Effets des probiotiques sur le microbiote intestinal de
l’abeille mellifère (Apis mellifera) et sur la performance
des colonies au printemps en climat nordique
Mémoire
Naomie Bleau
Maîtrise en biologie - avec mémoire
Maître ès sciences (M. Sc.)
Effets des probiotiques sur le microbiote intestinal
de l’abeille mellifère (Apis mellifera) et sur la
performance des colonies au printemps en climat
nordique
Mémoire
Naomie Bleau
Sous la direction de :
Pierre Giovenazzo, directeur de recherche Nicolas Derome, co-directeur de recherche
Résumé
Le microbiote intestinal regroupe l’ensemble des micro-organismes qui colonisent l’intestin d’un animal. Chez l’abeille, il participe à la digestion et aux fonctions immunitaires. Plusieurs facteurs environnementaux perturbent le microbiote de cet insecte, nuisant ainsi à sa santé. Pour prévenir cette situation, l’ajout de probiotiques dans l’alimentation des abeilles est une solution intéressante. Il a été montré que certains probiotiques favorisent la croissance des colonies et diminuent l’incidence de plusieurs maladies et parasites. Cependant, aucune information n’est disponible quant à leurs effets en climat nordique, là où la survie hivernale et le développement printanier des colonies sont des enjeux majeurs pour les apiculteurs.
L’objectif principal du projet est d’évaluer l’effet de formules probiotiques sur la composition taxonomique du microbiote de l’abeille et la performance des colonies. Deux souches probiotiques commerciales (Bactocell® et Levucell®SB), une bactérie endogène
(Parasaccharibacter apium) et un antibiotique ont été administrés à un total de 45 colonies d’abeilles à l’automne 2017 et au printemps 2018. Un profilage du microbiote bactérien a été réalisé à quatre moments au cours du projet : en septembre, novembre, avril et juin. La survie hivernale et la performance des colonies ont été évaluées en mesurant le nombre de larves, le nombre d’abeilles dans la colonie, la consommation hivernale de nourriture et le poids des ruches.
Le moment de l’année a influencé la composition du microbiote intestinal de l’abeille, suggérant une adaptation aux conditions environnementales. De plus, la prise de probiotiques n’a modulé que légèrement le microbiote de l’abeille. Quant à la performance des colonies, les formules probiotiques Bactocell® et Levucell®SB ont amélioré le développement printanier des colonies.
Ces résultats indiquent que les probiotiques peuvent améliorer la performance des colonies d’abeilles, et ce, sans perturber leur microbiote. Ces informations seront utiles dans l’optique de développer une formule probiotique destinée à l’industrie apicole canadienne.
Abstract
The microbiota represents microorganisms that inhabit the gut of an animal. The honeybee’s gut microbiota contributes to digestion, immunity and protection of the gut lining. Many environmental factors are known to disrupt the microbial balance of the microbiota and thus affecting honeybee health. The use of probiotics is now considered by many as a sustainable strategy to improve colony health. In fact, studies have shown that some probiotic strains could reduce dysbiosis while enhancing brood development, honey yield and pathogen tolerance. However, none of them investigated the benefits of probiotic supplementation under a cold climate, where winter survival and spring buildup are two critical moments for beekeepers.
The goal of this project is to assess the effects of probiotic formulas on taxonomical composition of the bees’ microbiota and colony performance. To this purpose, forty-five honeybee colonies were given a commercial probiotic strain (Bactocell® or Levucell®), an endogenous bacterium (Parasaccharibacter apium) or the antibiotic Fumagillin-B®. The
treatments were given twice in October 2017 and twice in April 2018. A molecular analysis of the bacterial microbiota was performed to characterize the seasonal changes happening in the honeybee’s gut and the impacts of the treatments on the microbiota’s bacterial community. Colony performance was assessed using brood coverage, number of frames covered in bees and hive weight.
The honeybee microbiota was impacted by the sampling month. We observed a compositional shift and a reduction of the bacterial diversity during the study. Furthermore, the probiotic treatments did not impact significantly the composition of the microbiota. Both Bactocell® and Levucell® favoured the spring buildup of the treated colonies.
Our results indicate that probiotics are useful to improve honeybee colony performance without harming their microbiota. With these results in mind, we aim to develop a safe and effective probiotic formulation specific to the Canadian beekeeping industry.
Table des matières
Résumé ... ii
Abstract ... iii
Table des matières ... iv
Liste des tableaux ... vi
Liste des figures ... vii
Liste des abréviations, sigles, acronymes ... ix
Remerciements ... xi
Introduction ... 1
Chapitre 1 – État des connaissances ... 2
1.1 Statut de l’abeille au Canada ... 2
1.2 État de santé des colonies ... 2
1.2.1 Principales maladies et parasites de l’abeille ... 3
1.2.2 Environnement ... 4
1.2.3 Conditions climatiques ... 5
1.3 Le microbiote intestinal d’Apis mellifera ... 6
1.3.1 Composition et acquisition du microbiote ... 6
1.3.2 Rôle du microbiote et de ses composantes ... 8
1.3.3 La dysbiose ... 10
1.4 Les probiotiques ... 12
1.4.1 Augmentation de la résistance aux agents pathogènes ... 13
1.4.2 Amélioration de la performance des colonies ... 15
1.5 Objectifs et hypothèses ... 16
1.5.1 Objectifs ... 16
1.5.2 Hypothèses ... 17
Chapitre 2 — Étude de l’impact des probiotiques sur le microbiote intestinal de l’abeille et la performance des colonies ... 19
2.1 Matériel et méthodes ... 19
2.1.1 Préparation et gestion des colonies ... 19
2.1.2 Préparation des traitements expérimentaux... 20
2.1.3 Protocole expérimental ... 22
2.1.4 Les traitements ... 23
2.1.5 Échantillonnage ... 23
2.1.6 Performance des colonies et présence des agents pathogènes ... 25
2.1.7 Extraction d’ADN et séquençage ... 28
2.1.8 Analyses bio-informatiques... 30
2.1.9 Analyses statistiques ... 30
2.2 Résultats ... 33
2.2.1 L’influence de l’hiver sur le microbiote intestinal des abeilles ... 33
2.2.2 Effets des probiotiques sur le microbiote intestinal ... 40
2.2.3 Effets des probiotiques sur la performance des colonies ... 42
2.2.4 Taux d’infestation de Varroa destructor ... 46
2.3 Discussion ... 49
2.3.2 Effets des traitements probiotiques sur le microbiote intestinal ... 51
2.3.3 Efficacité des probiotiques sur la performance des colonies ... 53
2.3.4 La diversité des probiotiques... 56
2.3.5 Les défis de l’étude des probiotiques en apiculture ... 58
Conclusion ... 60
Bibliographie ... 63
Liste des tableaux
Tableau 2.1 : Description des traitements administrés aux colonies des groupes expérimentaux et de leur concentration.
Tableau 2.2 : Calendrier des manipulations réalisées sur les colonies entre le mois d’août 2017 et le mois de juin 2018. La couleur bleue indique le moment où a eu lieu la manipulation. La gestion des colonies comprend toutes les manipulations normalement réalisées par les apiculteurs. Les manipulations expérimentales sont propres à ce projet.
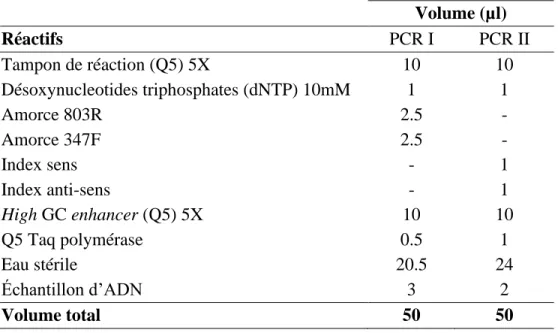
Tableau 2.3 : Volume des réactifs utilisés dans les mélanges pour les réactions PCR I et PCR II.
Tableau 2.4 : Comparaison statistique (test de Kruskal-Wall) de l’abondance relative des taxons bactériens de l’intestin moyen des abeilles entre les quatre temps d’échantillonnage (septembre (T0), novembre (T1), avril (T2) et juin (T3)) pour le groupe CTRL. Les bactéries sont en ordre décroissant d’abondance. Tableau 2.5 : Table d’ANOVA du nombre de cellules de couvain (CO et CNO) en
fonction du groupe en septembre (T0, pré-traitement), mai (une semaine après l’hivernage) et après 4 semaines de développement printanier en juin. Tableau 2.6 : Modèles utilisés pour la sélection de modèle avec l’AIC.
