~.' [ î :1
1
1(
.
C-C
/ 1..
$li 4),
\,,
•
,.
•
j-• ETUDE SUR LES STADES PHEJOLOGIQUES DU MAIS
AU Q.UEBEC ET SUR' LEUR TRANSMISSIBILITE
par
Raymond Martin
Il
/
Thèse soumise
à
la Faculté des Etude~ Avancées / et' de la Recherche comme par1;ie des exigences'pour l'obtention du diplôme de . Maîtrir ès Science
,
Departmen t: of Plant Science COllège r-lacdonald
Uni vc l:si tÉ> NcGill Nontréal {T ~i ,::i:", .. \ . ~ r
"~;"~~:~~1,n
d,..Ma rt;n
( 1/
1978of
/"
" Août 1977L
,Q,
\
,
tz(
\C
1/
, , ( 1 ,.
1 / 1..
".
• \ "LES STADES PHENOLOGIQUES DU MAlS ET LEUR TRANSMISSIBI4ITE" '
I,'\,,,, " 1 1 !
•
)!j
'"
..
\\
"
, 1 ,-'
"
1
\
1
j.
l't4iL. _'J)
..
l'"..
\
c,"
"---f\
..
f ' SOMMAIREM.Sc~ RAYMOND MARTIN Plant Science
ETUDE SUR LES STADES PHENOLOGLQUES DU MAIS AU QUEBEÇ ET SUR LEUR TRANSMISSIBILITE
Cet,te étude comporte deux volets; dans un premier t~ps on a
analysé la relation entre différen~stades de développement phénologi~ue,
;'
•
les carac~ères morphologiques ,reliés'
à
ces stades et la maturité mesurée, .
~ar ,le pourcentage d'humidité à la récolte (PHR) chez trente-sfx lignées
pures de maïs. oins un deuxième temps no'us avons eétudié la
transmissi-b~lité de ces stades et de ces caractères morphologiques par une étude
en diallèle de dix lignées pures. Pour cette étude le
~odèle
un méthodedeux tel que décrit par Griffing (1957) a été utilisé.
Entre les trentes-six lignées des différences significatives ont \
été observées pour les variables suivantes: nombr~ de jours~du semis
à la levée, nombre de jours de la levée à six feuilles, nombre de jo~rs
de six feuilles à neuf feuilles, nombre de jours de la levée à
l'appJ~it~on
de la panicule, la hauteur~ le nombre de feuilles, le CF de la cinquième
feuille, nombre d'épis par plante et le PHR. qe plus on a obtenu des ,
coefficients de co~rélation hautement significatifs entre'toutes les
variables considérées dans cette étude et le PHR. L'analyse de l~
.régression a démontré q~e la variable unique la plus précise pour prédire
le PHR est le nombre de jours de la levée à l'apparition de la pan1culè •.
Nous avons en outre observé quatre patterns distinpts dans le'développement
du maïs.
Les résultats de l'analyse en dial~èle donfirment las observations
l'
de la premières
e~périence
en ce qui a trait au pattern de développement.f
Par ailleurs, seul l'effet de l'aptitude générale
à
la combinaison pour,le no~re de jours de la levée
à
l'apparition de la panicule est marqué.Pour ce qui est des différentes variables morphologiques mesurées, nous
avons constaté que les
co~rélations po~itives è~
hautementsignificati~es
en~re elles et le PHR ne tiennent plus au niveau des effets de l'AGe de
1 ces var.J.ables. , /
.
!,
i 11
(
c
/
C)
-,
/
.-)
•
ABS.TRACTM.Sc. RAYMOND MARTIN Plant Science
A PHENOLOGlCAL STUDY AND HERITABILITY OF THE DIFFERENT STAGES OF
J DEVELOPMENT IN CORN
~,
The first experiment involvlng an analysis of the relation between
sorne stages of developdent, the morphological characters
a~sociated
withthese stages and, the maturity defined by the percentage of moisture at
harvest (p~m) was carr~ed out. Then, a diallel study of ten parent
\
/
lines was donel to evaluate the heritability of these stages and charac-ters using Griffing (1957) model one, method two.
We noted ~ignificant difference between the thirty-six line studied
in the first experiment for the following variables: number of days
from seeding to emergencei number of days fram emergence ta six leaves, nine leaves, tassellingi heighti number of leaves; LA of the fifth leaf;
number of ears per plant and PMH. Moreover, we found highly significant
correlations between the variables studied and the PMH. Regression
analysis indicatedlthat the singlè most reliable variable to predict the PHH was the number of days from emergence to tasselling. Four distinct development patterns were ê1sa observed.
The results of the diallel analYSi1 confirmed the observations of
1
the first experiment with regard to the pattern of development. The
general combinïng ability effects for aIl characters was significant, \ but only the effect of GCA for the number of days from emergence to tasselling was marked. As for-the morphological characters studied, we
noted that the positive' corrqlations with the PMH observed in the first
experiment were not in complete agreement with the calculate~ GCA for
these characters.
1
1
\"' \c
l
1 ,1 _ f1
'1 \.
\ 1 , Id~ " . "L auteur eSl.re exprllller sa reconnal.ssance a son directeur de thèse, monsieur Ghislain Gendron, Ph.D., et
à ses co-di~ecteurs, messieurs Howard A. Steppler, Ph.D.,
-.
et Norman C. Lawson, Ph.D., pour leur assistance constante et éClair~e. Il tient aussi à lemercier la direction du
Collège Macdonald pour l' ex~ellence de sa ~llaboration::
De plus, cette recherche ,a été rendue possibleogrâce au
..
support financier du Mînistère de l'Education du Québec.
( ' ) <>
/
.
\1
. " ,
1
(
(
()
\
'TABLE DES MATIEREs
LISTE DES TABLEAUX
. . J. . . .
INTRO'bUCTION • ~. • CONTEXTE THEORIQUE 1. Définition de la maturité • • • • 2. Développement Phénolpgique 3. Caractères morpholog~ques • • • •. .
4. Génétique de la maturité . •. J .
EXPERIENCE l 1. Matériel et Méthode- • • • • 1.1 Le facteur,étudié • • • • 1.2 Matériel expérimental ' .• 1. 3 Dispositif expérimental • • • • • • • • • • • 1. 3.1DisP~sitif
statistique • • • • • • • . • • • • 1. 3 • 2 Nombre de blocs • • ._. r. . , '1.3.3 Taille et description de l'unité
expérimenta~1e '~. • • • . .'. • • • •
1.4 Variables mesurées • • • • • • • •
1.5 Conduite et déroul.ement de l'e~érience • 1 . • • • • •
2. Résultats.et Dis~ussion • • • • • • • • • • • • •
2.1 Analyse des résultats • • • • . • • • • • •
2.1.1 Le nombre de plantes levées . • . .
2.1.2 Analyses des variables mesurées • . ' • •
2.1.3 Corrélation entre les variables mesurées
et la facteur étudié .• • • • •. • •
2.1.4 Etude de la régression en placant le PHR
connne, variab1e dépendante • • • • • • • •
2
..
.2 DiscussionEXPERIENCE II ,
1. Matériel et Méthode • • . • • • _. • • • •
1.1 Facteurs étudiés • • . ' . • • • • • • • 1.2 Choix du modèle d'analyse en dia1lêle 1.3 Matériel expérimental • • . • 1.4 Dispositif expérimental
..
.
.
.
,. . .
.
i Page" iii 1 2 6 6 .. 8 11 11 12 13 13 13 15 15 18 19 19 19 20 27 27 33 37 37 38 39 40 (' J-
;.1 .
1
,
f
1
o
Il .
