènes à l’aire de contact cellule T-cellule présentatrice d’antigène (Irvine et al. 2002) (Wulfing et al. 2002). Le plus surprenant était que ces molécules étaient conduites à la synapse immunologique par des interactions faibles avec le TCR. Ce phénomène, pris avec certains résultats montrant qu’un simple complexe CMH/peptide agoniste peut initier l’activation de la cellule T (Irvine et al. 2002), a permis la suggestion du modèle de pseudo-dimérisation (Krogsgaard et al. 2005). Le modèle est illustré sur la figure 10.
Figure 10 : Le modèle de pseudo-dimérisation. D’après (Krogsgaard et al. 2005).
S’appuyant sur les données de Li sur la fonction de CD4 à la synapse immunologique (Li et al. 2004), il est proposé dans ce modèle que dans un premier temps, la liaison d’un TCR à un ligand agoniste (TCR1) crée un lieu d’activation assez stable pour le recrutement d’un autre complexe TCR-CD4 (TCR2). Dans un deuxième temps, le TCR2 ainsi recruté se lie à un complexe CMH/peptide endogène (qui jouera le rôle d’un co-agoniste dans cette circonstance) et le pseudo-dimère résultant (TCR1/agoniste/TCR2/peptide endogène/CD4) engagera une cascade de signalisation induisant le recrutement de la proteine tyrosine kinase