Liste des figures
Figure 1.1 : Pourcentage de mortalité hivernale des colonies d’abeilles au Canada et au Québec de 2004 à 2018 (tirée du document Enquête sur la mortalité hivernale des colonies d’abeilles au Québec en 2017-2018 produit par le MAPAQ) Figure 1.2 : Représentation de l’intestin d’Apis mellifera et composition bactérienne
générale du microbiote (Figure adaptée de Kwong et Moran (2016)) Figure 2.1 : Carte satellite de l’est du Canada. Le point rouge sur la carte indique
l’emplacement du Centre de recherche en sciences animales de Deschambault.
Figure 2.2 : Stades de développement du couvain sur un cadre. Les œufs (1) et les larves (2) forment le couvain non-operculé (CNO). Les nymphes en métamorphose (3) composent le couvain operculé (CO).
Figure 2.3 : Abeilles en grappe vues du dessus de la ruche.
Figure 2.4 : Abondance relative des principaux taxons bactériens composant le microbiote de l’abeille à quatre moments d’échantillonnage. Chaque barre verticale représente le microbiote moyen de toutes les colonies d’un groupe à un moment donné. Seuls les taxons ayant une abondance relative supérieure à 1% sont présentés dans ce graphique.
Figure 2.5 : Graphique de PCoA du microbiote de chaque colonie basé sur l’indice de dissimilarité de Bray-Curtis. Groupe CTRL (A) et tous les groupes expérimentaux (B). Chaque point représente une colonie.
Figure 2.6 : Réseaux des principaux taxons bactériens du groupe CTRL en septembre (T0), novembre (T1), avril (T2) et juin (T3). Les traits verts indiquent une corrélation positive et les rouges, une corrélation négative. Un lien épais indique une forte corrélation alors qu’un lien mince indique une faible corrélation.
Figure 2.7 : Indices de diversité (Shannon, Chao1 et Pielou) pour chaque groupe expérimental et chaque moment d’échantillonnage en septembre (T0), novembre (T1), avril (T2) et juin (T3). Les liens indiquent les différences entre les moments d’échantillonnage pour le groupe CTRL. Les barres d’erreur représentent l’erreur standard. * : p ≤ 0,05; ** : p ≤ 0,01; *** : p ≤ 0.001
Figure 2.8 : Abondance relative des taxons qui diffèrent entre le groupe contrôle et au moins un groupe traité. Échantillonnages en septembre (T0), novembre (T1), avril (T2) et juin (T3). (A) Snodgrassella sp. diffère à CTRL chez les groupes FMG et PB1 à T1. (B) Enterobacteriaceae diffère entre CTRL et PB3 à T2. Les barres d’erreur représentent l’erreur standard. Toutes les comparaisons sont faites avec le groupe CTRL. ■: p ≤ 0,1.
Figure 2.9 : Gain moyen de cellules operculées (CO) et non operculées (CNO) selon le groupe pendant le mois de mai 2018. Les barres d’erreur indiquent l’erreur standard. Les comparaisons sont faites avec le groupe contrôle. * : p < 0,05; ** : p < 0,01
Figure 2.10 : Nombre moyen de cadres recouverts d’abeilles selon le groupe le 9 novembre et le 27 avril. Les barres d’erreur indiquent l’erreur standard.
Figure 2.11: Évolution du poids moyen des groupes pour toute la durée de l’étude. Les barres d’erreur indiquent l’erreur standard. La mesure a été répétée une fois
Figure 2.12 : Indice de consommation hivernale (Ch) moyen selon le groupe expérimental.
L’indice Ch est calculé à l’aide du nombre de cadres d’abeilles en novembre
2017 et de la perte de poids hivernale. Les barres d’erreur indiquent l’erreur standard.
Figure 2.13 : Nombre moyen de varroas tombés par chute naturelle selon le groupe. Les varroas ont été récoltés pendant une semaine et la moyenne quotidienne a ensuite été calculée. La mesure a été répétée une fois par mois. Chaque point représente une colonie.
Figure 2.14 : Nombre moyen de spores de Nosema sp. par abeille. Les mois représentent le moment de l’échantillonnage. Les barres d’erreur indiquent l’erreur standard. Les comparaisons sont faites avec le groupe contrôle. * : p ≤ 0,05.
Liste des abréviations, sigles, acronymes
ACPA: Association canadienne des professionnels de l’apiculture AIC : Critère d’information d’Akaike
ASV : Amplicon Sequence Variants
Ch : Indice de la consommation hivernale de sirop
CO : Couvain operculé CNO : Couvain non-operculé CTRL : Groupe contrôle
FMG : Groupe traité à la Fumagiline-B®
MAPAQ : Ministère de l’Agriculture, des Pêches et de l’Alimentation du Québec MI : Microbiote intestinal
Pb : Paires de bases
PCR : Amplification en chaine par polymérase (Polymerase Chain Reaction) PB1 : Groupe traité avec Parasaccharibacter apium
PB2 : Groupe traité avec Bactocell® PB3 : Groupe traité avec Levucell®SB
SDS : Dodécylsulfate de sodium UFC : Unités formatrices de colonies
À mon père, qui m’a toujours poussé à terminer ce que j’entreprends. Merci.
Remerciements
Je tiens d’abord et avant tout à remercier mon directeur de recherche Pierre Giovenazzo. C’est son enthousiasme contagieux pour l’apiculture qui m’a donné envie de joindre son laboratoire pour réaliser ce projet. Son dévouement et ses encouragements m’ont motivé à poursuivre ma maîtrise quand je ne voyais pas le bout. Merci pour cette belle expérience!
Merci à mon co-directeur Nicolas Derome pour son soutien tout au long de ma maîtrise et pour la confiance qu’il a eue en moi.
Je tiens à remercier chaleureusement toute l’équipe apicole du CRSAD. Merci pour l’aide sur le terrain, mais aussi pour les innombrables fous rires. Travailler avec vous est un réel plaisir! J’espère pouvoir me faire piquer encore de nombreuses fois en votre compagnie.
Merci à Pierre-Luc Mercier, Sarah El-Khoury et Sidky Bouslama du laboratoire Derome pour votre précieuse aide. J’étais particulièrement novice en biologie moléculaire et en bio-informatique et grâce à vous, mon expérience en laboratoire s’est merveilleusement bien passée.
La réalisation de ce projet a été possible grâce à la contribution financière du Projet Apis m., de Lallemand inc. et du CRSAD. Merci du soutien.
Enfin, je tiens à remercier Xavier pour tout, absolument tout. Tu as été là beau temps, mauvais temps, et ce, sans jugement. Ta présence à mes côtés pendant ces deux années d’études m’a fait le plus grand bien.
Introduction
Au Canada, on retrouve environ 800 espèces d’abeilles qui vivent de façon solitaire ou en colonies. Pour sa part, l’abeille domestique, ou Apis mellifera, est un insecte eusocial qui est élevé par l’homme depuis des milliers d’années. De nos jours, l’apiculture est pratiquée principalement dans le but de récolter du miel, mais la pollinisation des cultures par les abeilles est une activité qui prend de l’importance. Ainsi, l’accès à plusieurs ressources alimentaires dépend directement de la présence des abeilles.
Partout dans le monde, l’apiculture fait face à de nombreux défis. L’usage de pesticides en grande quantité, la propagation de plusieurs parasites et l’émergence de maladies contribuent tous à l’augmentation de la mortalité des colonies d’abeilles. Au Canada, l’hiver long et rigoureux est également un élément contraignant pour les apiculteurs et les abeilles. Au cours des dix dernières années, le taux de mortalité hivernal moyen des colonies est d’environ 25%, et on observe des pics à plus de 35%. Pour compenser les pertes hivernales, les apiculteurs ont recours à l’importation d’abeilles au printemps. Cependant, cette mesure n’est pas sans risque : l’importation d’abeilles malades ou ayant une génétique indésirable sont deux possibilités bien présentes.
Dans l’optique d’améliorer la santé des abeilles localement et d’améliorer la survie et la performance des colonies, il est urgent de développer un traitement qui soit efficace, sans danger pour l’abeille et adapté au climat nordique dans lequel nous vivons. Au cours des dernières décennies, l’utilisation de probiotiques s’est avérée efficace tant chez l’humain que chez le bétail, mais peu d’information existe quant aux bénéfices qu’ils peuvent apporter aux abeilles. Ce projet a donc pour but de vérifier l’impact de trois formules probiotiques et d’un antibiotique sur le microbiote intestinal des abeilles et sur la performance printanière des colonies. Les résultats nous permettront de développer une formule probiotique qui soit adaptée à l’apiculture et qui permet d’avoir des colonies plus vigoureuses lors de la pollinisation des cultures.
