O
pen
A
rchive
T
OULOUSE
A
rchive
O
uverte (
OATAO
)
OATAO is an open access repository that collects the work of Toulouse researchers and makes it freely available over the web where possible.
This is an author-deposited version published in : http://oatao.univ-toulouse.fr/
Eprints ID : 17453
To cite this version :
Semat, Julie. Mise en place d’une méthode de diagnostic
moléculaire par PCR quantitative pour détecter et identifier les microfilaires sanguines de Dirofilaria immitis et de Dirofilaria repens chez le chien. Thèse d'exercice, Médecine vétérinaire, Ecole
Nationale Vétérinaire de Toulouse - ENVT, 2016, 180 p.
Any correspondence concerning this service should be sent to the repository administrator: [email protected].
ANNEE 2016 THESE : 2016 – TOU 3 – 4092
MISE EN PLACE D’UNE MÉTHODE DE
DIAGNOSTIC MOLÉCULAIRE PAR PCR
QUANTITATIVE POUR DÉTECTER ET IDENTIFIER
LES MICROFILAIRES SANGUINES DE
DIROFILARIA IMMITIS ET DE DIROFILARIA
REPENS CHEZ LE CHIEN
_________________
THESEpour obtenir le grade de DOCTEUR VETERINAIRE
DIPLOME D’ETAT
présentée et soutenue publiquement devant l’Université Paul-Sabatier de Toulouse
par
SEMAT, Julie
Née, le 23/04/1992 à Thiais (94)___________
Directeur de thèse : M. Philippe JACQUIET
___________ JURY PRESIDENT : M. Antoine BERRY ASSESSEURS : M. Philippe JACQUIET M. Emmanuel LIENARD MEMBRE INVITE : Mme Christelle GRISEZ .
Professeur à l’Université Paul-Sabatier de TOULOUSE Professeur à l’Ecole Nationale Vétérinaire de TOULOUSE
Maître de Conférences à l’Ecole Nationale Vétérinaire de TOULOUSE Ingénieur de Recherche à l’Ecole Nationale Vétérinaire de TOULOUSE
Remerciements
À notre président de jury, Monsieur le Professeur Antoine BERRY
Professeur des Universités, Praticien Hospitalier Faculté de Médecine Toulouse-Rangueil
Parasitologie-Mycologie du CHU de Toulouse
Qui m’a fait l’honneur d’accepter la présidence de mon jury de thèse, Hommages respectueux.
À Monsieur le Professeur Philippe JACQUIET
Professeur à l’Ecole Nationale Vétérinaire de Toulouse
Parasitologie et Maladies Parasitaires
Pour avoir accepter d’être mon Directeur de thèse, Pour m’avoir encadrée et accompagnée dans ce travail Pour votre patience et votre grande disponibilité, Mes remerciements les plus sincères.
À Monsieur le Docteur Emmanuel LIENARD
Maître de conférences à l’Ecole Nationale Vétérinaire de Toulouse
Parasitologie et Maladies Parasitaires
Pour m’avoir fait l’honneur de participer à ce jury de thèse Merci pour votre implication et vos nombreux conseils, Sincère reconnaissance.
À Madame Christelle GRISEZ
Ingénieur de recherche INRA-ENVT
Laboratoire de Parasitologie de l’École Nationale Vétérinaire de Toulouse
Pour m’avoir enseigné les bases du travail de laboratoire, Merci pour votre patience, votre flexibilité Et votre investissement dans mon travail de thèse.
À Madame le Docteur Armelle DIQUELOU
Maître de Conférences à l’École Nationale Vétérinaire de Toulouse
Médecine Interne
Pour son aimable collaboration et son aide précieuse, Tous mes remerciements.
À Madame le Docteur Christelle CAMUS
Maître de Conférences à l’École Nationale Vétérinaire de Toulouse
Biologie et Génétique Moléculaires
Pour ses conseils avisés, Tous mes remerciements
À Monsieur le Docteur Fabrice CONCHOU
Maître de Conférences à l’École Nationale Vétérinaire de Toulouse
Service d’Imagerie médicale
Sincères remerciements.
À Madame le Docteur Séverine BOULLIER
Maître de Conférences à l’École Nationale Vétérinaire de Toulouse
Immunologie
Sincères remerciements.
Au Professeur Domenico OTRANTO de l’Université de Bari en Italie,
Pour nous avoir fait parvenir l’ADN nécessaire pour initier nos manipulations, Sincère reconnaissance.
Aux Docteurs Angela IONICA et Loredana POP de la Faculté de médecine vétérinaire de Cluj-Napoca en Roumanie
Pour nous avoir envoyé les filaires, Sincères remerciements.
À Madame Françoise PREVOT
Assistante ingénieur au laboratoire de Parasitologie de l’ENVT
Pour m’avoir encadré pour une partie des manipulations au laboratoire, Un grand merci.
À l’ESCAPP (European Scientific Counsel Companion Animal Parasites)
Pour l’intérêt porté à mon projet de thèse et leur indispensable contribution, Sincère reconnaissance.
À tous les vétérinaires qui ont participé de près ou de loin à mon travail de thèse,
Au Docteur RAMSAMY sur l’île de la Réunion, ainsi qu’aux Docteurs THOMAS et MELOT,
Au Docteur FRANTZ en Nouvelle-Calédonie et aux Docteurs LESEIGNEUR et LAMAGNERE,
Au Docteur IBENE en Guadeloupe,
Au Docteur GREMION, vétérinaire du refuge de Flayosc, et à Julie, l’ASV avec qui j’ai pu réaliser les prélèvements sur place,
À la Clinique Vétérinaire du Vernet et aux Docteurs CHARRU, MAMOUHDI et ZAHRA pour avoir pensé à moi quand l’occasion s’est présentée.
Pour avoir accepté de participer à ce travail de thèse, Pour avoir eu la patience et le courage de réaliser les prélèvements, Sans vous, ce travail n’aurait pu être mené à bien. Sincères remerciements à tous.
À l’ensemble de mes proches,
Pour m’avoir toujours soutenu dans mes choix et m’avoir permis de réaliser mes rêves, Merci infiniment.
Table des matières
Table des matières ... 1
Table des annexes ... 6
Table des figures ... 7
Table des tableaux ... 11
Introduction ... 13
Partie 1 : Étude bibliographique : La dirofilariose canine à Dirofilaria immitis et les autres filarioses du chien ... 15
I. Les espèces de filaires parasites du chien ... 16
1) Classification systématique des filaires du chien ... 16
2) Répartition géographique ... 18
a) Répartition géographique des filarioses canines ... 18
b) Répartition géographique des cas zoonotiques liés aux filaires du chien ... 20
3) Morphologie des filaires adultes et des microfilaires ... 22
a) Morphologie des vers adultes ... 22
b) Morphologie des microfilaires (larves L1) ... 24
4) Biologie et cycle évolutif ... 27
a) Dirofilaria immitis ... 27
i. Hôtes définitifs ... 27
ii. Hôtes intermédiaires ... 29
iii. Transmission des larves L3 infestantes à l’hôte définitif ... 32
iv. Schéma récapitulatif du cycle de Dirofilaria immitis chez le chien ... 33
b) Dirofilaria repens ... 34
c) Acanthocheilonema reconditum ... 35
d) Acanthocheilonema dracunculoïdes ... 37
e) Cercopithifilaria spp. ... 37
f) Onchocerca lupi ... 38
5) La bactérie Wolbachia pipientis, symbiote des filaires ... 40
a) Présentation générale ... 40
b) Symbiose entre les bactéries et les filaires ... 40
c) Wolbachia et Dirofilaria immitis ... 40
6) Dirofilaria immitis, agent de la dirofilariose cardio-pulmonaire ... 42
i. Pathogénie de la forme chronique classique de la dirofilariose cardio-pulmonaire .. 45
ii. Pathogénie du Syndrome de la Veine Cave ... 49
iii. Pathogénie de la forme occulte liée à la destruction immunologique des microfilaires 49 c) Étude clinique chez le chien ... 51
i. Forme classique = Forme chronique cardio-pulmonaire ... 51
ii. Formes aigues ... 53
iii. Forme occulte liée à la destruction immunologique des microfilaires ... 55
iv. Formes atypiques ... 55
d) Eléments d’orientation de biologie clinique ... 57
i. Lors de forme classique de la dirofilariose ... 57
ii. Lors de Syndrome de la Veine Cave ... 58
e) Anomalies des images radiographiques, échocardiographiques et de l’électrocardiogramme lors de dirofilariose cardio-pulmonaire ... 59
i. Les images radiographiques thoraciques lors de dirofilariose ... 59
ii. Les images échocardiographiques lors de dirofilariose ... 61
iii. L’électrocardiographie (ECG) ... 62
f) Potentiel zoonotique de Dirofilaria immitis ... 63
7) Pouvoir pathogène de Dirofilaria repens et des autres filaires parasites du chien ... 64
a) Dirofilaria repens ... 64
b) Acanthocheilonema reconditum ... 66
c) Acanthocheilonema dracunculoïdes ... 66
d) Genre Cercopithifilaria ... 67
e) Onchocerca lupi ... 68
II. Diagnostic expérimental des filarioses canines ... 71
1) Mise en évidence et identification des microfilaires ... 71
a) Mise en évidence des microfilaires sanguines ... 71