/"
• 1.5 Variables mesurées \ • • . • _ • _ . <1 • •-1.6 Conduite et déroulement de l'e~périence . ~ • • • • 2. Résultats et Discussion • • '. • ~ " .•
2.1 Résultats • . , _ • • • • _ . _ •
'2~1.1 Nombre de plantes levées. • • • • • • •
2.1.2 Analyses de la variance pour les variables ,
mesurees • . • . . • • • l' • • • • •
2 • .1,.3 Etude de l"AGC et l'ASC pour les variables , mesurees 2.2 Discussion CONCLUSION • • • o· • • • • • • • -BIBLIOGRAPHIE i i
/
Page 40 40 40 40 40 41' 43 48 55 56 \r!:
1
(
'-,1\ 1 \\
\\
\ \ 0 ; \ '.\
o
Tableau 1. 2.=If"
3. 4. 5. 6. 7. 8. 9. 10. I l . 12. 13./
LISTE DES TABLEAUX
Liste des lignées de l'expérience 1 •
Analyse de la va:r;iance de la .-riable "nombre de 0
plantes leyées"
. .
. .
.
. .
.
.
.
. . .
.
.
Analyse de la variance de la variable "semis a ... la
levée"
..
. . . . . .
. .
.
. . . . . .
~ésumé des résultats des analyses de la variance pour
les phases l, II, III . . • .
Développement qes 4 lig~ées représentatives des 4
patte,rns obse~és • . . • • • • . • . . 1. • • • • •
1
Moyenne de chaque lignée pour les phases l, II et III
Résumé des résultats de l'analyse de-l~ variance pour
les variables: nombre des feuilles, CFI, nombre d'épis par plante et hauteur • • • • • • • • • • • .
Analyse de la variance pour le facteur PHR
Tableau des moyennes du PHR pour chaque lignée
.
".
(
Coefficient de corrélation de Pearson entre les diverses phases du développement et le PHR
o
Coefficient de corrélation de Pearson entre les'
caractè'res morphologiques et le PHR • • • • .
o
Régression utilisant . 1'" r ... R2 minimum pour la variable
dépendante PHR • • • . . • .
.
\
.
v~leur
du PHR prédit à l'aide desé~uations
des droites de régression pour l et 5 variables au niveau du 0.05et ce pour les lignées 76-26, 76-11, 76-31 et 76-8
Page 14 19 20 21 22 23 24 25 26 28 28 29 32 14." ",,,~lyse de la variance selon la méthode 2 modèle l de
• Gri'ffing (957) • • . • • • • • 38
15-. Liste des lignées sélectionées pour l'analyse en
diallèle • • • • • • • • • • • • • • • • 39
iii
" i l ,
oo
! / Tableau 16. 17. 18. 19. , 20. 2l. 22. 23. ' - -::r 24. 25. 26. 27. 28. 29 .. ~30.Croisements diallèles des 10 lignées sélectionnées Analyse de la variance pour le nombre de plan tes le"Vées • • . • • • . •
Résum~ de l'analyse dei la variance pt>ur les
différentes variables mesurée$ • . • •
,
Nombre de jours du semis à la levée pour les - différents c;-oisernen ts . . . . . . . .
.
.\
.
Analyse de l'AGe et ASC pour le nombre de jours du semis à la levée
Estimation des AGe pour le nOmbre de j ours du semis à
la levée • • . • • • • • . • • • .
1. . . . .
1-Estimation des effets de ASC pour le nombre de jours du
semis à la levée
. .
.
.
..
Résumé de l'analyse de la variance pour l'AGe et l'ASe des diffé1entes phases de développement • • • • • Estimation des effets de l'~Ge des différentes lignées parentales en ce qui a trait aux différentes ~phases du développement . • • • • • . • • • • . • • •
, JUc
pour les croisements 73-11 x Q209, 73-3 x 73-4!73-6 x Q209, 73-2 x Q209 ' " • • • _ • • • • • • _ •
Analyse de la variance des AGe et de l'ASC pour lés
différents caractères morphologiques étudiés
Estimabions de l'effet de l'AGC pour les caractères morphologiques, la hauteur, le nombre de feuilles, le
CF de la cinquième feu~' le, le nombre d' épis par plante
et le PHR • • • • • • • • • • • • .' • ~ • • • • • • •
.?
Estimé des effets de l' ASC pour la hauteur
Estimé des effets de l'ASC pour le CF dé la cinquième feuille
. .
. .
• ! •. .
.
. .
. .
.
.
.
.
.
.
·
.
/
Estimé des effets de l'ASC pour le PHR
. . .
"·
.
·
,•
iv Page 40 41 42 42 43 44...
45 46 47 47 49 49 50 51 l 52, - r
(
•
(
/
l,La culj:ure du J;naïs.au Québec surtout des conditions climatique moment que se manifestent les lorsque le temps est froid et
risques d'échec qui résultent
abl~ du printemps. C'est
à
cecroissance, ~rticulièrement
Grâce
à
la séléction i l~'obtenir
desl~gnée~ prééO~
rentabilité, ma~s qu~ ont par
aille~s l'avantage de mûri,r après année. Par cette étude on tentera
\
d'identifier le patter éveloppement de ces lignées précoces. Nous
procèderons en deux temys, d'abord nous étudierons trente-six lignées précoces pour définir les stades importants dans la croissance du maJs en fonction de la maturité. Puis nous ferons une étude en diallèle de
croisements obtenus de dix lignées pour.voir si on peut retrouver dans les
1
hybrides les mêmes modes de développement " c! est-à-dire s' i~ Y a une
tPtitude à la combinaison pour les caractères choisis au départ.
/
Le but est ,de déterminer 'des stades végétatifs repères pour identifier les lignées et les hybrides précoces avant même qu'ils ne soient
mat~re
plus efficl,ce le trkvail du sélectionneur.1 ", rendus à ~
,
/
1 "- r
"\ .f
(
!
..
0-- "...
cl' , 0=
CONTEXTE THEORIQUE \ ' 1. Définfti~n de la rnaturi té/
,Toute é'fude qui a: pour obj et la ma turi té des plan tes doi t au départ définir de façon précise ce concept. Wolfeo
(1927),' Alherts (1926), Leonard (1935) rapportent. que cette définition manque souvent de rigueur voire même 1 . . . , de préc:j.sion. L'a,mp!gul.té vient du faii. que trop 'souvent l'on confond la maturité et sa mesure. Aldrich (1942), Kiesselbach (1950), et Hallauer et Russel (1962) s' accorden t à dire que le màis est mûr lorsqu'on obtient
"""'!> ..
Q
le poids sec maximum du grain. Pour sa part, Kiesse1bach (1950) ajoute que l'épi
d~it:
avo'tr~~
pourcent d'huntidlté le grain 34." A ce stade, leo ~
" grain ~st trop dur po.ur être denté par une pression de- l'ongle.; Certains auteurs (Shaw et 'r,c;omis 1951; Mayetl 1962; Hanway 1~63) considèrent ce stade comme celui. de la.,.a turi té physio1og~que. On voit qu' i l
est facile de 1 Si embourber au départ si l'on ne peut déteÎminer clairement
la maturité. , 1
..
Nous tenterons donc de résumer
fe~a
façon la plus succinte et.>
exhaustive possible les différentes approches et défini tians de la ma turi té. Evidemment tout dépend de l'objectif visé dans la production du ma1si"
Kae"wei (1954), se référant au-point du vue du producteur
~e s~~ence,
juge le maïs mûr quand la semence .récol t;,ée pourra produire une pousse vigoureuse -même s;t. elle est semée dans des conditions adverses. Hi11son et -penny(1965) utilirent l 'hmnidité du grain comme élément déterminant pour l'entreposage et considèrent J.'humi~ité critique comme le niveau maximum
o où l'on peut stpcker le mài·s dans un crib sans, danger de peéte. Pour Miles
(1940) et Càuderon (1958) ce niveau est de 38 pourcent pour l'épi et ,35
pour le grain.