Chapitre 1 – État des connaissances
1.1 Statut de l’abeille au
Canada
L’abeille mellifère (Apis mellifera) est l’insecte pollinisateur ayant la plus grande valeur commerciale dans le monde (1). En 2016, le miel produit par les 670 000 colonies d’abeilles canadiennes a rapporté aux apiculteurs 176 millions de dollars. L’abeille est également essentielle à la pollinisation de plusieurs cultures. La mise à fruit du bleuet, de la canneberge et de la pomme dépend de la pollinisation, et 90 % de celle-ci est réalisée par l’abeille domestique. La valeur de ce service est estimée à près de deux milliards de dollars par année au Canada (2). L’abeille nous fournit également plusieurs produits uniques tels la cire, la gelée royale, la propolis et le venin, qui sont utilisés entre autres en apithérapie pour leurs vertus antibactériennes (3).
1.2 État de santé des colonies
Bien que l’abeille mellifère soit un insecte domestiqué, le taux de mortalité hivernal des colonies est en augmentation partout sur la planète (4, 5) et le Canada n’y échappe pas. Au cours de l’hiver 2017-2018, les apiculteurs canadiens ont perdu environ le tiers de leurs colonies (6), ce qui représente le pire taux de mortalité observé depuis 2007 (Figure 1.1). Cette situation a plusieurs conséquences pour les apiculteurs : la perte de revenus associés à l’exploitation des ruches, l’augmentation des dépenses nécessaires au renouvellement du cheptel apicole et la réduction du nombre de ruches disponibles pour la pollinisation des cultures. La survie et la santé des colonies dépendent de nombreux facteurs, dont la présence de maladies, la modification des pratiques agriculturales et les conditions climatiques locales.
Figure 1.1 : Pourcentage de mortalité hivernale des colonies d’abeilles au Canada et au Québec de 2004 à 2018 (tirée du document Enquête sur la mortalité hivernale des colonies d’abeilles au Québec en 2017-2018 produit par le MAPAQ)
1.2.1 Principales maladies et parasites de l’abeille
Les abeilles sont susceptibles d’être affaiblies par plusieurs maladies et parasites. Les apiculteurs doivent donc être vigilants et faire des suivis réguliers afin d’éviter la perte de leurs colonies. Le principal parasite de l’abeille mellifère est sans aucun doute l’acarien
Varroa destructor ou varroa (7). Cet ectoparasite est néfaste tant chez l’abeille individuelle
que chez la colonie. Il agit de façon directe en se nourrissant de l’hémolymphe (7) et des matières grasses (8) de l’abeille juvénile (larve, nymphe) et de l’adulte, entraînant ainsi une perte de poids et une diminution de l’espérance de vie (9). De façon indirecte, le varroa est responsable d’une diminution de la réponse immunitaire de l’abeille (10) et il est le vecteur de plusieurs virus (7), comme le virus des ailes déformées qui empêche les abeilles de voler. La croissance des populations de varroas est rapide et, en l’absence de traitement antiparasitaire, la mort de la majorité des colonies survient en moins de trois ans (7).
Le parasite intracellulaire Nosema spp. est une microsporidie qui est l’agent pathogène responsable de la nosémose. Deux espèces causent cette maladie : N. apis et N. ceranae (11). Lorsqu’une abeille ingère une spore de Nosema spp., cette dernière germe dans l’intestin et pénètre dans une cellule épithéliale dans l’intestin moyen. L’accumulation de spores dans ces cellules provoque leur rupture, et les spores sont expulsées dans la lumière intestinale.
Lorsque les nouvelles spores sont évacuées dans l’environnement avec les fèces, l’infection se répand aux autres abeilles de la ruche et ultimement, la maladie est déclarée dans la colonie lorsque l’apiculteur remarque des symptômes cliniques (10). On observe une augmentation de la prévalence du parasite dans les colonies au cours de l’automne et de l’hiver, suivi d’une diminution au printemps lorsque les abeilles effectuent leur vol de propreté (12). La présence du parasite en grande quantité chez plusieurs abeilles peut causer la mort de la colonie en l’absence de traitement (13).
Enfin, la loque américaine et la loque européenne sont deux maladies qui sont causées par les bactéries Paenibacillus larvae et Melissococcus plutonius respectivement (10). Ces bactéries infectent les larves d’abeilles et celles-ci meurent avant d’atteindre le stade adulte ; la colonie atteinte voit donc sa population diminuer rapidement. L’antibiotique utilisé pour contrôler ces deux maladies est l’oxytétracycline, mais certaines souches bactériennes ont développé une résistance à ce produit (14, 15). La loque américaine est la plus dévastatrice des deux maladies puisque P. larvae produit des spores qui restent viables pendant plusieurs décennies dans l’environnement et dans le matériel apicole. Au Québec, il est obligatoire de déclarer la présence de cette maladie au MAPAQ et les ruches concernées doivent être détruites par le feu (16).
1.2.2 Environnement
L’habitat des colonies d’abeilles est un élément déterminant de leur santé. L’intensification des pratiques agricoles, notamment par l’augmentation de la superficie cultivable dominée par les monocultures, réduit la disponibilité et la biodiversité florale dont les abeilles ont besoin pour se nourrir : elles doivent donc parcourir de plus grandes distances pour avoir accès à de la nourriture de qualité (17). De plus, l’utilisation massive de pesticides, tant par les agriculteurs que par les apiculteurs, a des répercussions négatives sur les abeilles. Les néonicotinoïdes ont la mauvaise réputation de nuire aux fonctions reproductives des reines (18) et à l’orientation des ouvrières au vol, en plus d’augmenter la mortalité des abeilles exposées (19). Le déplacement des ruches pour la pollinisation des cultures fruitières ajoute un stress supplémentaire aux abeilles (20).
1.2.3 Conditions climatiques
L’hiver long et rigoureux du Canada est une particularité de l’apiculture nordique. D’octobre à avril, les ouvrières n’ont pas accès à du miel ou à du pollen frais et elles ne peuvent pas sortir de la ruche au risque de mourir de froid. L’hiver, la colonie ne s’endort pas : elle reste active et les ouvrières font vibrer les muscles de leurs ailes pour produire une chaleur pouvant atteindre 43 °C (21). Les abeilles assurent aussi une bonne circulation de l’air dans la ruche sans laquelle l’humidité devient trop importante et la moisissure apparaît. Une colonie doit donc être très populeuse, avoir suffisamment de réserves alimentaires et être en santé afin d’être en mesure d’affronter tous ces défis et de survivre à la saison froide.
Pour remédier aux importantes pertes hivernales, les apiculteurs importent au printemps des reines de la Californie, de la Nouvelle-Zélande et de l’Australie (2). Cette pratique est toutefois risquée, car elle favorise l’entrée accidentelle d’agents pathogènes et de parasites exotiques et d’abeilles ayant une génétique indésirable, comme les abeilles africanisées, qui sont reconnues pour leur agressivité (2). Ainsi, il est préférable de concentrer nos efforts sur la réduction de la mortalité et l’amélioration de la santé des abeilles canadiennes.
Actuellement, pour atteindre cet objectif, des antibiotiques et des acaricides sont utilisés pour diminuer l’incidence des principales maladies et sont appliqués principalement à l’automne. Cependant, cette solution n’est pas sans danger. En effet, les parasites Nosema ssp. et V.
destructor présentent des signes de résistances à ces produits (22, 23). De plus, il a été montré
certains antibiotiques, comme l’oxytétracycline et la fumagiline, ont un effet toxique sur l’abeille (24, 25). L’oxytétracycline nuit également au processus de digestion des protéines (26). Certains apiculteurs vont se tourner vers la sélection génétique pour obtenir des lignées d’abeilles adaptées à leur environnement. Malgré l’efficacité de cette pratique, cela reste une solution à long terme qui ne permet pas de régler les problèmes sanitaires aigus. Il est donc urgent d’agir et de trouver des alternatives de remplacement aux solutions traditionnelles qui soient efficaces et sans danger pour la santé des abeilles.