b) Identification de l’espèce de microfilaires ... 73
c) Limites des méthodes de détection des microfilaires sanguines ... 76
d) Mise en évidence des microfilaires dermotropes ... 79
2) Mise en évidence d’antigènes circulants de Dirofilaria immitis ... 80
a) Principe des tests sérologiques ... 80
b) Les différents tests sérologiques disponibles... 82
c) Intérêts et limites des tests de détection des antigènes circulants ... 83
3) Méthodes de diagnostic moléculaire ... 84
III. Traitement et prévention de la dirofilariose canine ... 86
1) Traitement de la dirofilariose canine à Dirofilaria immitis ... 86
a) Traitement médical ... 86
i. Evaluation préalable au traitement ... 86
ii. Traitement adulticide ... 87
iii. Traitement microfilaricide ... 90
iv. Traitements adjuvants ... 92
v. Traitement de la thromboembolie pulmonaire ... 93
b) Traitement chirurgical ... 94
i. Syndrome de la Veine Cave ... 94
ii. Localisation oculaire ... 95
c) Recommandations actuelles en termes de traitement ... 96
2) Prophylaxie de la dirofilariose canine ... 98
a) Mesures sanitaires et chimiques pour éviter l’infestation... 98
b) Mesures médicales de chimioprévention ... 99
c) La résistance aux lactones macrocycliques ... 101
3) Traitement et prévention des autres filarioses canines ... 103
a) Dirofilaria repens ... 103
i. Traitement de la dirofilariose sous-cutanée à Dirofilaria repens ... 103
ii. Prévention de la dirofilariose sous-cutanée à Dirofilaria repens ... 104
b) Acanthocheilonema reconditum et Acanthocheilonema dracunculoides ... 105
c) Cercopithifilaria spp. ... 105
d) Onchocerca lupi ... 105
Partie 2 : Partie expérimentale : Mise en place d’une méthode de diagnostic moléculaire par PCR quantitative pour détecter et identifier les microfilaires sanguines de Dirofilaria immitis et de Dirofilaria repens chez le chien ... 107
I. Objectifs de l’étude expérimentale ... 108
II. Matériels et méthodes ... 109
1) Recrutement des chiens et modalités de prélèvements ... 109
a) Principes de recrutement des chiens ... 109
b) Modalités de recrutement des chiens ... 110
c) Profil des chiens inclus dans l’étude... 111
d) Modalités de prélèvement ... 112
e) Conservation et envoi des échantillons ... 112
2) Méthodes diagnostiques ... 113
b) Test rapide de détection des antigènes solubles de Dirofilaria immitis ... 114
c) PCR quantitative duplex ... 115
i. Extraction de l’ADN... 115
ii. Principe de la PCR en temps réel avec la méthode EvaGreen® ... 116
iii. Témoins positifs et Témoin négatif ... 119
iv. ADN Matrice et amorces de la qPCR ... 120
v. Protocole pour la PCR en temps réel ... 121
III. Résultats de l’étude expérimentale ... 125
1) Résultats du Filarassay® et du Snap 4DX® IDEXX ... 125
2) Résultats de la PCR en temps réel ... 128
a) Expériences préliminaires ... 128
i. Mise en place à l’ENVT du protocole de PCR quantitative décrit par Latrofa et al. en 2012 avec les extraits d’ADN de Dirofilaria immitis et de Dirofilaria repens provenant d’Italie ... 128
ii. Nouveaux témoins positifs obtenus à partir de filaires adultes de Roumanie ... 129
iii. Résultats des qPCR simplex sur les témoins positifs et séquençage des séquences amplifiées ... 130
b) Résultats de la qPCR pour la recherche d’ADN des microfilaires sanguines sur les chiens prélevés dans l’étude ... 131
i. Pour les chiens de France métropolitaine ... 131
ii. Pour les chiens des DOM-TOM ... 134
3) Synthèse des résultats ... 136
a) Pour les chiens de France métropolitaine ... 136
b) Pour les chiens des DOM-TOM ... 137
c) Tableau Bilan ... 138
d) Étude statistique ... 139
IV. Discussion ... 141
1) Population canine étudiée et prélèvements ... 141
a) Choix des chiens prélevés ... 141
b) Modalités du prélèvement ... 141
c) Conservation du sang total sur EDTA ... 142
d) Transport des prélèvements ... 142
2) Méthode de concentration pour détecter les microfilaires ... 143
a) Choix de la méthode ... 143
b) Limites de la méthode ... 143
3) Test de détection des antigènes de Dirofilaria immitis ... 144
b) Limites de l’interprétation ... 144
4) PCR en temps réel ... 145
a) Choix des amorces ... 145
b) Choix des témoins positifs ... 145
c) Sensibilité de la PCR en temps réel ... 145
i. Nécessité d’une dilution au 1/10ème de l’extrait d’ADN ... 145
ii. Sensibilité de la qPCR par rapport au test Filarassay® ... 147
iii. Sensibilité de la qPCR par rapport au Snap 4DX® ... 147
d) Spécificité de la PCR en temps réel... 148
i. Amplifications non spécifiques ... 148
ii. Spécificité de la qPCR comparée aux autres tests ... 148
5) Interprétation des résultats ... 149
Conclusion ... 152
Bibliographie ... 154
Table des annexes
Annexe 1 : Fiche de consentement éclairé et Fiche d’accompagnement des prélèvements …………168 Annexe 2 : Taux de réponse et de participation des vétérinaires contactés……….170 Annexe 3 : Liste des chiens inclus dans l’étude et résultats des différents tests………..171 Annexe 4 : Protocole d’extraction de l’ADN avec le kit QIAGEN « QIAamp®DNA Mini Kit »…..176 Annexe 5 : Résultats des PCR en temps réel réalisées………..………177 Annexe 6 : Purification du produit d’amplification de PCR avec le Kit d’extraction d’ADN
Table des figures
Figure 1 : Classification taxonomique des principales espèces de filaires parasites du chien ... 17 Figure 2 : Distribution géographique actuelle des parasites Dirofilaria immitis et Dirofilaria repens d’après Simón et al, 2012 ... 18 Figure 3 : Zones approximatives d’enzootie de Dirofilaria immitis (à gauche) et Dirofilaria repens (à droite) en Europe d’après Otranto et al. 2013 ... 19 Figure 4 : Distribution d’Acanthocheilonema reconditum, Cercopithifilaria spp. et Onchocerca lupi en Europe d’après Otranto et al. 2013 ... 20 Figure 5 : Distribution actuelle des cas de dirofilariose humaine liée à Dirofilaria immitis et à Dirofilaria repens d’après Simón et al. 2012 ... 21 Figure 6 : Schéma de filaires adultes mâle et femelle de Dirofilaria immitis (Raillet Alcide, 1895) ... 22 Figure 7 : Vers adultes d’Onchocerca lupi au sein d’une masse rétro-bulbaire d’un chien ... 23 Figure 8 : Coupe histologique d’un nodule sous conjonctival montrant les caractéristiques de la cuticule de la femelle d’Onchocerca lupi avec les arêtes (flèches) et les stries (triangles) ... 23 Figure 9 : Extrémités antérieures et postérieures des microfilaires de Dirofilaria immitis (A-B) et Dirofilaria repens (C-D) D’après Traversa et al. 2010 ... 24 Figure 10 : Observations de microfilaires d’Acanthocheilonema reconditum au microscope photonique (A) Microfilaire entière ; (B) Extrémité caudale en crochet ; (C) Corps interne D’après Napoli et al. 2014 & Rodríguez García 1990 ... 24 Figure 11 : Taille moyenne des microfilaires évaluée après fixation avec la technique de Knott modifié sur 377 chiens D’après Magnis et al. 2013 ... 25 Figure 12 : Schéma représentant une microfilaire de l’espèce Cercopithifilaria sp.II en vue latérale et en vue dorso-ventrale, d’après Otranto et al. 2011 ... 25 Figure 13: Observation microscopique de microfilaires de : (A) Cercopithifilaria sp. II; (B) Cercopithifilaria bainae ; (C) Cercopithifilaria grassii D’après Cortes et al. 2014 ... 26 Figure 14 : Microfilaire cutanée d’Onchocerca lupi observée au microscope électronique. D’après Otranto et al. 2013 ... 26 Figure 15 : Photographie de filaires adultes de Dirofilaria immitis dans le cœur droit d’un chien ... 28 Figure 16 : Modélisation de l’extrémité antérieure d’une filaire adulte de Dirofilaria immitis Source : Curtis. 2015 ... 28 Figure 17 : Culex pipiens pipiens (à gauche) et Aedes albopictus (à droite) ... 29 Figure 18 : Schéma d’un moustique absorbant des microfilaires au cours du repas sanguin et photographie de la trompe d’un moustique contenant une larve L3 (Sources : P. Bourdeau, ONIRIS) 31 Figure 19 : Nombre de générations potentielles de Dirofilaria immitis chez les moustiques vecteurs en une année d’après Genchi et al. 2009 ... 31 Figure 20 : Représentation schématique du cycle de Dirofilaria immitis élaborée d’après un schéma issu de l’article de Genchi et al. 2013... 33 Figure 21 : Représentation schématique du cycle de Dirofilaria repens élaborée d’après un schéma issu de l’article de Genchi et al. 2013... 35 Figure 22 : Coupes histologiques d’une puce infestée par A. reconditum colorées à l'hématoxyline et à l'éosine ... 36