\
Que o~ soit du point de ~e du product~,~ de semeI\.ces, ou du producteur de ,grain une chose est certaine: ·1 'humidité du, grain
là
la récolte est le2
(
-(
....
-<:J'---/
3facteur déterminant. Donc on revient à notre point de départ, c'est-à-dire que la défini. tian de la _mat uri té se confond avec sa mesure., Et selon·
,
Aldrich (1942) cette mesure devrait posséder les critères suivants: (a) être unique, (b) définir de façon non équivoque si la maturité a été atteinte ou nOJl, (c) servir de base fiable pour comparer les humidités relatives. Selon lui le pourcentage de matière sèche du grain répond
à
cescritères. Nous apportons à la discussion une notion nouvelle, l' humidi té
realtive qui est définie comme suit: le pourcentage d'humidité d'un
hybride ou d'une lignée pris à~ un intervalle fixe de jours après l'apparition des soi.es, comparé au pourcen'l:agt d'un aU,tre hybride ou lignée pure (Hi.llson et Penny, 1965). Ce confept d'humidité relativ~ est étroitement relié au " concept de maturité relative (Snelling et Hower, 1940; Mi.les, 1940;
Dessureaux ~ al., ,1948) qui pourrait être définie comme la maturité \d'un
hybride ou d'une l.i,gne pure comparée
à
un autre hybride ou J..ignée pureà
unstade donné de la croissance. Cette notion permet d'avqir un point de
•
repère qui tient compt~ des variations saisonnières. Les mesures de la maturité relative sont nombreuses et varient avec les auteurs. Pour
Snelling et Hower (1940), par exemple, le contenu en matière sèche Glu grain
à la récolte s'avère un indice peu fiable alors que le nombre de jours du semis à l'apparition dès soies semble satisfaisant.
Donc on constata.. qu'il Y' a vraiment deux approches pour, définir la
mat~rité.
Un groupe s'attache au produit fini, c'est-à-dire, le pourcentage d'humidité du grain. Kaewer (1954), par exemple, s'appuyant sur la corrélation ~égati ve entre la vigueur de la ,pousse et le pourcentage d' humidité à la récolte conclut que du point de vue du producteur de semence_ le pourcentage,
d ' humidi té à la récol te- devi en t le cr itère 0 fondamental. L 1 autre groupe
se basant sur le fait que la translocation de matière sèche dans le grain cesse lorsque l'humid{té du grain est de 40 pourcent (Rather et Martson,
1940), considère le nombre de jours @u semis à ce stade comme critique.
Cependant cette notion i,mplique que le sèchage
d~
g;ainprocèJ~
de façon uniforme et similaire pour chaque hybride ou lign~ N~anmoins 1·
(
c
Il
..
4
Chase (1964) rapporte qu'il Y a de-s différences entre les rythmes de p.erte d'eau des ,différentes lignées. En comparant des hybrides hâtifs ayant le-mpJn~ pourcentage d'humidité du grain à la récolte, i l note' que les hybrides
..
de la même classe ayant fleuri tard perdent leur eau rapidement et donnent
D
de meilleurs rendements que ceux qui fleurissent tôt et sèchent lentem~nt.
----Hillson et Penny (1'965) suggèrent que le meilleur indiée de la - 1
maturité relative reste le rythme de séchage. Ils ajoutent que tout système.
de
c~tssifi~ation
utilisable doit être fondé sur l ' appari tion des soies,la -maturité physiologique et le rythme de perte d'eau. Ils reprennent en essence les critères .de Mayerl'';;à962)
à
l'exception de la date de sortie des soies. En effe:, pour ce~~rnier, ~'écart
e?tre le semis et 80 pourcent des panicules sorties est plus valable que l ' appari tion des soies. De plus,Gun et Christensen (1965) observent que les variétés à sortie hâtive des
"
soies et
à
haute teneur-- en eau n'indiquent pas toqjours un rythme de séchage lent pas plus que l'inverse ne s'avère vrai'.On con~tate qu'un~ nouvelle dimension s'ijoute
à
la discussion,l'importance de la floraison mesurée par la sortie de la panicule ou des
soies. \
L'utilisation de la floraison pour définir la maturité relative est peut-être le critère qui fut le premier utilisé. En effet, Alberts (1926)' rapporte que les variétés hâtives nécessitent moins de temps du semis
à
l'apparition
d~s
soies que les variétés tardives. De Jlus, il observe 1 que l'écart entre{I
apparitiàn des soiesl et le moment où le grain est denté est de 40 jours tant pour les variétés hâtives que tardives. Wolfe (1927) confirme les IObservations de Alberts (1926) et suggère que les dates de sortie des panicules et des soies sont des indices fiables pour définir la maturité.Miles (1940), pour sa part, considère que .l'apparition ~Ts soies seulement n'est pas un indice satisfaisant pour défini!: une lignée comme
Il
'
..<
,
C'
11
o
-
.,.
-Il
5 "tardi ve ou hâtive. Il note cependant en mesurant la maturité relative, que chaque jour de délai dans l'apparition des soies ret~rde la maturité physiologique (38 pourcent d'humidité) de 1.6 jours et l'obtention de 20 pourcent d 'hÙmidité de 2.1 jours. Il semble donc qu'if Y a une
i
relation positive entre le nombre de jours du semis
à
l'appar-ition dessoies et la perte d'eau. /'\
Dessureaux ~ al. (1948) remarql,1ent par contre que cette relation n'est pas toujours positive. En effet, ils observent que d'ordre général l" les· lignées qui fleurissent tôt ont tenC!ance
à
mûrir PlJS rapidement que celles qui fleurissent tard. Cependant, ils notent qùe le rythme de1
séchage est l~nt pour certaines lignées à floraison hâtivre et rapide chez d'autres à floraison tardive.
Di
M~~tino
(1957) observe le même phénomène et cite l'exe~le
d'un1
hybride ~ui fleurit sept jours plus tard qu'une variété témoin mais se développe plus rapidemerlt par la suite. Rumbaugn '(1959) rapporte
à
la suite de ses études que le nombre de joursà
1 'apparition. des soies s'avère un critère imprécis.Il semble donc qu'il n' y a pas une seule mesure comme Aldrich (1942) souhaitait, même si certains succès furent obtenus en n'utilisant qù'un critère. En eff~t, Giesbrecnt (1961) recommande Morden 88 au ~itoba sur la base du pourcentage en ea ~/ de l'épi à la récolte.
'"
Comme mesure multiple, Oberdoz et Muller (1957) suggèrent le nombre de
1 jours du semis à la sortie de la panicule e~ le pourcentage d 'humidité du
grain à la récolte COmme critères de classification internationale des lignées de maïs. Cependant, Hallauer ~ al. (1962) rejettent cet,te idée parce que disent-ils, ces mesures varient d' un endroit
à
l'autre et ci 'une..
,
annele a l'autre. ~o
1 !.
,..
..
• ,,-- r
. ( )(
, \(
- ,,- " / 6Il semble y avoir tellement de contradictions parmi les auteurs, que
/
nous avons donc décidé d'étudier le développemen~ phénologique pour
essayer de déduire certaines relations aVeC la maturi té dans des conditions
\ ~
doanées.
-,
\
2. Développement phénologique o
Shaw et Thom (1951) avancent que~la mafurité est déterminée avant la sortie de la panicule. Ainsill 'expres.sion génétique de la précocité
s'exprime du semis
à
l~ sortie de la panicule. Ils définissent trois(a) semis - levée, (h) levée - panicule,
~ases dans la croissance du mais:
•
U(~icule - app~ition des soies.
""-s'appuyant sur l'hYPOthèse que les différences entre las lignées se. sont Ils retiennent ce dernier stade en
exprimées à ce moment-là.