1.3 Le microbiote intestinal d’Apis mellifera
1.3.1 Composition et acquisition du microbiote
Le microbiote intestinal (MI) regroupe l’ensemble des microorganismes qui vivent dans l’intestin d’un animal (27). Bien qu’on y retrouve des Levures, des Virus et des Eucaryotes, ce sont les Bactéries qui sont dominantes et qui sont les plus étudiées. Dans le MI de l’Abeille domestique, qui comporte environ un milliard de cellules bactériennes (28), 95 % de toutes les bactéries appartiennent seulement à 9 taxons, chacun correspondant à une espèce ou à un groupe d’espèces apparentées (29). Sa simplicité en fait un modèle intéressant à étudier (29).
L’intestin de l’ouvrière est divisé en trois sections distinctes et chacune d’elle abrite différentes communautés bactériennes (Figure 1.2). La partie antérieure se nomme l’intestin moyen et c’est le lieu de la digestion des aliments et de l’absorption des nutriments. En raison de ce rôle, la membrane intestinale à cet endroit est plutôt perméable et elle est la porte d’entrée de nombreux agents pathogènes, tels les virus et les parasites du genre Nosema (30). C’est la portion du tube digestif qui contient le moins de bactéries (environ 107 cellules),
mais on y observe la plus grande diversité (31). Chez l’ouvrière, l’intestin moyen est dominé par Gilliamella apicola, Snodgrassella alvi (32) et Bartonella apis (31). D’autres bactéries, comme Parasaccharibacter apium et Lactobacillus kunkeei, sont souvent présentes et proviennent principalement du pollen et du nectar (31). La bactérie Frischella perrara colonise plutôt le pylore, qui est un petit segment situé tout juste en aval de l’intestin moyen (33). Par la suite, l’illéum est le tube qui relie l’intestin moyen au rectum (30) et il est recouvert d’un important biofilm composé de S. alvi, G. apicola et des bactéries du genre
Lactobacillus Firm-5 (31). Enfin, le rectum est le site de l’absorption de l’eau et de la
formation des fèces (30). C’est l’endroit où les bactéries sont les plus abondantes : on y retrouve environ 109 cellules bactériennes (34). Le rectum est dominé par les
Lactobacillus Firm-4, L. Firm-5 et les Bifidobacterium spp. (31). La composition du MI
diffère également entre les différentes castes, soit les ouvrières, les faux-bourdons et la reine (35). Chez le faux-bourdon, on retrouve plus de deux fois plus de bactéries appartenant au genre L. Firm-5 alors que chez la reine, c’est plutôt P. apium qui domine l’intestin (36).
Figure 1.2 : Représentation de l’intestin d’Apis mellifera et composition bactérienne
L’acquisition du MI débute dès le stade larvaire grâce au contact avec les ouvrières nourrices et la nourriture, mais sa composition n’est que transitoire. Lors de la métamorphose de la larve en abeille adulte, la couche superficielle de la paroi intestinale se détache et est excrétée, libérant ainsi les bactéries l’ayant colonisée (37). L’acquisition du MI mature débute dès l’émergence de l’abeille adulte, lorsqu’elle retire l’opercule de cire de sa cellule. Le contact avec les autres ouvrières par trophallaxie, avec les composantes de la ruche (35) et le contact fécal oral (38) sont nécessaires au développement d’un MI sain. Sa composition est définitive et stable après quatre jours de vie (34).
1.3.2 Rôle du microbiote et de ses composantes
Il existe une relation symbiotique entre le microbiote et son hôte : le premier améliore les fonctions digestives et métaboliques, alors que le second offre aux microorganismes un environnement protégé où vivre, en plus de ressources alimentaires (39). Grâce aux études réalisées sur des abeilles axéniques — qui ne possèdent pas de microbiote — et aux analyses génomiques, il a été possible d’établir comment chaque membre du MI interagit avec l’abeille.
Une des fonctions principales du MI chez tous les animaux est la conversion d’aliments en molécules assimilables par l’hôte. Chez l’abeille, cette fonction est assurée principalement par les taxons G. apicola, Lactobacillus spp. et Bifidobacterium (40, 41). Par exemple, la pectine contenue dans la paroi du pollen est indigeste pour l’abeille, mais G. apicola possède des enzymes qui lui permettent de dégrader cette molécule en une forme assimilable par l’hôte (42), l’acide galacturonique (29). Les bactéries du MI synthétisent aussi des molécules essentielles, comme le font les membres du taxon L. Firm-5, qui possèdent des gènes permettant la biosynthèse du tréhalose, un glucide utilisé comme réserve d’énergie chez les insectes (35).
Les microorganismes composant le MI de l’abeille participent activement à la protection de son hôte de diverses façons. Certains symbiotes, comme la bactérie Acinetobacter sp., freine la croissance de l’agent causal de la loque américaine, P. larvae, lors de tests d’inhibition en
plat de pétri (43). Il a également été montré qu’un débalancement du microbiote normal d’A.
mellifera augmente la susceptibilité de l’insecte à être infecté par le parasite Lotmaria passim
(44).
La couche de mucopolysaccharides qui recouvre le tube digestif protège les cellules épithéliales (41). Bien qu’au sein de ce biofilm on y retrouve une grande diversité de microorganismes, les bactéries lactiques, G. apicola et S. alvi sont celles qui assurent principalement sa formation et son maintien (45, 46). De plus, F. perrara cause l’apparition d’une bande colorée à la surface du pylore (33), phénomène futile en apparence, mais qui serait lié à la stimulation du système immunitaire par cette même bactérie. Sa présence provoque une augmentation de l’expression du peptide antimicrobien apidaecine (47), ce qui peut être bénéfique pour l’abeille en l’aidant à combattre certaines bactéries pathogènes. Enfin, Bartonella apis est une bactérie souvent présente dans l’intestin des ouvrières, mais dont le rôle est incertain. Elle est souvent identifiée comme une bactérie opportuniste (48,
49) qui a le potentiel d’être bénéfique pour l’abeille (34).
Les autres bactéries du MI de l’abeille exercent des fonctions plutôt diversifiées. Par exemple, P. apium possède des gènes qui sont liés à l’oogenèse, ce qui explique possiblement pourquoi elle domine l’intestin de la reine (36). Cette bactérie peut aussi vivre dans la gelée royale alors que c’est un environnement toxique pour la plupart des bactéries (48). Chez certains insectes, les symbiontes de la même famille que P. apium, c’est-à-dire les
Acetobacteraceae, permettent le développement des tissus (50) et contribuent à la maturation
de l’immunité intestinale (51). Pour sa part, Bifidobacterium asteroides stimule la production de prostaglandines et d’hormone juvénile, deux molécules connues pour influencer le développement de l’abeille (52). La présence de la bactérie S. alvi dans l’intestin permet de préserver l’anoxie du milieu, ce qui favorise les activités métaboliques qui craignent la présence de l’oxygène (53).
Bien que chaque espèce ait son importance en tant qu’individu, on ne peut pas uniquement réduire le microbiote à la somme de ses composantes : il faut le considérer comme un tout et observer les interactions qui existent (40). Par exemple, G. apicola et S. alvi semblent
dépendre l’une de l’autre. En effet, ces deux bactéries forment un biofilm qui se superpose l’un sur l’autre dans l’iléum (35). Grâce à cette proximité, l’acétate produit par G. apicola lors de la conversion des glucides complexes est utilisé par S. alvi. Cette molécule lui sert de donneur d’électron lors du maintien de l’anoxie du milieu intestinal (54).
1.3.3 La dysbiose
Le MI d’une abeille doit être à l’équilibre pour réaliser toutes ses fonctions de façon adéquate. Il est en dysbiose lorsqu’il diffère de son état normal et sain, et que les changements observés ont des conséquences sur la santé de l’hôte (40). La dysbiose se manifeste dans l’intestin par un changement dans la proportion relative des taxons, par une augmentation ou une diminution du nombre de microorganismes présents (31) ou par une variation dans le nombre d’interactions positives et négatives entre les bactéries (40). Le débalancement du MI de l’abeille est principalement provoqué par un stress physiologique résultant de variations environnementales, de la présence d’agents pathogènes ou de l’utilisation de médication.