Figure 23 : (A) Coupe histologique du derme où des microfilaires (indiquées par les flèches) ont été détectées par biopsie cutanée; (B) (C) (D) Larves de Cercopithifilaria sp.II provenant de tiques disséquées (B) Stade L1, (C) Stade L2, (D) Stade L3 D’après Otranto et al. 2012 ... 38 Figure 24 : Photographies de Simulium reptans (à gauche) et d’un Culicoïde (à droite) Crédit Illustration : Google Images ... 39 Figure 25 : Mise en évidence de protéines de surface de Wolbachia (WSP) par immuno-histochimie 41 Figure 26 : Cartes indiquant le nombre moyen de cas de dirofilariose à Dirofilaria immitis recensés par clinique vétérinaire aux Etats-Unis en 2001 et en 2013, d’après le site web de l’American Heartworm Society (AHS) ... 42 Figure 27 : Répartition géographique des cas de dirofilariose cardio-pulmonaire des carnivores en France d’après Bourdeau et al. 2008 ... 44 Figure 28 : Représentation schématique de la physiopathologie de la dirofilariose cardio-pulmonaire 46 Figure 29 : Altérations pathologiques des artères pulmonaires associées à la dirofilariose cardio-pulmonaire ... 47 Figure 30 : Comparaison d’une coupe histologique d’un glomérule normal et d’un glomérule avec glomérulonéphrite à immuns complexes (d’après Cours ENVT – Immunologie A2) ... 48 Figure 31 : Forme oculaire de la dirofilariose à Dirofilaria immitis, d’après Hayasaki et al. 2013 ... 56 Figure 32 : Radiographies thoraciques d’un chien atteint de dirofilariose cardio-pulmonaire ... 60 Figure 33 : Images échocardiographiques d’un chien atteint de dirofilariose cardio-pulmonaire. Noter la présence de deux filaires adultes qui apparaissent comme deux lignes hyperéchogènes parallèles. Crédit photographique : cal.vet.upenn.edu/project ... 62 Figure 34 : Radiographie thoracique d’un homme montrant un nodule pulmonaire attribué à Dirofilaria immitis D’après Simón et al. 2012 ... 63 Figure 35 : Photographies de chien atteints de dirofilariose sous-cutanée. ... 64 Figure 36 : Exemples d’atteintes oculaires liées à Onchocerca lupi ... 68 Figure 37 : Analyses histologiques de lésions d’onchocercose canine (A) Coupe histologique d’un nodule montrant plusieurs sections de filaires mâles et femelles gravides enroulées (B) Coupe histologique des tissus environnants mettant en évidence des microfilaires ... 69 Figure 38 : Observation d’une microfilaire de Dirofilaria repens sur un frottis sanguin coloré au May-Grünwald Giemsa à l’objectif x40 ... 71 Figure 39 : Observation de microfilaires de Dirofilaria immitis (à gauche) et de Dirofilaria repens (à droite) sur filtre Filarassay® au grossissement x400 ... 73 Figure 40 : Observation microscopique de microfilaires d’Acanthocheilonema reconditum (en haut) et de Dirofilaria immitis (en dessous). Crédit photographique: Byron Blagburn, PhD ... 74 Figure 41 : Schéma anatomique d’une microfilaire D’après Ducos de Lahitte 1990... 74 Figure 42 : Représentation schématique des zones d’activité phosphatasique acide des microfilaires D’après Schrey and Trautvetter. 1998 ... 75 Figure 43 : Observation microscopique des microfilaires et de leurs zones d’activité phosphatasique acide au grossissement x400. D’après Schrey and Trautvetter. 1998 ... 76 Figure 44 : Microfilaire cutanée d’Onchocerca lupi observée au microscope photonique. ... 79 Figure 45 : Principe du Snap Test® (d’après IDEXX) ... 81 Figure 46 : Représentation schématique du principe d’un test d’immunochromatographie modifié d’après un schéma de sciencesetavenir.fr ... 82 Figure 47 : Lecture des résultats du Snap Test Dirofilariose® des laboratoires IDEXX ... 82 Figure 48 : IMMITICIDE® des laboratoires MERIAL, le seul traitement adulticide avec une AMM 87
Figure 49 : Evolution de la microfilarémie chez des chiens recevant de l’ivermectine seule (IVM) 1 fois par semaine, de la doxycycline seule (DOXY) par intermittence, les deux (IVM+DOXY), avec ou sans mélarsonine (MEL) à la semaine 26 (protocole en 3 injections). Ces chiens s’étaient préalablement vu transplantés des filaires adultes 6 semaines avant le début du protocole. D’après McCall et al. 2008. ... 92 Figure 50 : Méthode d’extraction des filaires avec l’introduction d’un cathéter introducteur. (A) Introduction du cathéter introducteur dans la veine de jugulaire externe gauche guidée par le fil de guide pré placé dans l'atrium droit, le ventricule droit et l'artère pulmonaire. (B) et (C) Introduction de la pince préhensile pour retirer les vers sous contrôle fluoroscopique. D’après Won-Kyoung Yoon et al. 2011 ... 95 Figure 51 : Schéma récapitulatif du protocole de traitement actuellement recommandé par l'American Heartworm Society. ... 96 Figure 52 : Trois exemples de spécialités vétérinaires disponibles pour prévenir les piqûres de moustiques, vecteurs de la dirofilariose cardio-pulmonaire ... 98 Figure 53 : Sensibilité des différents stades évolutifs de Dirofilaria immitis aux lactones macrocycliques et à la mélarsomine, d’après Wolstenholme et al. 2015 ... 99 Figure 54 : Origine et profil épidémiologique des chiens dont le sang a été prélevé pour l’étude ... 111 Figure 55 : Illustration d’un test Filarassay® réalisé au laboratoire de parasitologie de l’ENVT Crédit photographique : J. Semat ... 113 Figure 56 : Observation de microfilaires de Dirofilaria immitis au microscope photonique au grossissement x 400 ... 113 Figure 57 : Réalisation des Snap 4DX® au laboratoire de parasitologie de l’ENVT ... 114 Figure 58 : Protocole simplifié de la méthode d'extraction de l'ADN avec le kit QIAGEN « QIAamp® DNA Mini Kit », qui utilise la technologie basée sur la silice. ... 115 Figure 59 : Principe de la PCR quantitative utilisant un agent fluorescent se liant à l’ADN double brin (EvaGreen®). D’après Poitras et Houde. 2002 ... 117 Figure 60 : Courbe d’amplification de la PCR en temps réel. L’intensité de la fluorescence à chaque cycle est proportionnelle à la concentration d’amplicons, le cycle seuil (Ct) représente le nombre de cycles requis où le signal d’émission de fluorescence est statistiquement et significativement plus élevé que la ligne de base ... 117 Figure 61 : Représentation schématique de l’évolution de la fluorescence en fonction de la température (à gauche) et de la courbe de dissociation d’une réaction de qPCR (à droite). La température de fusion d’un double brin correspond à un pic sur cette courbe. ... 118 Figure 62 : Courbe de dissociation obtenue à l’issue d’une réaction de qPCR qui illustre la présence d’une séquence amplifiée non spécifique. ... 118 Figure 63 : Microplaques PCR 96 puits Thermo Scientific AB-0600... 122 Figure 64 : Représentation schématique d’un plan de plaque de la qPCR avec 19 échantillons à tester ... 122 Figure 65 : Observation microscopique d’une microfilaire de Dirofilaria repens (identification moléculaire à postériori) sur le prélèvement de sang du chien V1 issu de la Clinique Vétérinaire du Vernet (31) par la méthode par filtration au grossissement x400 ... 125 Figure 66 : Observation microscopique de microfilaires de Dirofilaria immitis (identification moléculaire à postériori) sur le prélèvement de sang du chien R10 provenant de la Réunion par la méthode par filtration Filarassay® au grossissement x 100 (à gauche) et x400 (à droite) ... 125 Figure 67 : Résultats des Snap 4DX®IDEXX sur le sang des chiens provenant de Guadeloupe ... 126