-:Pour sa part Hanway (19)53) définit de façon plus précise les phases de Shaw et Thom (1951) et les divise ainsi: (a). semis - levée, (b) levée - 4 feuilles, (c) 4 - 8 feuilles, (d)' 8 - 12 feuill~s, (e) 12 - 16 feuilles,
(f) 18 feuilles..,. 75 pourcent des soies, (g) 12 jours, (h) 24 jours, (i)
\
36 jours, (j) 48 jours, (k) 60 jours"-a~ès l'apparition des soies.
Pour l'étude que nous entreprenons nous étudierons lef phases selon la méthode Hanway (1963) avec 9,es mOdific~ti.,gns pour nos ,:onditions (voir
Matériels et Méthodes, e~périence 1).
3. Carac"tères morphologiques
/
1
Il est
~endant
impossible d'étudier les sta..des de.
développement~
sans étudier 1 e;:; différents caractères morphologiques qui' s 'y rattachent. En effet, Kuleshov (1933) suggère l'utilisation du nombre de feuilles comme critère de classifi.cation des hybrides américai!ls et russes. Nozzolini(11~63) Observe des corréla tians posi ti ves de 0.94 et 0.92 entre le nombre de f'~uilles et la maturité. Il suggère que la maturité peut être prédite
,<1
avant la floraison en faisant\ des coupes longitudinales. de la plante après ~
1 f-I
,
[
' \ 1.. '_l'initiation de la première feuille, chaque feuille additionnelle accroît
de deux
à
tro~s jours la maturité. qne critique ~u~ nous po~vons formulerest qu'il fonde son étude sur des corrélatio~s qui peuvent être sujettes
à
interprétation. - Cependant, Chase et Nanda (1967) 1 notent eux-aussi des
corrélations positives très élevées entre le nombre de feuilles et la maturité, les hybrides hâtifs possèdant moins de feuilles que les hybrides tardifs.
Pour expliquer cette relation on peut se référer
à
Eik et Hanway(1973) qui supposent que les facteurs affectant le nombre de feuilles en influençant l'initiati9n de la panicule ou le rythme
feuilles avant la panicule. Hesketh et al. (1969) \
\
cette hYpOthèse en observant que le nombre de reuilles est relié la sottie de la panicule, au poids de la plante et au
coefficient (ci-après ÇF)
à
l'apparition de la panicule.I~
~
de feuilles est aussi étroitement relié
à
la maturitépourquoi ne pas 'utiliser' comme uniqûe facteur de sélection? La raison
en est simple
t>
isque le nombre de feuilles varie avec la date de semis,l'endroit, la saison et l'environnement selon Sieglinger (1936). Observation
qui fut reprise par Eik et Hanway (1""973) qui reconnaissent que le nombre de
feuilles peut être influencé par l'enviro~nement, particulièreme~t par la
température qui le fait accrottre lorsqu'elle augmente.
Il semble cependant incontestable qU'11 y ,a une relation étroite entre
le nombre de feuilles et; la maturité '(Yoshida 1973). Cette relation n'est
pas linéaire car les feuilles sont initiées plus lentement au niveau du
tissu méristématique immédiatement après la levée que plus tard, l'intervalle
entre l'initiationlde chaque feuille diminuant au fur et
à
mesure que laplante croît (Quinby 1973).
\
Un autre facteur relié au nombre de feuilles qui semble important
à
étudier' es't le CF. En effet, i l peuet servir de critère de sélection des
, -- r
(
1
(
1
o
\'.,
;'--
--,,---,àe déveÜ:>ppement (Derieux et al. 1973).· Qui plus est, Hunter
~
al. (1970)notent <ilue pour les variétés hâtives le CF le plus élevé est de 2.9 pour
un peuplement de 72,000 pieds à l'hectare. Cette valeur de 2.9 est de loin
inférieure à toute valeur obtenue à la même densité pour des vari~tés
tardives.
Cep~ndant, le CF varie trop en fonction du peuplement pour être'
utilisé co~e 'critère unique (Namez et Kamprath 1969). Néanmoins, quoique
difficile à mesurer, le CF es~ un critère intéres~ant à étudier (Wallace(~ al.
\ 'j
1972) •
j
,Parmi les autres caractères morphqlogiques secondaires mais intéressants
à mesurer, notons d'abord la hauteur. Selon Cross et Zuber (1973) l~ hauteur
de la plante à maturité est un excellent indice du nombre de jours du semis
à la floraison. De plus, Hallauer et Sears (1973) constatent que la relation
entre la hauteur de la plante et l'écart entre le semis et la sortie de la paniculeiest inversement proportionnelle. Cette corrélation négative fut observée par Kempton (1924) dans une étude quantitative des caractères morphologiques du mais. Par contre, Oberdorf et Muller (1957) contredisent cette corrélation et rapportent que 'la hauteur n'est pas associée avec la date de sortie de la panicule.
Enft' n parmi d'autres caractères morphologiques étudiés en fonction de
la matur té, mentionnons la longueur de l'épi (kempton 1924) la hauteur des
épis et le nombre d'épis (Aldrich 1942).
4. La génétique de la maturité
Selon certains aut~urs (Giesbrecht 1959; Hesketh 1969; Derieux ~ al.
1973) il n'y aurait que qaelques gènes impliqué dans la détermination
de la précocité. Qui plus est, la précocité est dominante sur la, maturation
tardive. Ainsi l'hybride produit par un croisement tardif x hâtif possède
la précocité du
paren~
hâtif et une augmentation de la taille et de,la, - r
-
".(
c
.
\() /
9que
~onfirmer
yne pratique courante,rap~orté~
par Eckhart et Byron (1940), pour produire des lignées à courte saison, on croisait une lignée hâtive ,e.t upe tardive.Leng (1949) note que chez le maïs le développement végétatif à partir
du semis à l'apparition des soies se fait en deux phases; la première période dite végétative caractérlsée par l'élongation du point de crois-sance, la deuxième jusqu'à l'apparition des soies, période où les structures
reproductrices se développent. Il constate que les lignéès pures ont des
.
..
"patterns" de développement héréditaires et caractéristiques.
L'influence des l~gnées parentales dans la-production d'hybrides
devient primordiale. En eff~t,·Whaley et al. (1950) dans une étude sur la
croissance et la morphologie de d,eux lignées pur~s et leurs hybrides,
observent ~r ces derniers l~s caractères suivants: une augmentation du
poids total, un ~ccroissement de la tige, des racines, de la longueur de
la feuille et du CF. Ils\\lconcluent que les hybrides gagnàient ~u départ
par rapport aux parents et gardaient ce gain jusqu'~ maturité. De plus
la croissance des-feuilles étai~ distincte pour les lignées pures alors!
que le nombre de feuilles était ~ntermédiaire pour les hybrides et que les
feuilles des hybrides étaient plus hâtives et formées plus rapidement.
Pour leur part, Hillson et Penny (1965) notent de très grandes
différences dàns le séchage des croisements rapide x rapi~e, rapide x lent,
lent x lent, mesuré 75 jours après l'apparition des soies. Une lignée
pure, 3~-11, démontre un effet dominant sur les autres en fonction du
rythme de séchage. ~ls concluent qu'il y a des différences sigrlificatives
1
entre les croisements en ce qui a trait à la perte d'eau et que les ligntes parentales ont un effet sur le Jythme de perte d'eau.
\
Si on essaie de séparer l'effet dû à l'environnement de la composante génétiqUei\il semble que les lignées faibles qui ont peu d'effet additif sont influenéées plus fortement que les autres, tout au moins en ce qui a
trait aux variables suivantes: la date d'apparition des soies et la
hauteur de la plante (Darrah et Hallauer 1972).
r
(
/
10
De cette 'revus de litJérature on peut dégager les questions suivantes: -quellé est la relation entre les différents stades de développement et la
maturité?, comment ces stades de développement sont-ils transmis
génétique-ment?, est-il possible d'utiliser ces différents stades de développeme~t
comme critère de sélection dans un \programme-- d' amélioratiol1 du mai's visant
o \
à pro~uire des hybrides hâtifs?, '
, o a , , }
,.