L’environnement dans lequel évoluent les abeilles influence directement la composition de leur MI. Le pollen et le nectar y jouent un grand rôle puisqu’ils nourrissent les abeilles en plus d’être l’unique lien entre certains microorganismes et le tube digestif. À cet effet, on observe une augmentation de la diversité bactérienne entre mai et juillet, probablement en raison de l’augmentation de la diversité florale accessible aux abeilles (32). Une alimentation saine et diversifiée favoriserait donc l’équilibre normal du MI (32). À l’opposé, la consommation de pollen vieilli provoque un débalancement de la flore bactérienne normale : il y a une diminution de S. alvi et une augmentation de F. perrara. Les abeilles subissant ces changements prennent moins de poids que les abeilles nourries avec du pollen frais, en plus d’avoir un taux de mortalité plus élevé que celles-ci (55). Puisque l’état altéré du MI semble provoquer changements physiologiques chez l’abeille, nous pouvons affirmer que l’insecte est bel et bien en dysbiose. Enfin, tout juste après l’hiver, la disponibilité limitée de nourriture s’accompagne d’une diminution de la densité et de la diversité bactérienne dans l’intestin de l’abeille (32). Les conséquences de cette diminution n’ont cependant pas été mesurées.
La présence de certains agents pathogènes et parasites est associée à la dysbiose du MI des abeilles infectées. Dans les ruches parasitées par V. destructor, on observe une augmentation de la proportion de S. alvi et une diminution de Lactobacillus spp. dans l’intestin des ouvrières (56, 57). Contrairement au microbiote des larves saines, qui est largement dominé par les Enterobacteriaceae, le MI des larves parasitées par un varroa est très diversifié et partage plusieurs similitudes avec celui du parasite (58). Les parasites responsables de la nosémose semblent aussi influencer la composition du MI de l’Abeille. L’infection artificielle d’abeilles avec N. ceranae indique que le symbionte G. apicola semble associé avec la présence de cette Microsporidie (59). Cette observation n’a toutefois pas été observée chez les abeilles naturellement parasitées (56). De plus, en présence de Nosema apis, on observe simultanément une augmentation de P. larvae, la bactérie responsable de la loque américaine (56). Il est important de noter que les mécanismes associés aux changements observés dans le MI en présence d’agents pathogènes ne sont toujours pas connus.
Enfin, l’utilisation d’antibiotiques et de pesticides favorise également l’état de dysbiose. Lors d’un essai réalisé avec la tétracycline, antibiotique utilisé pour combattre la Loque américaine (Paenibacillus larvae), le MI des abeilles traitées comportait environ trois fois moins de bactéries que les abeilles du groupe témoin. Les chercheurs ont aussi observé une diminution importante de S. alvi et de Bifidobacterium sp., deux bactéries essentielles aux abeilles. Une semaine après le traitement, les effets persistaient (60). De plus, chez les abeilles infectées et traitées avec de la pénicilline et de la streptomycine, on dénombre davantage de spores de
Nosema spp. que chez les abeilles qui ne sont pas traitées, ce qui suggère que la dysbiose
causée par les antibiotiques nuit aux mécanismes de défense immunitaire des abeilles (61). Dans le cas de la Fumagiline-B, antibiotique utilisé pour traiter la nosémose, ses effets sur la santé des abeilles ont été étudiés, mais l’impact de cet antibiotique sur l’intégrité du MI des abeilles reste à ce jour inconnu.
Étant donné le nombre important de facteurs pouvant nuire à l’intégrité du MI des abeilles mellifères et aux impacts que ces changements peuvent avoir sur leur santé, il est essentiel de trouver des solutions de remplacement aux traitements actuels qui favoriseraient la santé des colonies et minimiseraient la dysbiose intestinale.
1.4 Les probiotiques
Les probiotiques sont des microorganismes vivants qui, lorsqu’ils sont administrés en quantité suffisante, procurent des effets bénéfiques sur la santé (62). Les probiotiques agissent principalement de trois façons sur l’hôte qui les consomme, et le mode d’action de chaque microorganisme dépend de son métabolisme et des molécules qu’il sécrète (63).
Premièrement, les probiotiques stimulent le système immunitaire de l’hôte simplement par leur présence. Leur détection par les cellules épithéliales déclenche une réponse immunitaire (63). De plus, ils permettent de conserver l’intégrité de la barrière muqueuse de l’intestin, par exemple en augmentant l’expression des gènes impliqués dans la structure des jonctions serrées (64). Ce mécanisme permet de limiter l’entrée de microorganismes pathogènes dans les cellules épithéliales. Deuxièmement, ils rivalisent directement avec les autres microorganismes présents dans l’intestin de l’hôte. Les probiotiques produisent des substances antimicrobiennes, principalement les acides gras à courte chaîne comme l’acide lactique, ou encore des antibiotiques (63). Par leur présence, les probiotiques entrent en compétition avec les bactéries du MI pour les ressources limitantes comme le fer (65). Les probiotiques peuvent également inhiber les toxines sécrétées par les bactéries pathogènes. Par exemple, Saccharomyces cerevisiae boulardii sécrète une protéase qui est capable de détruire la toxine A sécrétée par Clostridium difficile , agent causal de graves diarrhées chez l’humain (66). Enfin, les probiotiques sont bénéfiques en agissant sur les molécules et les aliments présents dans l’environnement intestinal. Leur conversion dans une forme assimilable par l’hôte est une fonction bien connue de plusieurs bactéries probiotiques. En plus des principaux mécanismes d’action présentés, les probiotiques sont connus pour moduler la composition du microbiote, prévenant et traitant ainsi la dysbiose associée à un état de santé altéré chez l’humain (66).
Les probiotiques sont couramment utilisés par l’humain, soit en consommant des aliments fermentés, soit en prenant des suppléments alimentaires. Ils sont utilisés principalement pour prévenir la diarrhée d’origine bactérienne (67). Ils sont aussi ajoutés dans l’alimentation des
porcs, des poulets et des vaches d’élevage pour stabiliser le pH du rumen des ruminants (68) et stimuler la croissance et la réponse immunitaire des animaux (69). Les principaux microorganismes administrés en tant que probiotiques sont les bactéries du genre
Lactobacillus, Bifidobacterium et Bacillus (70).
En apiculture, les probiotiques ne sont pas encore utilisés couramment, mais il semble de plus en plus évident que leur ajout dans l’alimentation des abeilles a le potentiel d’améliorer la santé des colonies. Un bon candidat probiotique pour le domaine apicole serait idéalement isolé de l’intestin de l’abeille ou de la ruche et cultivable en laboratoire (71) pour être en mesure de reproduire le microorganisme en grande quantité. Il devrait aussi posséder des propriétés connues lui conférant le titre de probiotique, par exemple en produisant des métabolites bénéfiques ou en étant antagoniste à un agent pathogène (71). En plus du choix du probiotique, l’efficacité d’un traitement de cette nature dépend de plusieurs facteurs, comme la fréquence de leur administration ou encore leur concentration. Le mode d’administration est aussi important puisqu’il détermine comment le candidat probiotique sera distribué dans la ruche. En apiculture, on peut donner des probiotiques aux abeilles en les ajoutant à du pollen, à du sirop ou en les vaporisant dans la ruche. Tous ces éléments sont essentiels à considérer pour développer une formule probiotique qui est efficace ; autrement, elle peut être inutile (72) ou même néfaste pour les abeilles (73).
1.4.1 Augmentation de la résistance aux agents pathogènes
Plusieurs expériences réalisées in vitro, en cagette et sur des colonies indiquent que la prise de bactéries bénéfiques permet aux abeilles d’être plus résistantes aux agents pathogènes les plus communs, comme les parasites du genre Nosema et les bactéries responsables de la loque européenne et américaine. L’équipe de Corby-Harris (2016) a infecté artificiellement des abeilles avec 10 000 spores de Nosema spp., puis a supplémenté la moitié des insectes avec
P. apium. Dix jours plus tard, malgré une augmentation du nombre de spores dans l’intestin
de tous les individus, les abeilles ayant consommé P. apium étaient l’hôte de deux fois moins de parasites que celles du groupe témoin (74). Lors d’un essai similaire réalisé avec d’autres microorganismes, l’équipe de Ptaszynska (2016) a noté l’effet inverse. En effet, ces
chercheurs ont utilisé des formules probiotiques destinées à la consommation humaine qui étaient composées de Lactobacillus spp., Bifidobacterium bifidum et Saccharomyces
cerevisiae. Après trois semaines de supplémentation ad libitum, les abeilles ayant consommé
les probiotiques présentaient deux fois plus de spores de Nosema dans leur intestin (73). Cet exemple illustre bien le fait que deux formules probiotiques n’auront pas le même effet en fonction des microorganismes qu’elles contiennent. Dans une troisième étude, des abeilles inoculées avec un million de spores de Nosema ceranae avaient un accès illimité à différents candidats probiotiques endogènes, soit P. apium et Bacillus subtilis, ainsi qu’à des formules probiotiques commerciales Bactocell® (Pediococcus acidilactici) et Levucell® (Saccharomyces cerevisiae) pendant un mois. Toutes les abeilles traitées avec les probiotiques avaient un meilleur taux de survie comparativement aux abeilles du groupe contrôle. Contrairement à l’étude de Corby-Harris, où la prévalence initiale était cent fois plus élevée, le nombre de spores de N. ceranae n’était pas plus faible chez les abeilles traitées à la fin de l’expérience (75) : les insectes étaient possiblement plus tolérants à la présence du parasite. Les effets des probiotiques sur Nosema spp. peuvent aussi être observés sur des colonies en production. En effet, cinq mois après le début l’administration mensuelle d’un litre de solution contenant Bacillus subtilis à raison de 105 bactéries/ml, le nombre de spores détectées chez les colonies traitées était près de quatre fois moindre que celui des colonies du groupe témoin (76). À la lumière de ces résultats, l’utilisation de traitements à base de microorganismes vivants est possiblement une excellente alternative à l’unique traitement disponible, l’antibiotique Fumagilin-B®.