Figure 68 : Courbes de dissociation obtenues à partir des échantillons d’ADN extraits de filaires adultes des deux espèces : deux pics matérialisent des températures de fusion Tm différentes : un premier pic à 71,2°C pour l’extrait d’ADN de Dirofilaria repens et un deuxième à 76,7°C pour l’extrait d’ADN de Dirofilaria immitis. ... 128 Figure 69 : Courbes de dissociation obtenues à partir des échantillons d’ADN extraits de filaires adultes de Dirofilaria immitis et de Dirofilaria repens provenant de Cluj-Napoca en Roumanie. ... 129 Figure 70 : Courbes de dissociation obtenues à partir de l’échantillon d’ADN extrait du sang du chien ENVT 1 après dilution au 1/10ème, le pic à 70,7°C correspondant à l’espèce Dirofilaria repens ... 131 Figure 71 : Courbes de dissociation obtenues à partir de l’échantillon d’ADN extrait du sang du chien ENVT 2 après dilution au 1/10ème, le pic à 70,7°C correspondant à l’espèce Dirofilaria repens. ... 132 Figure 72 : Courbes de dissociation obtenues à partir de l’échantillon d’ADN extrait du sang du chien V1 de la clinique vétérinaire du Vernet (31) après dilution au 1/10ème ... 132 Figure 73: Courbes de dissociation obtenues à partir de l’échantillon d’ADN extrait du sang du chien V2 de la clinique vétérinaire du Vernet (31) après dilution au 1/10ème ... 133 Figure 74 : Courbes de dissociation obtenues à partir de l’échantillon d’ADN extrait du sang du chien F1 du refuge de Flayosc (83) après dilution au 1/10ème ... 133 Figure 75 : Courbes de dissociation obtenues à partir des échantillons d’ADN extrait du sang des chiens R10 (à gauche) et RL (à droite) de la Réunion... 134 Figure 76 : Courbes de dissociation obtenues à partir des échantillons d’ADN extrait du sang des chiens G3 (à gauche) et G4 (à droite) de Guadeloupe ... 134 Figure 77 : Courbes de dissociation obtenues à partir des échantillons d’ADN extrait du sang des chiens C6 (à gauche) et NC11 dilué au 1/10ème (à droite) de Nouvelle-Calédonie ... 135
Table des tableaux
Tableau 1 : Espèces de moustiques vecteurs et localisation de la région dans laquelle la transmission de Dirofilaria immitis a été démontrée, tableau traduit en français d’après Simón et al. 2012 ... 30 Tableau 2 : Modifications biochimiques fréquemment rencontrées lors de dirofilariose à Dirofilaria immitis ... 57 Tableau 3 : Modifications hématologiques fréquemment rencontrées lors de dirofilariose à Dirofilaria immitis ... 57 Tableau 4 : Modifications biochimiques et hématologiques fréquemment rencontrées lors de Syndrome de la Veine Cave ... 58 Tableau 5 : Description des anomalies radiologiques selon le stade d’évolution de la dirofilariose cardio-pulmonaire ... 59 Tableau 6 : Espèces d’helminthes impliquées dans des affections oculaires chez le chien d’après Sréter et Széll. 2008 ... 69 Tableau 7 : Caractères morphologiques des microfilaires sanguines rencontrées chez le chien D’après Magnis et al. 2013 et Euzeby. 1990 ... 74 Tableau 8 : Localisation des principaux éléments anatomiques des microfilaires sanguines du chien en % par rapport à la longueur totale ... 74 Tableau 9 : Caractères morphologiques des microfilaires sanguines, dermiques et des larves cutanées rencontrées chez le chien. D’après Sréter et Széll. 2008 ... 78 Tableau 10 : Recommandations actuelles en termes de protocole de traitement de la dirofilariose canine à Dirofilaria immitis, d’après les recommandations de l’American Heartworm Society ... 97 Tableau 11 : Les spécialités vétérinaires disponibles disposant d’une AMM pour la prévention de la dirofilariose canine ... 100 Tableau 12 : Amorces utilisées pour la détection de l’ADN de Dirofilaria immitis, d’après Latrofa et al. 2012 ... 120 Tableau 13 : Amorces utilisées pour la détection de l’ADN de Dirofilaria repens, d’après Latrofa et al. 2012 ... 120 Tableau 14 : Tableau récapitulatif des composants intervenants dans la PCR quantitative duplex .... 122 Tableau 15 : Tableau récapitulatif des résultats obtenus pour la recherche de microfilaires sanguines de Dirofilaria immitis et Dirofilaria repens avec la méthode par filtration Filarassay® et pour le Snap 4DX® d’IDEXX pour la recherche des antigènes de Dirofilaria immitis ... 126 Tableau 16 : Récapitulatif des températures de fusion et des Ct obtenus pour les extraits d’ADN des 2 espèces de filaires provenant de l’Université de Bari en Italie (résultats en triplicata) ... 128 Tableau 17 : Tableau récapitulatif des températures de fusion et des Ct obtenus pour les extraits d’ADN des 2 espèces de filaires provenant de l’Université de Cluj-Napoca en Roumanie (résultats en triplicata) ... 129 Tableau 18 : Tableau récapitulatif des résultats de Ct obtenus en qPCR simplex pour chaque couple d’amorces avec l’ADN de chacunes des 2 espèces de filaires provenant de l’Université de Cluj-Napoca en Roumanie ... 130 Tableau 19 : Nombres de cycle de qPCR (Ct) permettant aux sondes fluorescentes de dépasser le seuil de positivité pour les chiens de Nouvelle-Calédonie dont la qPCR est positive ... 135
Tableau 20 : Récapitulatif des résultats obtenus aux différents tests diagnostiques sur les chiens provenant de l’ENVT et de la Clinique Vétérinaire du Vernet (31)... 136 Tableau 21 : Récapitulatif des résultats obtenus aux différents tests diagnostiques sur les chiens provenant de la Réunion ... 137 Tableau 22 : Récapitulatif des résultats obtenus aux différents tests diagnostiques sur les chiens provenant de Guadeloupe ... 137 Tableau 23 : Récapitulatif des résultats obtenus aux différents tests diagnostiques sur les chiens provenant de Nouvelle-Calédonie ... 138 Tableau 24 : Tableau récapitulatif qui met en parallèle les résultats obtenus au test Filarassay® et au Snap 4DX® d’IDEXX avec ceux de la PCR quantitative. *Snap 4DX® positif pour Dirofilaria immitis ... 138 Tableau 25 : Tableau croisé des résultats obtenus pour le test Filarassay® et la qPCR pour détecter une infestation par Dirofilaria immitis à partir de prélèvements sanguins de chiens. NB : Le nombre total d’échantillons est ici de 58 car le Filarassay® n’a pas pu être réalisé sur 7 prélèvements ... 139 Tableau 26 : Sensibilité, Spécificité et coefficient Kappa de la qPCR en comparaison avec le test Filarassay® calculés à l’aide du logiciel DAG_Stat ... 139 Tableau 27 : Tableau croisé des résultats obtenus pour le test Snap 4DX® et la qPCR pour détecter une infestation par Dirofilaria immitis à partir de prélèvements sanguins de chiens ... 140 Tableau 28 : Sensibilité, Spécificité et coefficient Kappa de la qPCR en comparaison avec le test Snap 4DX ® calculés à l’aide du logiciel DAG_Stat ... 140 Tableau 29 : Nombre de chiens inclus dans l’étude qui sont infestés par Dirofilaria immitis ou par Dirofilaria repens en France métropolitaine et dans les DOM-TOM ... 149 Tableau 30 : Résultats obtenus avec le test Filarassay®, le Snap 4DX® et la PCR quantitative sur les chiens inclus dans l’étude et provenant de France métropolitaine et des DOM-TOM ... 149
Introduction
Les filarioses les plus fréquemment rencontrées chez le chien en Europe sont imputables à deux espèces appartenant au genre Dirofilaria : Dirofilaria immitis et Dirofilaria repens. Leur cycle évolutif fait intervenir des hôtes intermédiaires de la famille des Culicidés qui transmettent les parasites au cours d’un repas sanguin. Ces deux espèces de filaires ont un potentiel zoonotique, d’où l’importance de les dépister et de les traiter afin d’éviter qu’ils ne servent de réservoir à d’éventuelles infestations humaines.
Dirofilaria immitis est l’espèce la plus pathogène chez le chien et est responsable de la
dirofilariose cardio-pulmonaire, une maladie potentiellement mortelle. Les vers adultes sont localisés dans les artères pulmonaires et le cœur droit et sont responsables du développement d’une hypertension pulmonaire et d’une insuffisance cardiaque droite.
Dirofilaria repens est à l’origine de la dirofilariose sous-cutanée chez le chien. Cette filaire
est peu pathogène et l’infestation est le plus souvent asymptomatique. Néanmoins, il s’agit de l’espèce la plus fréquemment impliquée dans les infestations zoonotiques en Europe.