1
(
o
-.,
'1 EXPERIENCE l ~Le but de cette expérience est de définir la relation entre le rythme
.'
~e croissance aux stades végétatifs et ..- p~oductifs et la maturité chez le mais.
l~ Matériel et Méthode
1
1.1 Le facte~r étudié
D
Notre objectif est d'analyser la relation entre le développe-ment phénolo~ique du màis et la maturité dans les 1ondit10ns
locales. L'effet étudié est la maturité que nous mksurerons par le pourcentage d'humidité à la récolte (ci-après nammé PHR). Nous avons choisi cette mesure parce qu'elle s'avère celle qui représente le plus la récllité agricole. En effet, le PHR détermine la facilité d'entreposage sans ;isque de
pert~,
ainsi que la capacité de germination ..
;
\
Nous sommes conscient de l'importance du rythme de séchage, cependant ce facteur nécessite une approche expér1mentale particulière, de plus il n'influence pas,directernent la production du mais puisque cette dernière
!
s'attache au pro~uit fini.
Nous avons fesuré le PHR à la même date, le 20 septembre 1976, pour chaque lignée. ~oute personne ayant composé un tant soit peu avec les conditions du Québec, constatera que cette date est quelque peu hâtive.
O·
Nous l'avons choisi pour les motifs suivants: (a) ayant semé le 24 mai cela nous donne une' p~ri,ode de croissance de 120 jours qui représente la
période de croissance moyenne au Québec, Cb) à cause de la disponibilité
1 #
du personnel de soutien et del'é~~ipement de laboratoire, plus spécifique-ment des séchoirs à grain de la station expérime1tale.
, - r
(
(
- ".
12
~
po4r mesurer le PHR nous avons procédé comme suit: après avoir inscrit le nombre d'épis par plante dans chaque parcelle, tous les épis de la parcelle étaient placés d~s un cabaret étiqu~té et éta~ent ~sés. Le choix de sécher l'épi au complet fut basé sur les expériences de Brown et Rossman (1954) qui rapportent que si Il'on ,a un ~rès haut niveaud' humid~ té, l'aJustement sur le poureen,t'age d' hUIlUdi té de l'épi et des
"
grains est plus précis que l'aJustement sur le pourcentage du grain seulemen t. Les cabarets fur en t'ensui te placés au séchoir à une tempéra-tureconstante,76oC. A tous les quatre heures les échantillons furent pesés jusqu'à ce qu'il n'y ait plus de perte
d'ea~1
c'est-à-dire,q~'il
;,
n'y ait plus de variation dans le poids. était de 0 à 0.5 pourcent.
A ce point l'pumidité de l'épi
•
!
Nous étudions la maturité, comme facteur lié au développement
•
~orpho-physiologique de la plante. Nous analyserons donc ce facteur'
Jo
fonction de ses liens avec les différents stades de croissance, stades que nous définirons sous "variables mesurées."1.2 Matériel expérimental
l ,
A l'été 1975 nous avons pèmé 448 lignées pures d~ maïs, sans répétition. Ces 448 lignées comprenaient des l~gnées reCOnnues comme hâtives
sélec-tionnées par le Dr Brawn, des l~gnées id en ti fiées Q, produi tes à r~aedonald
,
College, des lignées importé~s,de France et de Bulgarie, ,des lignées provenant de la station fédérale à Ottawa identifiées C.O., et enfin des lignées résistantes au froid provenant des stocks du Dr Brawn.
-A partir de ce matériel nous voulions des lignées précoces homogènes.
\ '
~ous ~vons donc sélectionné 36 lignées (voir Tableau
f>
offrant un écartrelatif de maturité sans pour cela être inadaptées
au~
conditions du milieu. Pour cette raison les lignées qui exigeaient plus de 3200 unités thermiques(ci après UT) ne furent pas sélectionnées.
,
, -..1'
- ..,.,~
Les Itgnées utilisées
•
13
ne sont pas représentatives de la population "mais" au sens large mais au sens restreint elles le sont de la
population "mal.s québécoi~." Elles offrent un éventail assez juste du
mâis pouvant être cultivé au Québec.
" ,
Les lignees furent semees le 24 ma~ 1976 dans le champ 910 au centre
de recherche Emile A. Lods de ~cdonald Co+lege. En 1975 ce champ avait
servi à des essais de céréales. Pour le désherbage on utilisa un mélange
d'Atrazine à deux kilogrammes et deni
à
l'hectare et d'Alachlorà
cinqlitres soixante à l'hectare. Le programme de fwnure consis.ta én quatre
cents cinquante qua~re kilogrammes à l'hectare de 5-20-20 avant le semis
et un
ren~haUSSage d~
deux cents vingt huit kilogrammes à l'hectare de34-0-0. Toutes les lignées furent semées le même jour.
1.3 Dispositif expérimental
1. 3.1 Dispos1f statistique
Nous avons choisi un dispositif de blocs aléatoires où chaque lignée est considérée comme un traitement.
1.3.2 Nombre de blocs
.~ Selon Henry et al. (1942) 1 le nombre de plantes nécessaires pour
déterminer le PHR est fonction du nombre de variétés et du n~~e de
blocs. Pour le nombre de lignées que nous avions deux blocs auraient
suffi,
cepen~ant ét~nt
donné que nous voulions l'erreur expérimentale laplus précise possible pour les variables mesurées nous avons choisi
quatre blocs comme étant
~e
nombre qui corr:spondait le mieux à noscritères expérimentaux et pratiques.
.\
1
•
C
L-o
~_.@i.5 t -14TABLEAU 1." Liste de, lignées de l'expérience l
Lignée~ résistantes au froid
(produi tes par R.l. Brawn)
73-017 16-7P 73-018 76-1P
•
73-026 76-21P 73-034 76-2P 73-035 76-l9P 73-035A r- 76-5P 73-036 ~ 76-20P 73-040A ~6-16P 73-047 76-8P Lignées Q(produites à Macdonald College)
QI 76-33P
--Q165B ;16-36P QJ:84 76-29P Q204 76-28P Q205 76-32P Q219 76-34P Q221 76-35P Q225 76-31P Q226 76-30P/
~•
Lignées hâtives(produi tes par
R.
l . Brawn)73-278 76-9P 73-355 76-3P 73-~60 76-1SP 73- 68 76-4P 73-369 76-23P 73-370 76-11P . 73-373 76-24P 73-383 76-6P 73-387 76-12P 73-393 "- 76-10P 73-395 76-13P 73-398 76-14P 73-401 76-17P 73-406 76-1BP
Lignélls provenant d'Ottawa
C.Q-109 76-26P
C.Q-125 76-27P
Lignées provenant de France
Syn SI<G Syn GeR 76-22P 76-25P \ ~ o J, i
1
~ ", - r
- ...(
- - i , ro
1.3.3 Taille et description de 1 l'unit~ expérimentale , 0 15Chaque uni té 'expérimentale a 3.6 mètres de longueur avec l mètre entre chaque unité expérimentale. c Aux extrémités du champ servant à
l'e~p~rience i l y a trois lignes de Acco DC 103, qui servent de bordure. A chaque unité est assignée une lignée tirée au sôrt. A l'intérieur de chaque unité huit plantes sont semées avec 30 cm entre chaque plante. Aux deux extrémités de chaque uni té nous avons semé. deux plantes de
c
Acco DC 103:- Pour chaqu"e plante semée on utilisa deux graines et on a
1 l,
procédé à l'éclairèissage lorsque les deux graines germaient. Chaque
J 1
unité expérimental~ portait un numt!r~, YXX, y représentant le numéro du bloc soit de l à 4 et XX l e num~ro de l' uni. té' ..
