Un peu moins de répit pour les cancers du foie ?
Texte intégral
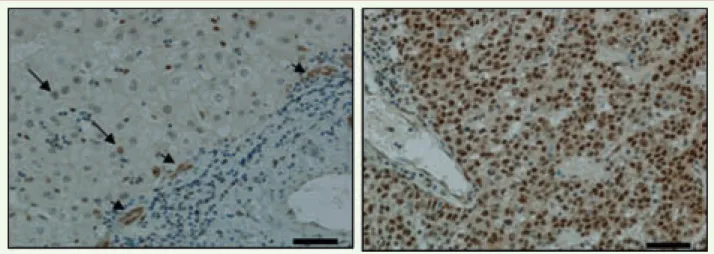
Figure
Documents relatifs
Dans certains cas, le cancer peut être suspecté au cours d’un bilan médical : une analyse de sang ou une imagerie abdominale anor- male peuvent laisser suspecter la présence
A) Influence de la méthylation de l’ADN des îlots CpG dans l’expression des gènes et stratégie anticancéreuse ; B) deux molécules approuvées pour traiter des cancers
Il consiste à appuyer le développement de nouveaux outils de commercialisation en cuircuits courts et à apporter une réponse à l'insécurité alimentaire à travers le projet
© DR et Direction mutualisé e de la communication de Grand Chambéry et de la Ville
C’est dans ce cadre que le ministère chargé de l’environnement lance en mars 2012, dans le cadre de la Stratégie nationale de ges- tion intégrée du trait de côte, un appel
J’ai privilégié ici deux objets d’étude sans existence formelle ex ante, mais dont j’interroge la structuration comme conséquence du développement des circuits courts et
Les intermédiaires de la distribution (détaillants, grossistes) qui participent largement à ces dynamiques sont pourtant souvent négligés tant dans la littérature que dans les
Nous pensons que commerce équi- table et relocalisation de l’économie peuvent être les deux faces d’un même monde plus juste et plus rési- lient. Tout en développant des