Plusieurs bactéries semblent avoir le potentiel de prévenir et de traiter des maladies bactériennes de l’abeille. Lors d’essais in vitro en plat de pétri, plusieurs bactéries appartenant au genre Lactobacillus et aux Bifidobacterium ont montré un effet inhibiteur sur la croissance de P. larvae (77-79) et de M. plutonius (46), agents bactériens responsables de la loque américaine et européenne. Dans le même ordre d’idée, lorsque des abeilles étaient supplémentées avec les bactéries Bacillus licheniformis et Lactobacillus kunkeei alors qu’elles étaient infectées artificiellement par la loque américaine (P. larvae), leur taux de mortalité était beaucoup plus bas que celui des abeilles non traitées (80). D’autres recherches
devront être menées afin de vérifier si les effets observés en laboratoire peuvent être transposés sur des colonies en production.
1.4.2 Amélioration de la performance des colonies
L’ajout de probiotiques dans l’alimentation des abeilles peut également améliorer la performance des colonies ; l’augmentation de la quantité de couvain et la hausse de la production de miel sont deux effets souvent constatés (76, 81, 82). Par exemple, la supplémentation pendant un mois avec des bactéries des genres Lactobacillus et
Bifidobacterium, administrées sous forme de sirop ou vaporisées directement dans la ruche,
augmente le nombre de cellules occupées par du couvain (stades juvéniles de l’abeille), ainsi que la production de miel d’environ 50 % comparativement aux colonies n’ayant pas eu accès aux probiotiques (83, 84). En laboratoire, on observe une meilleure survie des abeilles qui ont consommé des probiotiques (75, 85), en plus d’un gain de poids plus important (86, 87).
En plus de leur impact sur la santé de l’abeille et les performances de la colonie, la prise de microorganismes bénéfiques peut modifier la physiologie de l’insecte. En effet, la prise de formules probiotiques commerciales destinées à l’humain (Enterobiotics® et Enterolactis®
Plus), à base de Lactobacillus et de Bifidobacterium, augmente significativement la taille des glandes cirières des ouvrières après trois semaines (88). Ce changement a le potentiel d’augmenter la quantité de cire produite, facteur important dans le développement des colonies d’abeilles.
Très peu d’études ont porté sur la prise de probiotiques dans un contexte hivernal, malgré le fait que cette période soit critique pour les colonies. La seule étude à ce sujet a utilisé du pollen enrichi avec la bactérie Parasaccharibacter apium et ils ont observé un léger effet positif sur la survie hivernale (74). Toutefois, cette étude a été réalisée en Arizona, là où la température hivernale moyenne est de 5 °C (89). Étant donné l’efficacité de certains probiotiques sur la tolérance aux pathogènes et la performance des colonies, il est plausible de croire qu’ils puissent améliorer la survie hivernale et le développement printanier des colonies qui vient dans un climat nordique, comme le Québec.
1.5 Objectifs et hypothèses
1.5.1 Objectifs
L’objectif général de ce projet est d’évaluer les effets de l’administration de trois probiotiques et d’un antibiotique sur la santé et le microbiote des colonies d’abeilles dans un environnement typique du Nord-est canadien. Les objectifs spécifiques sont :
- D’étudier l’évolution de la diversité et de l’abondance relative du microbiote de l’intestin moyen des abeilles durant un hiver typique du Nord-est canadien
- D’évaluer l’effet des traitements probiotiques d’automne et d’un traitement antibiotique sur :
o la composition et la structure du microbiote intestinal des abeilles ; o la survie hivernale des colonies ;
o le développement printanier (poids, population d’abeilles juvéniles et adultes) des colonies ;
o sur la présence des parasites Varroa destructor et Nosema spp. ;
Dans le cadre de mon travail, une bactérie endogène (Parasaccharibacter apium), deux formules probiotiques commerciales (Bactocell® et Levucell®, Lallemand inc.) et un
antibiotique (Fumagilin-B®) seront administrés aux colonies d’abeilles. Voici les
caractéristiques principales des traitements :
- Parasaccharibacter apium est une bactérie isolée de l’intestin des abeilles. On la retrouve principalement chez les larves, chez la reine et dans les réserves de nourriture, mais également dans le tube digestif des ouvrières (35). En laboratoire, lorsqu’elle est administrée comme probiotique, elle améliore la survie des larves d’abeilles en développement (48) et la résistance au pathogène N. ceranae (74, 75). Elle semble améliorer légèrement la survie hivernale des colonies d’abeilles (74).
- Bactocell® est une formule probiotique contenant la bactérie lactique Pediococcus
acidilactici (MA18/5M). Elle est actuellement utilisée comme additif dans
l’alimentation des animaux d’élevage monogastriques (porcs et poulets) afin d’augmenter leur croissance et inhiber le développement de certains agents pathogènes (90, 91). Chez les porcelets, l’ingestion de cette bactérie provoque un gain
de poids et l’épaississement de la muqueuse intestinale (92). On retrouve naturellement P. acidilactici dans le pollen récolté par les abeilles (93).
- Contrairement à la majorité des formules probiotiques qui contiennent des bactéries, Levucell® contient la levure Saccharomyces cerevisiae boulardii. Cette levure est
efficace chez l’humain pour traiter les diarrhées (94). Elle est également utilisée dans la production aviaire et porcine, notamment pour diminuer la présence de Salmonella
spp. chez le poulet de chair (95). Les levures du genre Saccharomyces représentent
presque la totalité des levures qui vivent dans l’intestin des jeunes abeilles (96). ). Toutefois, il est important de mentionner qu’il a été montré qu’une grande consommation de cette levure pouvait endommager les cellules épithéliales de l’intestin moyen (97).
- L’antibiotique Fumagilin-B® est utilisé en apiculture pour lutter contre les parasites
Nosema apis et Nosema ceranae (98). C’est le seul traitement autorisé à ce jour pour
traiter la nosémose. Les ingrédients actifs du médicament sont la fumagiline et le dicyclohexylamine (99). L’antibiotique est administré aux colonies à l’automne pour diminuer la présence du parasite pendant l’hiver. Dans le cadre du projet, ce traitement nous sert de témoin pour vérifier l’effet des probiotiques sur la présence des parasites du genre Nosema.
1.5.2 Hypothèses
La réalisation de cette étude nous permet de vérifier quatre hypothèses (H) et leur prédictions (P) associées.
Impact de l’hiver sur la composition et la structure du microbiote de l’abeille
H1: Le moment de l’échantillonnage des abeilles influence la composition et la structure de son microbiote
P1.1 : La diversité microbienne diminue pendant l’hiver et augmente au printemps. P1.2 : L’abondance des Enterobacteriaceae augmente pendant l’hiver, ce qui suggèrerait un état de dysbiose.
Influence des traitements sur le microbiote intestinal des abeilles
H2 : Les probiotiques permettent de restaurer l’intégrité du MI de l’abeille sans être nocif pour celui-ci.
P2 : Les groupes recevant des traitements probiotiques auront une diversité bactérienne stable pendant l’hiver.
H3 : L’antibiotique Fumagiline-B® influence la diversité bactérienne du microbiote intestinal de l’abeille.
P3 : La diversité microbienne diminue après la prise de l’antibiotique à l’automne.