1.4 Va~iables mesurées
"
,
.
Nous eXp?serons les différentes variables mesurées en expliquant les motifs qui nous ont amené à les sélectionn,Or et le mode de mesure
,",,''''
de ces variables.
(a) La levée
C'est 'le point le plus imRQrtant à définir' car toutes les données subséquentes sur le développement morphologique' s
''Y
réfèrent. Shaw et Thom(1951) ayant recensé 10,000 plantes définissent la levée comme le moment où une portion de la plante est visible. Hanway (1963) utilise la même définition
d~s
une"etud~
des stadeJ de croissançe du mais...
Dans cette expérience un~ plante sera co~sid~rée comme levée lorsque l'épicotyle sera visiblè alors la date sera enregistrée.
(h) Les Çlifférents stades de ~éveloppement . étucliés
,
.
Comme nous avons expliqué plus haut, nous nous sommes basés
sur
les études de Shaw et . Thom (1951) et Hapway (1963) pour définir less~ades de développement que nous dési;ons étudier.
1
'.0·
-
.,
16
Le' premier~ stade est la levée. ,C'elle-ci n'est pas un stade en soi
mais plutôt un condition nécessaire
à
la croissancer °L'écart entre le)
~ semis et la levée peut-être considéré comme une phase du développement
et i l est 9aractéristique de chaque lignée. Il est évident que cette
phase est fonction des facteurs du milieu mais toutes choses étant
\
égales les différences" si différences i l y a, entre les lignées peuvent
être attribuées a~ caract~ristiques inhérentes à la lignée (Shaw et
Loomis 1951).
L,a seconde Jhase du
~~
sortie de la panicule
Il
dé~~loppement étudiée est de la levéé à la
(Shaw et Thom 1951) dite phas'e végétàtive. Nous
l'avons subdivisée selon le nombre de feuilles, en adaptant les critères de Hanway (1963) aux conditions locales. Nous obtenons les phases
suivantes: (i-} levée à six feuilles, (i,i) six à neuf feuilles, (Hi)
°neuf feuilles à la sortie de la panicul~.
Avant d'aborder CeS phases il est important de décrire quand nous
considérons une feuille développée. Une feui~le-sera comptée lorsqu'elle
~
aura émergée de 1~ gaine de la 'feuille précédente et qu'elle aura
, '\ , ~
commence a se derouler.
Cette méthode qUQique
suivant~: (i) "il est très
----empirique s'avère adéquate pour
f~s
simple d'identifier
1
feuilleà
cemotifs stade!
(ii1 étant donné que l'expérimentateur est la seule personne à
prendre~
les données, cela assure une uniformité dans le décomptei (iii) à cause
de cette uniformité l'erreur est éonstante, donc on peut se servir des
résultats comme base de comp~aison. -De plus,1 il- nous a semblé que
,
c'était~'la méthode la plus efficace compte tenu de l'ampleur de l'expéri-ence.
Vu
qu~
les,!l~ées
utilisées au~uébeG:,
ont rarement plus de 12, \feuilles, en di~isant le développem~t des feui+les ainsi: levée - six
feuill~s, 6-9 feuilles, 9 feuilles - nombre final de feuilles - on obtient
..
"
1 ,.
, - r
c
o
\1 1 .'Il
17une vue chrOnOlOg~qUe du développement du mâis. De plus, cette division
nous permet de voir où s'établit la différence entre les lignées.
La première phase levée - 6 feuilles (ci-après'phase 1) - consistait
à mesurer~le nombre de jours entre la levée et l'apparition de la sixième
feuille. Il aurait été intéressant comme nous verrons plus tard, de
.
diviser cette phase en deux, soit levée - 3 feuilles et 3 feuilles - 6
feuilles, mais la difficulté "de pouvoir colliger les du
nombre de plantes à mesurer et de l'écart trop court e
divisions rendaient cette approche inaccessible à
1
La~euxième phase, 6-9 feui~les (ci-après phase II) est définie
.
comme .. le ·temps entre ~ 'apparition de la' sixième et la neuvième feuilie.
ce stade la plante est assez haute pour faciliter la mesure.
La troisième phase du développement que ~ous mesurons est la phase
-
..
19 feuilles sortie de la panicule (ciaprès phase III) ou 9 feuilles
-A
nombre final de feuilles. En effet une fois l'inflorescence initiée, il
n'y a plus de développement de nouvelles feuilles. Ceci nous amène
à
~onsidérer le nombre de feuilles comme un indicateur de l'â~e physiologique~
de la floraison (Duncan-Hesketh 1968). Cette dernière phase est fonction
Il
de la sortie de la rani cule. Nous considérons la panicul-e sortie lorsqu: elle forme une croix mais avaht qu'on ne voit les anthères.
Nqus avons choisi la sortie de la panicule parce~qu'elle coincide avec
la cessation de production de feuille et qu'elle est reliée à la maturité
selon certains auteurs (Albert 1926;'Wo1fe 1927; Oberdof et Muller 1957).
Lorsque la panicule est apparue 0 nous ,comptons le nombre d~ feuilles
car il est considéré parmi les caractères morphologiques étudiés. En effet
plusieurs expériencès ont démontré une cOlréla~ion positive en~re le nombre 0
.de feuilles et la'"maturité (Nozzolini 1963).
!
.
'1
(
o
.... 11..1
18
uri autre caractère morphologique retenu est le CF. Il semble y
avoir une certaine corrélation entre le CF et la maturité (Hunter ~ ~.
1970). De plus, Derieux (1973) conclut qu'il
peu~
servir de critère desélection pour la précocité chez le'mâis. \
Etane donné le nombre de plantes à 'mesurer pour
le~ deux-éxpérienc~s
. /
.
soit 3000 plantes, nous avons choisi la méthode de "nerieux (1973). Il '
sagit ~e mesurer le CF de la cinquième feuille ce qui ne nous donne pas
le CF 1 de la plante mais son effet relatif. Le CF / de la cinquième
feuille fut mesuré à la phase III.
La dernière varia;ble mesurée fut le nombre d'épis à la récolte.
Etant donné l'importance du stade de remplissage du grain (Shaw et Loomis
1951; Hanway 1963) il hOUS apparatt important de voir cO~Tnt le nombre
J'éPiS
à remplir influence la maturité finale.\. 1.5 Conduite et déroulement de l'expérience-?
Les données furent prises quotidienneme~t par l'expérimentateur
,
jusqu'à la sortie de toute,s les Péficules. Pour les phases l , II et III
les dOnnées\\fUrent enregistrées en nombre de jours. 1 De pIus les données
en jour Jurent transposées ;én° unité de chaleur selon le système d:Ontario
qui d'après Mederski et aL (1973) s'avère très précis. Pour la trans,:,,\
formation on utilisa la formule suivante:
uni té de chaleur
=
\
{1.85(max - lOOe) O.026(rnax - lOOC)2} + {min - 4.40C}
2
max température maximum de la journ~e (24 h) exprimée en degré
celsius
IlÜn
\ 1
température minimum
âe
la journée (24 h) exprimée en degrécelsius q \
\
'ri
(
c
()
.,
~. '\ 19 1"La ~eco1te fut effectuéè le 20 septembre. Chaque parcelle fut récoltée individuellement et le poids frais enregistré. Les boîtes contenant les échantillons furent ensuite séchées tel que décrit plus
haut.
,
2. Résultats et Discussion
"1
,2.1 Analyse dés résultats
2.1.1 Le nombre de plantes levées
1
La première étape dans l'analyse des résultats
consist~
à voir s'ily a une différence de germination entre les lignéBs. En effet à cause
de l'âge des semences (1973) il apparaît important de vérifier cette
différence. Notre hypothèse est la suivante: s'il n'y a pas de
différence significative entre les lignées en ce qui a trait au nombre
de plantes tevées, nous pourrons analyser les autres résultats en utilisant
la moyenne des lignées.