Les vers femelles de ces deux espèces produisent des microfilaires qui sont libérées dans la circulation sanguine et qui peuvent être observées directement à l’examen du frottis sanguin (sauf dans les cas de dirofilariose occulte). Des méthodes de concentration de ces microfilaires existent et permettent d’augmenter la sensibilité de la détection. Néanmoins, la différentiation morphologique des microfilaires des deux espèces est difficile et nécessite une coloration histochimique, longue et fastidieuse à réaliser pour l’opérateur.
L’objectif de la partie expérimentale de ce travail de thèse est de développer une PCR quantitative duplex qui détecte simultanément les microfilaires de ces deux espèces et permet de les différencier à partir d’un prélèvement de sang périphérique chez le chien.
Partie 1 : Étude bibliographique
La dirofilariose canine à
Dirofilaria immitis
I.
Les espèces de filaires parasites du chien
1) Classification systématique des filaires du chien
Les principales espèces de filaires parasites du chien appartiennent aux genres Dirofilaria,
Acanthocheilonema, Cercopithifilaria et Onchocerca.
Elles se classent dans l’embranchement des Nématodes qui sont des vers cylindriques, non segmentés, pseudo-coelomates avec un tube digestif complet et des sexes séparés.
Elles appartiennent à l’ordre des Spirurida, caractérisé par la présence de phasmides (organes sensoriels en position caudale), d’une bouche non trilabiée, d’un œsophage dépourvu d’élément glandulaire sacciforme et par l’absence de bourse copulatrice chez le mâle (Ducos de Lahitte. 1990).
La dénomination « filaire » provient de l’appartenance à la super-famille des Filarioïdea : ces vers sont très allongés, filiformes et ne présentent pas de vestibule buccal. Leur vulve est antérieure.
L’absence de vulve juxta-buccale classe les filaires du chien dans la famille des Onchocercidae chez laquelle les vers adultes produisent des larves dénommées microfilaires.
Dans cette famille, on distingue deux sous-familles :
- Les Dirofilarinae avec Dirofilaria immitis et Dirofilaria repens.
Le genre Dirofilaria se caractérise par des spicules très inégaux, par des ailes caudales très développées soutenues par de grosses papilles cloacales chez les individus mâles et par l’absence d’ailes latérales.
- Les Onchocercinae avec :
o Acanthocheilonema reconditum (antérieurement Dipetalonema reconditum)
o Acanthocheilonema dracunculoïdes (anciennement Dipetalonema
dracunculoïdes)
o Le genre Cercopithifilaria (Cercopithifilaria bainae, Cercopithifilaria sp.II
et Cercopithifilaria grassii)
La Figure 1 résume la classification taxonomique des principales espèces de filaires rencontrées chez le chien.
Figure 1 : Classification taxonomique des principales espèces de filaires parasites du chien Crédit Illustration : J. Semat
Le chien est l’hôte définitif des espèces de filaires précédemment citées qui sont des parasites du système cardio-vasculaire pour Dirofilaria immitis, du tissu conjonctif pour Dirofilaria
repens, Acanthocheilonema reconditum et Cercopithifilaria spp., de la cavité péritonéale pour Acanthocheilonema dracunculoïdes ou de la région périorbitaire pour Onchocerca lupi.
Certaines de ces espèces ont un potentiel zoonotique et peuvent être impliquées dans des infestations humaines. Leur prévalence ainsi que leur pouvoir pathogène sont très différents.
Dirofilaria immitis et Dirofilaria repens sont les espèces les plus fréquemment retrouvées en
Europe. Dirofilaria immitis présente un pouvoir pathogène important chez le chien et est à l’origine de la dirofilariose cardio-pulmonaire, une maladie potentiellement mortelle. L’infestation du chien par Dirofilaria repens, agent de la dirofilariose sous-cutanée, est le plus souvent asymptomatique mais il s’agit de l’espèce de filaire la plus souvent impliquée dans les infestations zoonotiques en Europe (Simón et al. 2012).
Les espèces du genre Acanthocheilonema et Cercopithifilaria sont beaucoup moins fréquentes et leur pouvoir pathogène est plus anecdotique. Onchocerca lupi est un parasite récemment reconnu causant des lésions nodulaires associées à une atteinte oculaire chez le chien (Sréter et Széll. 2008).
Il faut tenir compte de l’existence de ces espèces dans la diagnose différentielle des microfilaires puisque les espèces des genres Dirofilaria et Acanthocheilonema produisent toutes des larves L1 qui sont libérées dans la circulation sanguine du chien. Pour Onchocerca
lupi et Cercopithifilaria spp., les microfilaires sont présentes dans le derme mais pourraient
accidentellement être retrouvées sur des étalements sanguins si elles ont été ponctionnées dans la peau au moment du prélèvement.
2) Répartition géographique
a) Répartition géographique des filarioses canines
Les filarioses sont des parasitoses rencontrées sur tous les continents du globe et dont la répartition est étroitement liée à celle de leurs vecteurs arthropodes hématophages.
Dirofilaria immitis est endémique en Amérique, en Afrique, en Asie, en Australie et au sud
de l’Europe. Cette espèce est particulièrement présente dans les zones humides et chaudes avec un climat sub-tropical propice au développement des moustiques vecteurs mais elle s’est établie dans des régions plus tempérées durant les vingt dernières années. La répartition des cas de dirofilariose canine à Dirofilaria immitis sera détaillée dans la partie 6) a).
Dirofilaria repens est présente en Afrique, en Asie et en Europe chez mais elle n’a jamais été
signalée sur le continent américain ou en Océanie : par conséquent, le problème du diagnostic différentiel de ses microfilaires avec celles de Dirofilaria immitis ne se pose pas dans ces régions.
Figure 2 : Distribution géographique actuelle des parasites Dirofilaria immitis et Dirofilaria repens d’après Simón et al, 2012
En Europe, le profil épidémiologique de la dirofilariose canine se caractérise par la présence des deux espèces de filaires Dirofilaria immitis et Dirofilaria repens qui coexistent dans certaines régions. Leur aire de répartition tend à s’étendre vers l’Europe centrale et l’Europe du Nord. Ainsi, Dirofilaria repens a été détectée récemment dans de nouvelles régions telles que l’Ukraine, l’Allemagne et le Moyen-Orient. En Italie, la prévalence de Dirofilaria repens atteint les 30% dans la Plaine du Pô (Tarello. 2010).
En France, Dirofilaria repens est plus largement répartie que Dirofilaria immitis : elle est retrouvée en Corse, dans les Bouches du Rhône, dans le Vaucluse, en Haute-Garonne, en Dordogne, dans le Gard, le Gers, en Gironde, en Corrèze, dans la Vienne, le Loir-et-Cher et la région parisienne. La prévalence nationale a été estimée à 1,3% dans une étude publiée en 1986 qui se basait sur la détection des microfilaires dans le sang de 5503 chiens (Ducos de Lahitte. 1990).
Acanthocheilonema reconditum a une distribution mondiale : elle est signalée en Europe au
niveau du bassin méditerranéen, au Moyen-Orient, en Amérique, en Afrique du Sud, en Asie et en Océanie. Cette filaire est largement présente aux Etats-Unis où ses microfilaires sanguines doivent être différenciées de celles de Dirofilaria immitis.
En Sicile, une étude menée sur 2512 chiens par Giannetto et al. en 2007 a démontré qu’il s’agissait de l’espèce de filaire la plus fréquente avec une prévalence de 4-5%. Sa présence en France est plus anecdotique : elle a été estimée à 0,05 % dans une étude publiée en 1990 (Ducos de Lahitte. 1990).
L’aire de distribution d’Acanthocheilonema dracunculoides est réduite à l’Afrique et dans une moindre mesure, à l’Europe. En Afrique, cette filaire est endémique au Maroc, en Algérie, en
Figure 3 : Zones approximatives d’enzootie de Dirofilaria immitis (à gauche) et Dirofilaria repens (à droite) en Europe d’après Otranto et al. 2013
Tunisie, au Mali, au Niger, au Congo, au Soudan, en Somalie, au Kenya, en Tanzanie et en Afrique du Sud (Schwan et Schröter. 2006).
Les espèces du genre Cercopithifilaria sont décrites en Europe et au Brésil. L’espèce
Cercopithifilaria grassii, initialement décrite comme Filaria grassii en 1907 a été retrouvée
en Italie, en Sicile et en Espagne. Cercopithifilaria bainae, découverte au Brésil a été décrite chez des chiens en Italie, en Espagne et en Grèce. La dernière espèce Cercopithifilaria sp. II a été mise en évidence en 2012 en Italie et en Espagne (Cortes et al. 2014).
Onchocerca lupi est présente en Europe (Grèce, Portugal, Allemagne, Hongrie, Suisse) et aux
Etats-Unis (Otranto et al. 2013).