Effets des traitements sur la performance des colonies d’abeilles
H4 : L’ajout de probiotiques dans l’alimentation des abeilles a un effet sur la survie hivernale et la performance printanière des colonies.
P4 : Les colonies recevant les probiotiques auront un meilleur taux de survie hivernal et seront plus performantes au printemps.
Chapitre 2 — Étude de l’impact des probiotiques sur le
microbiote intestinal de l’abeille et la performance des colonies
2.1
Matériel et méthodes
2.1.1 Préparation et gestion des colonies
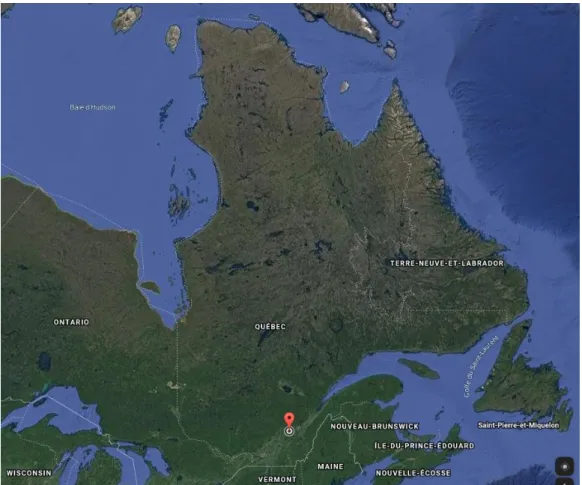
Au début de l’été 2018, quarante-cinq colonies d’abeilles (Apis mellifera) ont été préparées à partir de reine-sœurs issues de la même lignée génétique provenant du cheptel de notre centre de recherche (Figure 2.1, Centre de recherche en sciences animales de Deschambault; 46° 40’30.0’’ N, 71° 54’52.3’’ O). Lorsqu’elles ont atteint la maturité, elles ont été placées à Neuville (Québec), dans deux ruchers situés à environ 6 kilomètres l’un de l’autre : 23 ruches au rucher A et 22 ruches au rucher B.
Figure 2.1 : Carte satellite de l’est du Canada. Le point rouge sur la carte indique
La phase expérimentale du projet s’est déroulée entre septembre 2017 et juin 2018. À l’automne, toutes les colonies ont été traitées contre l’acarien avec deux produits antiparasitaires. D’abord, une bandelette de Thymovar® a été placée entre la hausse à couvain
et l’entre-couvercle pendant 21 jours. Ce traitement a été effectué avant le début de l’étude. Puis, au début du mois de novembre, une solution de 35 g d’acide oxalique dihydrate mélangé à du sirop de saccharose 1 :1 a été administrée par égouttement dans la ruche, pour un total de 5 ml entre chaque cadre. Les colonies ont été placées dans une salle environnementale (4 °C et 45 % HR) pour toute la durée de la période hivernale, soit du 22 novembre 2017 au 20 avril 2018. Au printemps, les colonies vivantes ont été sorties de l’hivernage et placées à Neuville dans deux ruchers : 22 ruches au rucher C et 22 au rucher D.
2.1.2 Préparation des traitements expérimentaux
Le traitement probiotique à base de Parasaccharibacter apium n’étant toujours pas commercialisé, les solutions bactériennes utilisées dans ce projet ont été préparées en laboratoire. Cette bactérie a d’abord été isolée de l’intestin d’ouvrières en santé et a été conservée dans une solution de glycérol à 15% à -80 °C jusqu’à son utilisation (75). Pour démarrer la croissance bactérienne, une portion du milieu glycérol a été inoculée à l’aide d’un fil à boucle en condition stérile, puis étalée sur un pétri contenant du milieu Sabouraud solide (Annexe 1). Le pétri a été incubé à 37 °C pendant 48 h pour permettre la croissance de colonies bactériennes. Par la suite, 5-6 colonies ont été repiquées dans 10 mL de milieu Sabouraud liquide, puis celui-ci a été placé à 37 °C sur un agitateur rotatif pendant 48 h. Ensuite, la totalité du bouillon a été transférée dans 1 L de milieu Sabouraud liquide frais, puis placée à nouveau à 37 °C sur un agitateur.
Afin de déterminer la concentration de cellules bactériennes de la solution probiotique, nous avons mesuré la densité optique (DO) à l’aide d’un spectrophotomètre (Spectronic 700, Bausch & Lomb) à 600 nm. Cependant, pour être en mesure d’interpréter la lecture de l’appareil, il est essentiel d’établir une courbe standard de la croissance de la bactérie d’intérêt. Pour ce faire, nous avons préparé 10 mL de bouillon de culture de P. apium. Ensuite, quatre tubes contenant des dilutions 1/2 , 1/4 ,1/8 et 1/16 de ce bouillon ont été préparés,
en plus d’un cinquième tube contenant du bouillon non dilué. La DO des cinq dilutions a été mesurée à 600 nm avec le spectrophotomètre.
En parallèle, pour connaître le nombre d’unités formatrices de colonies (UFC) associées à la DO, nous avons fait des dilutions 10-3, 10-4, 10-5, 10-6 et 10-7 des cinq solutions préparées précédemment, puis nous avons étalé en duplicata 0.1 mL de chaque dilution sur des pétris contenant du milieu Sabouraud solide. Après 48 h d’incubation à 37 °C, nous avons sélectionné les géloses ayant entre 30 et 300 colonies, nous les avons dénombrées, et nous avons fait la moyenne des duplicata. Enfin, pour obtenir la concentration (UFC/ml) des bouillons préparés, nous avons utilisé la formule suivante :
𝐶𝑜𝑛𝑐𝑒𝑛𝑡𝑟𝑎𝑡𝑖𝑜𝑛 (𝑈𝐹𝐶 𝑚𝑙⁄ ) = 𝑁𝑜𝑚𝑏𝑟𝑒 𝑚𝑜𝑦𝑒𝑛 𝑑𝑒 𝑐𝑜𝑙𝑜𝑛𝑖𝑒𝑠 0,1 𝑚𝑙 ∗ 𝐹𝑎𝑐𝑡𝑒𝑢𝑟 𝑑𝑒 𝑑𝑖𝑙𝑢𝑡𝑖𝑜𝑛
En connaissant la concentration des solutions et leur DO, nous avons établi l’équation de la courbe standard suivante:
𝑦 = 5 × 10−8𝑥 + 0,0582
où y est la DO et x est la concentration de la solution (UFC/mL).
À l’aide du spectrophotomètre et de la courbe standard, nous avons calculé la concentration bactérienne du milieu de culture quotidiennement. Quand le bouillon a atteint la concentration désirée, il a été transféré dans des bouteilles de 250 mL, puis centrifugé à 4000 G pendant 20 minutes afin de faire précipiter les bactéries. À l’aide d’une pipette, le surnageant a été retiré des bouteilles et le culot a été dilué dans environ 200 mL de sirop de saccharose 1 :1 (p:p). La solution a été conservée à 4 °C jusqu’à son administration.
Les probiotiques commerciaux Bactocell® et Levucell®SB (Lallemand inc.) sont disponibles sous forme de poudre. La formule Bactocell® a une concentration de 1010 UFC/g de produit, alors nous avons ajouté 0,1 g à un litre de sirop 1 :1 pour atteindre une concentration de 109 UFC/L. Pour Levucell®, le produit est deux fois plus concentré, alors nous avons ajouté 0,05 g à la même quantité de sirop pour obtenir la concentration 109 UFC/L.
L’antibiotique Fumagilin-B® a été administré selon ce qui est prescrit sur l’étiquette : 9,08 g
du produit ont été mélangés à un litre de sirop de saccharose 1 :1. Cette dose traite la nosémose dans une colonie ayant une hausse à couvain. Les trois solutions probiotiques (P.
apium, Bactocell® et Levucell®) ainsi que le traitement antibiotique ont été préparés la veille de leur administration aux abeilles (75).
2.1.3 Protocole expérimental
Le protocole expérimental comprend cinq groupes expérimentaux comportant chacun neuf colonies, pour un total de quarante-cinq colonies (Tableau 1.1). Un échantillonnage stratifié aléatoire qui tenait compte de la force des ruches et de leur emplacement géographique a été utilisé pour faire la distribution aléatoire des colonies dans les différents groupes expérimentaux. Ainsi, toutes les colonies ont été évaluées (section 2.6) avant le début de l’étude et par la suite, nous les avons classées selon leur force. Cette information nous a permis de former les groupes de colonies de force similaire. Nous nous sommes aussi assurés que les groupes soient représentés équitablement dans les deux ruchers. Les traitements ont été attribués aux groupes de façon aléatoire.