\-_ lb
Nous avons obtenu pour \l'ensemble de l'expérience 76.8 pourcent de
germination avec un
coeffic~eht
de variation de 21. 9 pourcent •.~i
pn (regarde l'analyse de la variance pour le nombre de pl~ntes levees nous
remarquons qu'il n'y a pas de différences signifi~atives èntre les blocs
.1
et entre les ligné,s (Tableau 2). Nous pouvons tlonc poursuivre l'analyse
des résultats en
~tilisant
la moyenne de chaque parcelle plutôt quechaque plante prise individuellement.
TABLEAU 2. Analyse de la vari~ce de la variable "nombre de plantes
l~vées" Source Bloc l' -~ignées
"
Erreur NS - non"
1\ dl ~ 3 35 105 significatif _s.e.
19.6875 198.6875 497.1750 Valeur F ;1 •. 39 NS 1.20 NS•
o CI/
(
(:
\1
, "
. 20
'l. 1.2 Analyse des variables mesurées
(a) La levée •
L'analyse de la variance démontre qu'il y a une différence hautement significative entre les lignées pour ce qui est de l'écart entre le semis et la levée-.
TABLEAU 3. Analyse de la variance de 1, variable .. semis a , la levée
Sou'rce dl S.C. Valeur F
'~
FR > FBloc 3 5.13 1.89
J
0.13Lignée 35 72.1B 2.27
\
0.01**Erreur 105 95.39
~*Significative au niveau de .01
. Il n'y a pas de différence entre les blocs et d'autre part le ,. coefficient de variation est de 9.15 pourcent. La moyenne est de 10.40 -jours avec un maximum de 12.75 et un minimum de 7.99 et un écart de 0.95.
L'hypothèse du test F, qui veut que la différence entre les lignées soit due aux traitements et non au hasard, est vérifiée par les résultats du test (hautement significatif) et renchérie par le fait que le
coefficient de variation est faible. De plus, l'écart entre le maximum
et le minimum indique clairement la différence entre les
l~~nées
au"
début du développement.( )
.
, ,\
b Lès diverses phases du developpement
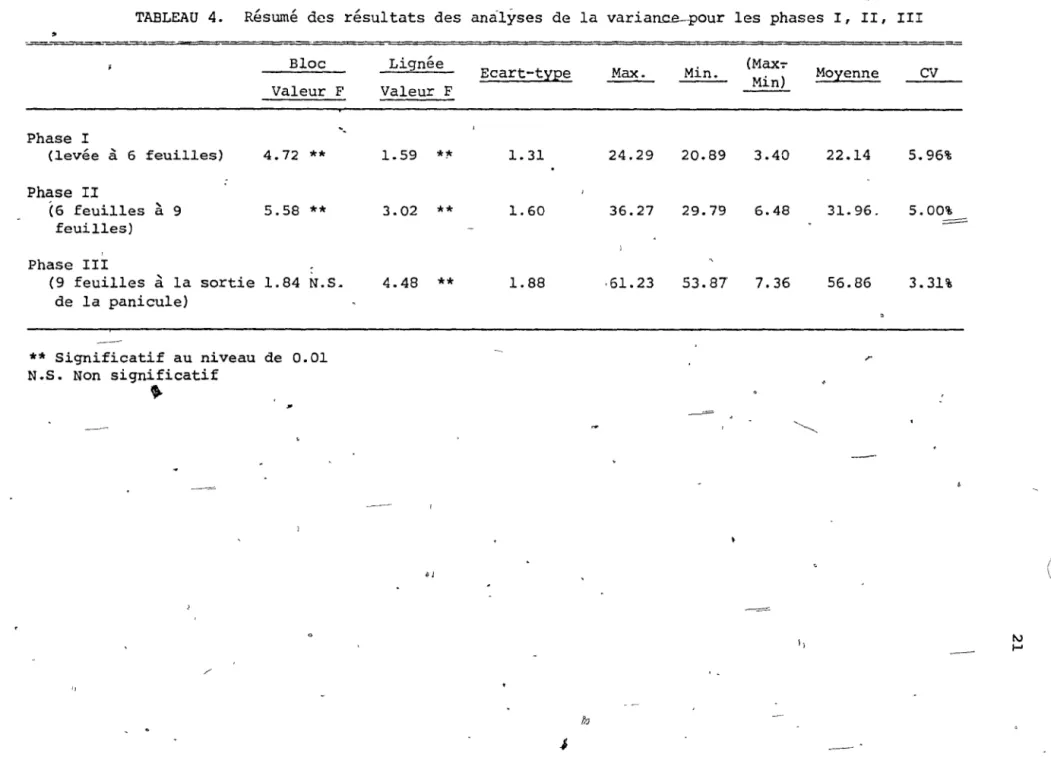
Si o~ se réfère au tableau 4 résumant les analyses de la variance pour les phases l, II, Illon constate d'abord qu'il y a une différence hautement significative entre les lignées pour les trois phases. II y a aussi des différences entre les :blocs.
)
phases le coefficient de vari9tion est
/
Il est à noter que pour les troi~ très faible, et on
pe~t
donc-
I~
,..
~
---TABLEAU 4. Résumé des résultats des analyses de la variance-pour les phases l, II, III
~ Bloc Valeur F Phase l
-,
(levée à 6 feuilles) 4.72**
Phase II (6 feuilles à 9 5.58**
feuilles) Phase III (9 feuillesà
la sortie 1.84 N.S. de la panicule) ** Significatif au niveau de 0.01 N.S. Non significatif•
/ ' "tftiI'7û~~iÎ~~~À-~~ <,' Lignée Ecart-t:iEe Valeur F 1.59 *1< 1. 31 3.02**
1. 60 4.48 ** 1.88 6l JMax. Min. (Max-:- Moyenne CV
Min) 24.29 20.89 3.40 22.14 5.96% 36.27 29.79 6.48 31. 96. 5.00%
=
,61.23 53.87 7.36 56.86 3.31% r--=
~ --= 1) fb..
t\J....
(=.
lA...
'II1
..
u
(
0
22
cO~~lure
que l'expérience s'est dérollée de façon uniforme et' que les résultats'sont représentatifs du cho~x.
des lignées et ne dépendent pas de fa,cteurs extérieurs à 1,' expérience.-
.
\
,Un au't.re ; i n t
à
'souligner est l'écart entre les maxima et minima pour les trois phases. On peut déduire ;ue;la diff,rence qui existe entre les lignées va en s'accentuant jusqu'à la sortie de la panicule. Ceci s'accor~e avec les conclusions de Shaw et Loomis (1951) qui•
suggèrent que le potentiel de précocité de l ' appari tion de la panict\le et que par t a
o \... • \
similaire pour toutes les lig~ees.