Figure 4 : Distribution d’Acanthocheilonema reconditum, Cercopithifilaria spp. et Onchocerca lupi en Europe d’après Otranto et al. 2013
b) Répartition géographique des cas zoonotiques liés aux filaires du chien
Les cas de Dirofilariose humaine
Les espèces Dirofilaria immitis et Dirofilaria repens possèdent toutes deux un potentiel zoonotique. La distribution mondiale des cas humains ne coïncide pas exactement avec celle des cas de dirofilariose canine, principalement en raison du manque de données puisqu’il existe des pays pour lesquels des cas humains sont rapportés en l’absence de données sur les infestations canines.
Selon des données de 2012, 1782 cas d’infestations humaines par les espèces du genre
Dirofilaria ont été recensées : 372 sont imputables à Dirofilaria immitis qui entraine des
atteintes pulmonaires chez les patients infestés ; 1410 sont dus à Dirofilaria repens qui est à l’origine de troubles cutanés ou oculaires chez l’Homme (Simón et al. 2012).
Sur le continent américain, au Japon et en Australie, les cas humains sont imputables à
Dirofilaria immitis, les infestations liées à Dirofilaria repens restant sporadiques.
En Europe, où ces deux espèces de filaires coexistent dans les populations canines réservoirs, les cas d’infestation humaine impliquant Dirofilaria repens (586 cas rapportés) prédominent largement sur les cas liés à Dirofilaria immitis (33 cas), même dans des zones de forte prévalence de cette dernière.
La France a recensé 71 cas de dirofilariose humaine à Dirofilaria repens et 2 cas de dirofilariose pulmonaire attribuée à Dirofilaria immitis. Elle est le deuxième pays d'Europe après l'Italie en nombre de cas humains recensés pour cette zoonose émergente (Simón et al. 2012).
Figure 5 : Distribution actuelle des cas de dirofilariose humaine liée à Dirofilaria immitis et à Dirofilaria repens d’après Simón et al. 2012
Les cas humains liés aux filaires des genres Acanthocheilonema et Cercopithifilaria
Le potentiel zoonotique de ces espèces de filaires est quasi nul : un seul cas humain a été décrit en 2001 en Australie impliquant Acanthocheilonema reconditum et dans lequel une filaire adulte a été retrouvée en position sous-conjonctivale (Huynh et al. 2001).
Les cas humains liés à Onchocerca lupi
Le potentiel zoonotique de cette filaire a été soupçonné pour la première fois en 2002 et a été confirmé en 2011 en Turquie où des nématodes sous-conjonctivaux ont été identifiés par analyse moléculaire comme étant des vers adultes appartenant à l’espèce Onchocerca lupi (Otranto et al. 2011). Depuis, des dizaines de cas humains ont été rapportés en Turquie, en Tunisie, en Iran et aux États-Unis (Baneth et al. 2015).
3) Morphologie des filaires adultes et des microfilaires
a) Morphologie des vers adultes
Dirofilaria immitis
Les filaires adultes de Dirofilaria immitis ont l’aspect d’une corde à violon blanchâtre. Les femelles mesurent jusqu’à 30 cm de long pour 1 mm de diamètre. Les mâles sont plus petits et plus fins et mesurent entre 12 et 18 cm de long pour 600 μm de diamètre. L’orifice buccal est entouré de 8 papilles médianes et de 2 papilles latérales. Le mâle est identifiable par son extrémité caudale spiralée, en tire-bouchon qui porte 4 à 6 paires de volumineuses papilles pédonculées et ovoïdes (Ducos de Lahitte. 1990).
Dirofilaria repens
Les filaires adultes de Dirofilaria repens, parasites du tissu sous-cutané, ont une taille inférieure à celle de Dirofilaria immitis. Le mâle mesure 5 à 7 cm de long sur 370 à 450 μm de large. Il possède 2 à 4 papilles pré-anales d’un côté et 5 ou 6 de l’autre. Les deux spicules sont inégaux et mesurent 465 à 590 µm et 185 à 206 µm. La femelle mesure 10 à 17 cm de long sur 460 à 650 μm de diamètre. Sa vulve est distante de 1,5 mm à 1,62 mm de l’extrémité antérieure (Ducos de Lahitte. 1990).
Acanthocheilonema reconditum
Cette filaire est plus petite que celles du genre Dirofilaria.
Le mâle mesure 9 à 17 mm de long sur 92 à 106 μm de large. Leur queue est spiralée et se termine par 3 pointes. Les spicules sont inégaux et mesurent respectivement 220 à 300 µm et 92 à 104 µm (Chauve, 1990). La femelle mesure 20 à 32 mm de long sur 146 à 186 μm de large. La vulve est située à 840 μm de l’extrémité antérieure.
Acanthocheilonema dracunloïdes
Acanthocheilonema dracunloïdes a une taille supérieure à celle d’Acanthocheilonema reconditum. Le mâle mesure 15 à 32 mm de long sur 100 à 200 μm. Son extrémité caudale est
spiralée. Les papilles génitales, au nombre de 5, sont très petites. Les spicules sont inégaux et mesurent respectivement 320 à 390 µm et 120 à 160 µm. La femelle mesure 30 à 60 mm de long et 100 à 300 μm de large (Schwan et Schröter. 2006). L’extrémité postérieure est souvent relevée vers la face dorsale. La vulve est éloignée de 1000 μm de l’extrémité céphalique. Cercopithifilaria spp.
Les filaires adultes de Cercopithifilaria grassi sont longues de 7 à 8 mm pour les mâles et de 17 à 21 mm pour les femelles. L’extrémité antérieure présente un rétrécissement suivi de deux renflements. Pour les adultes de Cercopithifilaria bainae : les mâles mesurent entre 9 mm à 12 mm de long sur 44 µm de large et les femelles 14 mm à 19 mm de long pour 72 à 100 µm de large (Otranto et al. 2011). La morphologie des adultes Cercopithifilaria sp.II n’est pas connue (Ionică et al. 2014).
Onchocerca lupi
Les adultes sont de minces vers blanchâtres : les mâles mesurent 4 à 5 cm de long pour 100 à 200 µm de diamètre, les femelles dépassent les 10 cm pour un diamètre de 200 à 400 µm. L’extrémité antérieure est arrondie. La cuticule est striée transversalement.
Chez le mâle, les papilles caudales sont grandes et charnues, le spicule gauche, long de 160-200 µm est légèrement courbé, tubulaire et tandis que le spicule droit est 2 fois plus court et plus large dans sa partie proximale (Sréter et Széll. 2008).
Chez la femelle, la cuticule est composée de deux couches distinctes avec une couche externe qui porte des arêtes disposées en anneau et une couche plus interne avec des stries.
Figure 7 : Vers adultes d’Onchocerca lupi au sein d’une masse rétro-bulbaire d’un chien Crédit photographique : Sréter et Széll. 2008
Figure 8 : Coupe histologique d’un nodule sous conjonctival montrant les caractéristiques de la cuticule de la femelle d’Onchocerca lupi avec les arêtes (flèches) et les stries (triangles)
b) Morphologie des microfilaires (larves L1)
Dirofilaria immitis
Les larves L1 ou microfilaires de Dirofilaria immitis pondues par les femelles fécondées dans le sang périphérique mesurent 290 à 330 μm de long sur 5 à 7 μm de large (Lichtenfelds et al. 1985). Elles sont nues, c'est-à-dire non entourées d’une gaine. Leur queue est rectiligne, fine et allongée.
Dirofilaria repens
Les microfilaires de Dirofilaria repens sont non-engainées et mesurent 300 à 370 μm de long sur 5-8 μm de large. L’extrémité caudale est filiforme et incurvée en « hameçon ».
Figure 9 : Extrémités antérieures et postérieures des microfilaires de Dirofilaria immitis (A-B) et Dirofilaria repens (C-D) D’après Traversa et al. 2010
Acanthocheilonema reconditum
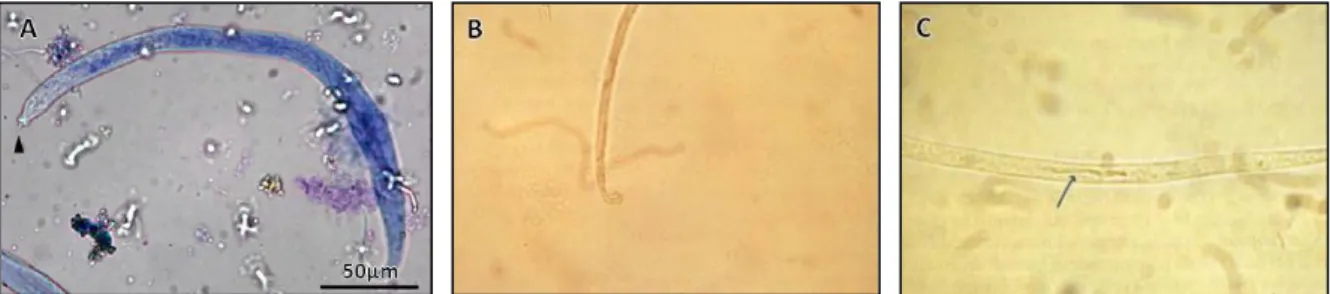
Les microfilaires d’Acanthocheilonema reconditum sont nues et mesurent 263 à 283 μm de long sur 4 μm de large. Leur extrémité caudale est incurvée et forme un « crochet » caractéristique. Le corps interne est visible au niveau du tiers caudal de la larve.