Tableau 2.1 : Description des traitements administrés aux colonies des groupes
expérimentaux et de leur concentration.
Groupe N Traitement Concentration
CTRL 9 Contrôle (sirop de sucre 1 :1) NA
FMG 9 Fumagilin-B® 9,08 g/L
PB1 9 Parasaccharibacter apium 1 x 109 UFC/L
PB2 9 Bactocell® (Lallemand inc.) 1 x 109 UFC/L PB3 9 Levucell® (Lallemand inc.) 1 x 109 UFC/L
2.1.4 Les traitements
À l’automne et au printemps, toutes les ruches ont reçu deux doses de traitement à une semaine d’intervalle, pour un total de quatre doses (Tableau 1.2). Chaque dose consistait en un litre de sirop de saccharose 1 :1 additionné du traitement probiotique ou antibiotique. Le groupe témoin CTRL n’a reçu que le sirop de saccharose. Les colonies du groupe FMG ont reçu la Fumagiline-B une seule fois à l’automne, dans la première dose, comme le recommande le fabricant. Pour les trois autres doses, FMG a reçu du sirop de saccharose, comme le groupe CTRL. Pour les groupes PB1, PB2 et PB3, les colonies ont reçu les différents probiotiques en solution dans un litre de sirop de saccharose 1 :1. Les traitements ont été administrés aux colonies à l’aide d’un nourrisseur de style ‘Miller’ (Propolis-etc.) placé sur le dessus de la ruche. De cette façon, le seul accès au traitement se trouvait à l’intérieur de la ruche, ce qui évitait le pillage par les abeilles des autres colonies, phénomène fréquent au moment des traitements en raison de la faible disponibilité de nourriture. Toutes les colonies ont consommé l’entièreté du sirop et ce, pour tous les traitements.
2.1.5 Échantillonnage
Nous avons échantillonné des abeilles à quatre moments pendant l’expérience : au début du projet en septembre (T0), après l’administration des deux premières doses en novembre (T1), avant les traitements du mois d’avril (T2) et à la fin du projet en juin (T3). À chaque fois, environ 100 abeilles ont été prélevées sur des cadres de couvain et placées immédiatement dans la glace sèche. Les échantillons ont été conservés à -80 °C jusqu’à leur utilisation.
Tableau 2.2 : Calendrier des manipulations réalisées sur les colonies entre le mois d’août 2017 et le mois de juin 2018. La couleur bleue
indique le moment où a eu lieu la manipulation. La gestion des colonies comprend toutes les manipulations normalement réalisées par les apiculteurs. Les manipulations expérimentales sont propres à ce projet.
2017 2018
Mois Août Septembre Octobre Novembre Avril Mai
Semaine 28 4 11 18 25 2 9 16 23 30 6 13 20 16 23 30 7 14 21 28
Gestion des colonies
Évaluation chute naturelle de V.destructor Traitement au Thymovar©
Traitement à l'acide oxalique Entrée des ruches au caveau
Sortie des ruches
Manipulations expérimentales
Administration des traitements
Pesée des ruches
Surface de couvain
2.1.6 Performance des colonies et présence des agents pathogènes
Afin de déterminer l’impact des traitements sur la performance des colonies ainsi que l’importance de la présence de deux agents pathogènes (V. destructor et Nosema spp.), plusieurs critères de performance et paramètres ont été mesurés (Tableau 1.2).
Le nombre d’alvéoles contenant du couvain a été mesuré pour déterminer le stade de développement des colonies. La superficie recouverte par du couvain est mesurée visuellement, puis l’aire est convertie en nombre de cellules en considérant le nombre de cellules par pouce carré et la forme hexagonale des alvéoles (100). Les cellules de couvain sont divisées en deux catégories : celles qui contiennent des œufs et des larves (cellules non operculées, CNO) et celles qui contiennent des pupes en métamorphose (cellules operculées, CO) (Figure 1.3). Cette différenciation nous permet de préciser le stade de développement de la colonie. À l’aide de ces mesures, nous avons calculé le gain brut de couvain, c’est-à-dire la différence entre le nombre de cellules de couvain au début et à la fin du printemps. La superficie recouverte de couvain a été évaluée au début de l’étude pour créer des groupes de force similaire, puis deux fois au printemps, soit une semaine et un mois après l’administration des traitements.
𝑁𝑜𝑚𝑏𝑟𝑒 𝑑𝑒 𝑐𝑒𝑙𝑙𝑢𝑙𝑒𝑠 = 𝑆𝑢𝑟𝑓𝑎𝑐𝑒 (𝑝𝑜2) ∗ 25 ∗ 0.8
Le nombre de cadres recouverts d’abeilles nous informe quant à la taille de la population d’abeilles. Elle est prise au mois de novembre et au mois d’avril lorsque les températures quotidiennes sont basses et que les abeilles sont collées les unes aux autres (Figure 1.4). Le nombre de cadres recouverts d’abeilles sur le dessus et le dessous de la ruche est noté, puis la moyenne est calculée pour chaque ruche. C’est une mesure qui minimise la perturbation du nid lorsqu’il fait froid et qui est utilisée par les apiculteurs pour estimer les pertes hivernales.
Figure 2.2 : Stades de développement du couvain sur un cadre. Les œufs (1) et les larves
(2) forment le couvain non-operculé (CNO). Les nymphes en métamorphose (3) composent le couvain operculé (CO).
Figure 2.3 : Abeilles en grappe vues du dessus de la ruche.
1
2
Le poids des ruches nous informe sur la taille de la population d’abeilles et de l’abondance de réserves alimentaires (pollen et miel). Les ruches ont été pesées une fois par mois pendant toute la durée du projet, à l’exception de la période d’hivernage, entre décembre et avril. La pesée était faite avec une balance qui se place sous la ruche. Cette mesure nous a permis de calculer le gain de poids qui correspond à la différence de poids entre les pesées.
L’indice de consommation hivernale des réserves (Ch) nous indique la quantité de réserves
glucidiques qu’une colonie consomme pendant l’hiver en fonction de sa population. Un faible Ch réduit les chances que la colonie meurt de faim avant l’arrivée du printemps. Cet indice
est calculé à partir du poids des colonies ainsi que la population d’abeilles avant l’hivernage :
𝐶ℎ =
𝑃𝑜𝑖𝑑𝑠𝑛𝑜𝑣𝑒𝑚𝑏𝑟𝑒 − 𝑃𝑜𝑖𝑑𝑠𝑎𝑣𝑟𝑖𝑙 𝐶𝑎𝑑𝑟𝑒𝑠 𝑑′𝑎𝑏𝑒𝑖𝑙𝑙𝑒𝑠𝑛𝑜𝑣𝑒𝑚𝑏𝑟𝑒
Taux d’infestation de V. destructor : Chaque mois, l’infestation par l’acarien V. destructor a été estimée à l’aide de cartons collants placés sous les ruches. Pendant une semaine, les acariens tombaient au fond de la ruche et s’accumulaient sur le carton. Ainsi, nous avons estimé le nombre moyen de varroas tombés quotidiennement.
Le niveau d’infestation par Nosema spp. a été mesuré en dénombrant les spores présentes dans l’intestin des ouvrières selon la méthode décrite par Cantwell (1970). D’abord, les abdomens de trente-cinq abeilles ont été broyés ensemble dans 35 mL d’eau à l’aide d’un pilon et d’un mortier. Par la suite, une goutte de 10 µl a été placée dans chacune des chambres d’un hématocytomètre. Les spores ont été dénombrées à l’aide d’un microscope optique à un grossissement de 400x. Les spores ont été comptées dans cinq carrés par chambre, et ce, dans quatre chambres. La moyenne par carré a ensuite été calculée. Pour déterminer le nombre de spores moyen par abeille, l’équation suivante a été utilisée :
𝑁𝑜𝑚𝑏𝑟𝑒 𝑑𝑒 𝑠𝑝𝑜𝑟𝑒 𝑝𝑎𝑟 𝑎𝑏𝑒𝑖𝑙𝑙𝑒 = 𝑁𝑜𝑚𝑏𝑟𝑒 𝑚𝑜𝑦𝑒𝑛 𝑑𝑒 𝑠𝑝𝑜𝑟𝑒 𝑝𝑎𝑟 𝑐𝑎𝑟𝑟é
80 × (4 × 10