Figure 10 : Observations de microfilaires d’Acanthocheilonema reconditum au microscope photonique (A) Microfilaire entière ; (B) Extrémité caudale en crochet ; (C) Corps interne
D’après Napoli et al. 2014 & Rodríguez García 1990
Acanthocheilonema dracunloïdes
Les microfilaires sont non-engainées et mesurent 185 à 230 mm de long sur 5-6 µm de diamètre. L’extrémité caudale est pointue et longue.
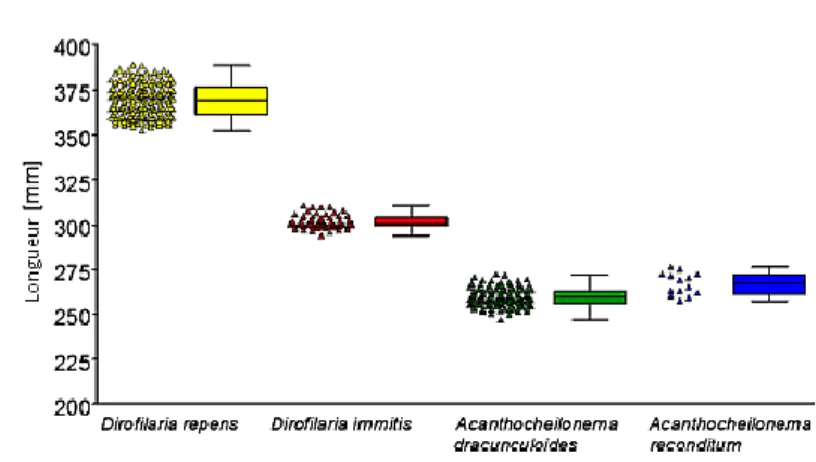
D’après une étude expérimentale publiée en 2013 par Magnis et al. et menée sur 377 chiens infestés provenant de différents pays d’Europe, la longueur moyenne des microfilaires pour ces quatre espèces était de :
- 301,77 ± 6,29 µm pour Dirofilaria immitis (sur 60 chiens) - 369,44 ± 10,76 µm pour Dirofilaria repens (sur 171 chiens)
- 259,43 ± 6,69 µm pour Acanthocheilonema dracunculoides (sur 133 chiens) - 264,83 ± 5,47 µm pour Acanthocheilonema reconditum (sur 13 chiens)
Figure 11 : Taille moyenne des microfilaires évaluée après fixation avec la technique de Knott modifié sur 377 chiens D’après Magnis et al. 2013
Cercopithifilaria spp.
Les microfilaires du genre sont enveloppées d’une mince gaine, sont aplaties dorso-ventralement et présentent un crochet céphalique gauche minuscule. Leur extrémité caudale n’est pas filiforme mais incurvée.
Les microfilaires de l’espèce Cercopithifilaria grassii sont de très grande taille comparée aux autres du genre et mesurent entre 567 et 660 µm de long sur 12,2 à 15,5 µm de large. Leur extrémité caudale est bifide.
Les microfilaires de Cercopithifilaria bainae et de Cercopithifilaria sp.II mesurent respectivement 185,18 µm de long pour 6,59 de large et 182 à 190 µm de long sur 3 à 3,5 µm de large (Otranto et al. 2011).
Figure 13: Observation microscopique de microfilaires de : (A) Cercopithifilaria sp. II; (B) Cercopithifilaria bainae ; (C) Cercopithifilaria grassii
D’après Cortes et al. 2014
Onchocerca lupi
Les microfilaires, qui sont libérées dans le derme sont droites, non engainées, avec une longueur de 98 à 118 µm et une largeur de 5-7 µm. L’extrémité antérieure est arrondie et contient deux à trois noyaux par rangée. La queue est effilée et contient une colonne de noyaux réduite à une seule rangée de trois (Egyed et al. 2001).
Figure 14 : Microfilaire cutanée d’Onchocerca lupi observée au microscope électronique. D’après Otranto et al. 2013
4) Biologie et cycle évolutif
Toutes les espèces de filaires parasites du chien présentent un cycle évolutif de type dixène obligatoire. Il fait intervenir des hôtes définitifs vertébrés et des hôtes intermédiaires arthropodes qui jouent le rôle de vecteurs, sans passage du parasite dans le milieu extérieur.
a) Dirofilaria immitis
Le cycle évolutif de Dirofilaria immitis est un cycle dixène obligatoire qui dure en moyenne entre 6 et 9 mois (Kotani et Powers. 1982).
i. Hôtes définitifs
Le chien et les canidés sauvages tels que le renard, le loup et le coyote sont les hôtes définitifs classiques de Dirofilaria immitis et en constituent les principaux réservoirs. Néanmoins, d’autres espèces peuvent permettre le développement complet du parasite qui présente une faible spécificité d’hôte : c’est le cas des félidés, des mustélidés tels que le furet, des ursidés mais aussi des pinnipèdes (otaries, phoques) chez qui une microfilarémie transitoire a été rapportée (Ducos de Lahitte. 1990).
Les chats sont moins réceptifs que les chiens et ne s’infestent que dans les zones de forte endémie. La prévalence observée chez les félins correspond à 5 à 20% de la prévalence observée dans la population canine pour une région donnée. L’infestation chez le chat est caractérisée par un faible nombre de filaires adultes, 2 à 4 en moyenne qui ont une durée de vie de 2 à 4 ans au maximum. La microfilarémie est rare : seuls 20% des chats infestés par des vers adultes présentent des microfilaires sanguines et ces dernières ne survivent pas plus d’un à 2 mois (Lee et Atkins. 2010) : cette espèce n’est donc pas un réservoir majeur du parasite. Les chats infestés sont souvent asymptomatiques et la guérison spontanée est possible avec la mort des filaires adultes. Il existe pourtant des cas rares d’atteintes respiratoires aigües qui font suite à la réaction inflammatoire liée à l'arrivée des microfilaires dans les artères pulmonaires ou à la mort des adultes.
Il est important de souligner le potentiel zoonotique de Dirofilaria immitis : l’homme peut s’infester mais constitue une impasse épidémiologique et ne peut héberger que des formes immatures qui migrent et meurent enkystés au sein de nodules pulmonaires. Il ne constitue donc pas un réservoir de parasites pour les moustiques (Bourdoiseau. 2000).
Les filaires adultes vivent dans l’artère pulmonaire et ses ramifications ainsi que dans le ventricule cardiaque droit. Ils peuvent migrer dans l’oreillette droite voire la veine cave caudale si l’infestation est massive.
Des localisations erratiques ont également été décrites dans la chambre antérieure de l’œil, le tissu conjonctif sous-cutané, la moelle spinale et la cavité péritonéale.
Les vers adultes ne sont pas hématophages mais se nourrissent d’éléments dissous dans le plasma.
Ilspeuvent survivre jusqu’à 7,5 ans chez le chien (Bowman et Atkins. 2009).
Figure 15 : Photographie de filaires adultes de Dirofilaria immitis dans le cœur droit d’un chien
Crédit photographique : Simón et al. 2012
Figure 16 : Modélisation de l’extrémité antérieure d’une filaire adulte de Dirofilaria immitis
Source : Curtis. 2015
Dirofilaria immitis est vivipare. Les femelles fécondées pondent des larves L1 nues appelées
microfilaires dans le courant circulatoire qui vont principalement se localiser dans les capillaires du tissu conjonctif sous-cutané et intermusculaire de l’hôte définitif. Leur abondance dans la circulation superficielle n’est pas uniforme au cours de la journée et un pic de présence crépusculaire est rapporté : en France, la microfilarémie maximale est observée aux environs de 20h (Euzeby et Lainé. 1951). Il existe également des variations saisonnières de la microfilarémie avec des concentrations plus importantes en été et au printemps, périodes qui correspondent à la période d’activité des moustiques vecteurs (Bowman et Atkins. 2009). Les microfilaires peuvent survivre jusqu’à 2,5 ans dans le sang circulant même après la mort des vers adultes (Bowman et Atkins. 2009). Notons qu’il n’existe pas de relation directe entre la microfilarémie et le nombre d’adultes présents.
Chez 20 à 30% des chiens infestés par Dirofilaria immitis, l’infestation est amicrofilarémique
ou occulte : il n’y a pas de microfilaires circulantes dans le sang périphérique (Schrey et Trautvetter. 1998). Ce type d’infestation s’observe pendant la période pré-patente, lors d’infestation par un seul sexe, un seul ver (infestation mono-parasitaire), lorsque les parasites sont stériles, d’un âge trop avancé ou suite à l’administration de lactones macrocycliques ou de doxycycline. Il existe également des animaux qui développent des réactions d’hypersensibilité vis-à-vis des microfilaires sanguines qui sont détruites et indétectables dans le sang circulant.
La période pré-patente chez le chien, qui sépare le moment où le chien s’infeste de celui où des microfilaires sont présentes pour la première fois dans le sang circulant est de 6 à 9 mois (Kotani et Powers. 1982). Ainsi, si des microfilaires sont retrouvées chez un chiot de moins de 6 mois, il est fort probable qu’elles aient été transmises in utero et que l’animal ne soit pas porteur de filaires adultes.
ii. Hôtes intermédiaires
Les hôtes intermédiaires de Dirofilaria immitis sont des moustiques de la famille des Culicidés appartenant aux genres Culex, Aedes (sous genres : Aedimorphus, Stegomyia),
Anopheles, Mansonia, Armigeres, Ochlerotatus, et Coquillettidia (Otranto et al. 2013).
Près de 70 espèces de moustiques ont été identifiées comme étant des vecteurs potentiels du parasite, ces espèces variant d’une région à l’autre du globe (Morchón et al. 2012).
En Europe, Culex pipiens pipiens, Aedes albopictus (dit moustique tigre) et Aedes vexans sont considérées comme les principales espèces impliquées dans la transmission des filaires du genre Dirofilaria. D’autres espèces ont également été recensées : Culex theileri, Ochlerotatus
caspius, Anopheles maculipennis, Dahliana geniculata, Coquillettidia richiardii.
Aux États-Unis, Aedes albopictus, Aedes sticticus, Aedes trivittatus, Aedes vexans, et Aedes canadensis sont les vecteurs les plus fréquemment mis en cause.
Figure 17 : Culex pipiens pipiens (à gauche) et Aedes albopictus (à droite) Crédit Illustration : Google Images
Tableau 1 : Espèces de moustiques vecteurs et localisation de la région dans laquelle la transmission de Dirofilaria immitis a été démontrée, tableau traduit en français d’après Simón et al. 2012
En région tempérée, la période d’activité des moustiques se limite à la saison chaude (de Juin à Octobre). L’infestation du chien par Dirofilaria immitis a un caractère saisonnier. Certaines espèces de moustiques, comme Culex pipiens, sont actives seulement la nuit tandis que d’autres peuvent l’être au crépuscule voire en pleine journée : Aedes albopictus, Anopheles
maculipennis.
Le moustique femelle est le seul vecteur naturel de transmission de Dirofilaria immitis. Il s’infeste en absorbant des microfilaires au cours de son repas sanguin de type solénophage. Le passage dans l’organisme du moustique est indispensable à la poursuite du cycle évolutif des microfilaires ou larves L1. Ces dernières atteignent les tubes de Malpighi du moustique 36 heures après l’infestation et deviennent des larves L2 au quatrième jour. Au neuvième jour, les larves passent au stade L3 et perforent la paroi des tubes de Malpighi pour atteindre la cavité générale du moustique, puis migrent dans le thorax, la trompe, et enfin dans la cavité du labium.
Ce sont ces larves L3 qui sont infestantes pour l’hôte définitif et qui vont être transmises à l’occasion d’un repas sanguin : les larves émergent de la trompe du moustique et sont déposées sur la peau. Elles sont recouvertes d’hémolymphe, les protégeant de la dessiccation jusqu’à ce qu’elles pénètrent activement dans le tissu sous-cutané du chien par la plaie de piqûre ou par un follicule pileux (McGreevy et al. 1974).
Figure 18 : (A) Schéma d’un moustique absorbant des microfilaires au cours du repas sanguin (Sources : Cours ENVT) (B) Photographie de la trompe d’un moustique contenant une larve L3 (Sources : P. Bourdeau, ONIRIS)
La durée du cycle évolutif chez l’insecte est étroitement liée au climat : pour une température extérieure de 27°C, 10 à 14 jours séparent l’infestation de l’insecte du moment où il est capable d’inoculer des larves L3. Ce délai augmente si les températures sont plus basses et le cycle ne peut se dérouler à des températures inférieures à 14°C (Slocombe et al. 1989). Notons que la durée de vie moyenne pour le moustique vecteur est de 30 jours mais il a été démontré pour plusieurs espèces dont Aedes albopictus qu’elles pouvaient vivre sur des périodes plus longues de 3 mois (Löwenberg Neto and Navarro-Silva. 2004).
La Figure 19 s’appuie sur des relevés de températures réalisées dans toutes les régions d’Europe et propose un modèle qui présente le nombre de générations potentielles de
Dirofilaria immitis chez le vecteur en une année. Ainsi, le sud de l’Europe est plus propice au
développement du parasite du fait des températures plus clémentes : d’une part, la période d’activité des moustiques vecteurs est plus longue et d’autre part, le cycle de développement de la filaire, qui nécessite une température minimale et qui est d’autant plus rapide que la température est élevée, peut se poursuivre sur des périodes plus longues dans l’année.
Figure 19 : Nombre de générations potentielles de Dirofilaria immitis chez les moustiques vecteurs en une année d’après Genchi et al. 2009
Le taux de mortalité chez les moustiques infestés par des stades larvaires de Dirofilaria
immitis est plus important que chez des moustiques indemnes et est positivement corrélé au
nombre de microfilaires ingérées (Serrão et al. 2001). Le nombre de microfilaires ingérées est directement lié à la microfilarémie du chien sur lequel est pris le repas de sang. Les microfilaires puis les stades larvaires L2 et L3 sont pathogènes pour les moustiques s’ils sont présents en trop grand nombre : la présence de plus de 5 à 10 larves L3 dans les tubes de Malpighi peut entrainer leur rupture et la mort de l’insecte. Ainsi, l’ingestion d’un grand nombre de microfilaires par le moustique vecteur au cours d’un repas de sang pris sur un chien présentant une microfilarémie élevée est associée à un taux de mortalité important chez ces derniers. Il a par exemple été démontré que pour un repas de sang pris sur un chien présentant 7000 microfilaires/mL de sang, le taux de survie des moustiques à 11 jours était de 1/10 (Serrão et al. 2001). Les animaux avec une forte microfilarémie ne constituent donc pas une bonne source de parasites puisque l’infestation du moustique est létale et ne permet pas la transmission à un autre hôte vertébré (Bowman et Atkins. 2009).
La capacité des moustiques à permettre le développement de Dirofilaria immitis et à résister à l’infestation est génétiquement déterminée et est contrôlée par des gènes récessifs liés au sexe. Les différences génétiques qui influencent la transmission peuvent aussi exister au sein d’une même espèce : ainsi, certaines souches permettraient l’évolution du parasite tandis que d’autres y seraient réfractaires (Morchón et al. 2007).
iii. Transmission des larves L3 infestantes à l’hôte définitif
Chez le chien, les larves L3 transmises par le moustique profitent de l’effraction causée par la piqûre ou d’un follicule pileux pour pénétrer dans le tissu conjonctif sous-cutané.
Elles effectuent une migration conjonctivo-musculaire pendant laquelle elles muent en L4 au bout de 3 à 12 jours puis en L5 entre le 50ème et le 70ème jour après l’infestation (Simón et al. 2012). Notons que dans le premier mois de migration, les larves (L3 puis L4) sont sensibles aux lactones macrocycliques qui peuvent donc être utilisées en prévention de la maladie.
Les larves L5 pénètrent dans la circulation veineuse, transitent dans le cœur droit et atteignent le tronc pulmonaire puis les artères pulmonaires. Elles s’y développent pendant 1 à 2 mois avant d’effectuer une migration rétrograde vers le ventricule droit où elles deviendront des vers adultes, sexuellement matures environ 120 jours post-infestation.
Les femelles fécondées commenceront à produire des microfilaires (L1) dans le sang périphérique en moyenne 6,5 mois après l’infestation.
Dans les régions tempérées, l’infestation du chien se fait principalement entre Juin et Octobre, période qui correspond à la période d’activité des moustiques vecteurs. Les filaires adultes apparaissent dans le ventricule droit à partir du mois de Décembre et s’ensuit la production de microfilaires vers le mois de Février de l’année suivante. Ces données sont valables pour une primo-infestation, les microfilaires étant présentes toute l’année chez des chiens déjà parasités.
Chez les chiennes en gestation, il est rapporté que les microfilaires peuvent traverser le placenta et ainsi passer dans le sang des chiots. Cette microfilarémie peut être détectable mais ne signifie pas que l’animal va développer une dirofilariose puisque les larves L1 sont incapables de se développer dans l’organisme du chien. La transmission verticale de la maladie n’existe pas.
iv. Schéma récapitulatif du cycle de Dirofilaria immitis chez le chien