o
la plante s'exprime ~vant
suite le développement est
/
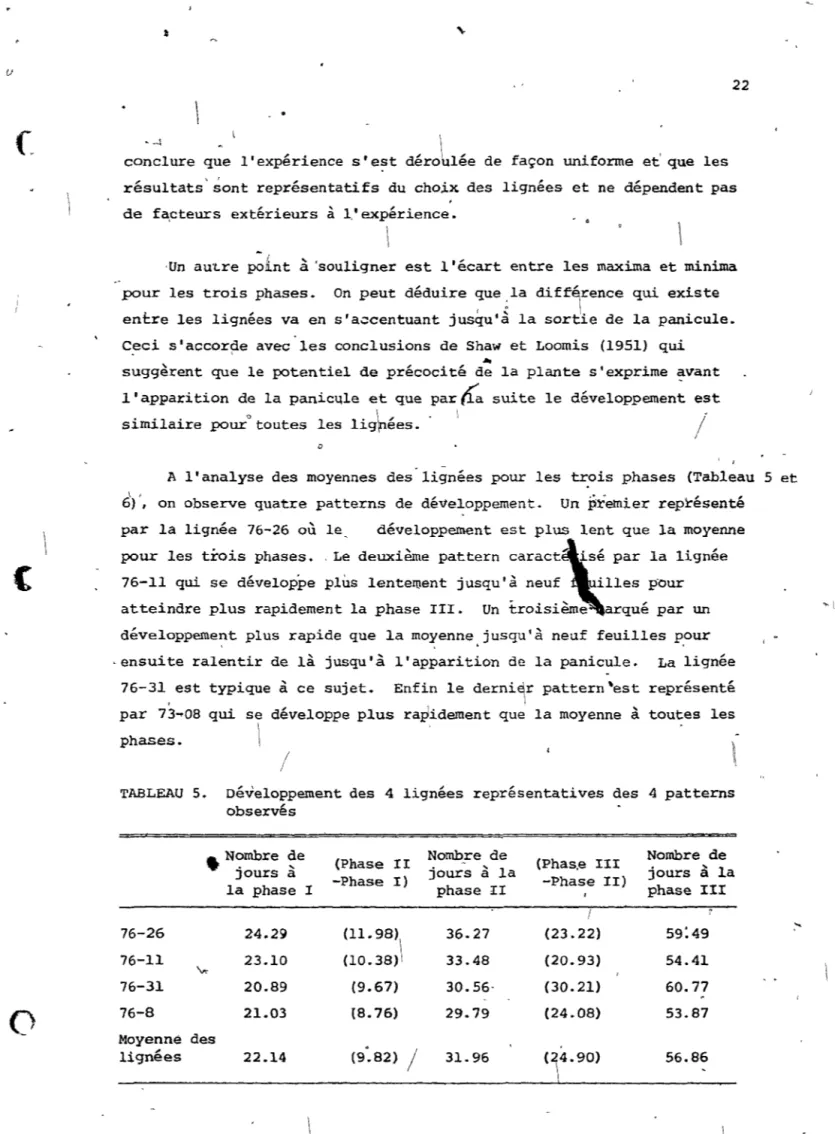
A l'analyse des moyennes des lignées pour les t~ois phases (Tableau 5 et
\ ,
.
6), on observe quatre patterns de développement. Un premier représenté par la lignée 76-26 où le, développement est plus lent que la moyenne pour les trois phases. ,Le deuxième pattern caractI'Sé par la lignée
76-11 qui se développe plus lentement jUSqU'~ n~u~, illes,~our atteindre plus rapidement la phase III. Un tro~S1eme arque par un développement plus rapide que la moyenne jusqu'à neuf feuilles p,our . ensuite ralentir de là jusqu'à l'apparition de la panicule. La lignée
76-31, est typique à ce sujet. Enfin le derni~r pattern~est représenté par 73-08 qui se développe plus rapidement que la moyenne à toutes les
\
phases.
/
'~TABLEAU 5. Développement des 4 lignées représentatives des 4 patterns
observés
• N<?rnbre ~e (Phase I I Nombre de (Phas,e III Nombre de Jours a
-Phase 1) jour"s à la -Phase II) jours à la
la phase l phase II phase III
/ 76-26 24.29 (11.98) 36.27 (23.22) 59:49 (l0.38)\ 76-11 23.10 33.48 (20.93) 54.41 'vt-76-31 20.89 (9.67) 30.56- (30.21) 60.77 76-8 21.03 (8.76) 29.79 (24.08) 53.87 Moyenné des lignées 22.14 (9.82)
/
31.96 (24.90)\
56.86:---..
"'1 -' i f ~ '~ ,j..
1
.,!
~
~ ... i:/1
,
- r
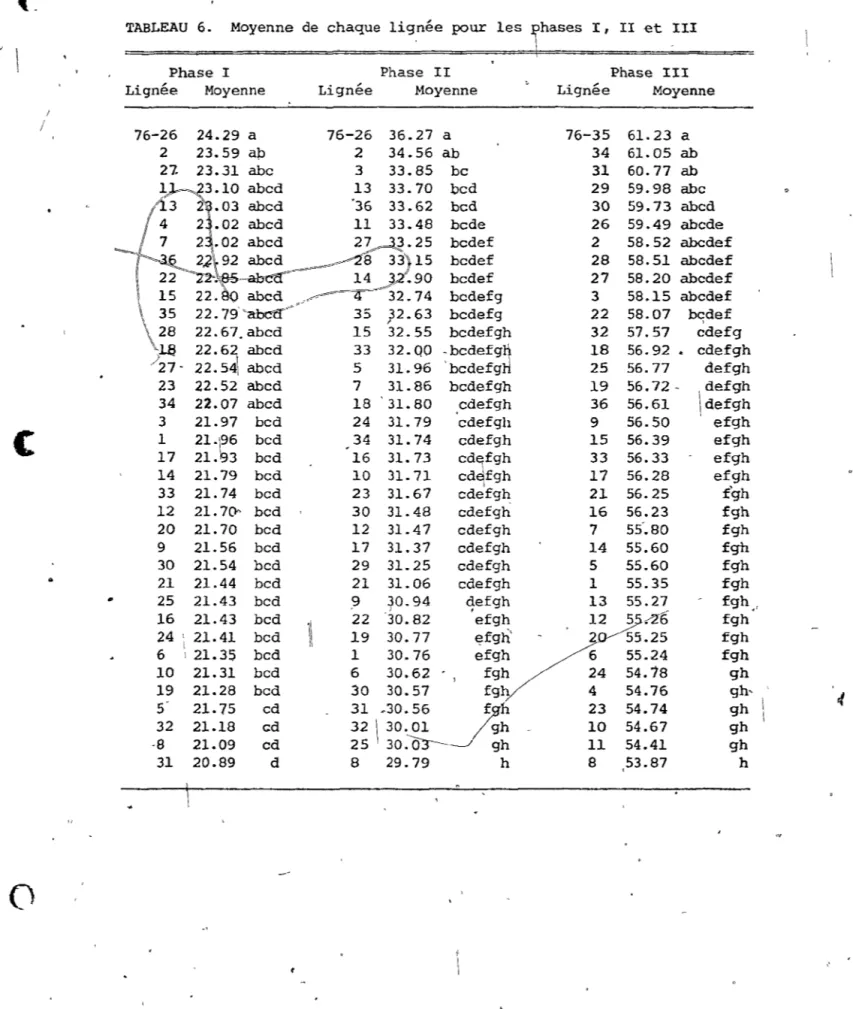
- ". / ' '\ 1 , :; 23i:'\.(-TABLEAU 6. Moyenne de chaque lignée pour les !haSeS I, II ~t III
~
\ Phase l Phase II Phase III
Lignée Moyenne Lignée Moyenne Lignée Moyenne
1 / 76-26 24.29 a 76-26 36.27 a 76-35 61. 23 a 2 23.59 ap 2 34.56 ab 34 61.05 ab 23.31 abc 3 33.85 bc 31 60.77 ab 13 33.70 bcd 29 59.98 abc abed '36 33.62 bcd 30 59.73 abed abed 11 33.48 bcde 26 59.49 abede
2~.25 bcdef 2 58.52 abcdef
~15
bcdef 28 58.51 abedef14 .90 bcdef 27 58.20 abedef
~/~"' 4 32.74 bedefg 3 58.15 abcdef
\ , 35 35 ;2.63 bcdefg 22 58.07 bqdef 28 22.67.abed 15 32.55 bedefgh 32 57.57 edefg
l.S 22.6~ abed 33 32.QO ~bedefg~ 18 56.92 • edefgh
/27- 22.54 abed 5 31. 96 bedefg 25 56.77 defgh 23 22.52 abed 7 31.86 bcdefgh 19 56.72 - defgh 34 22.07 abcd 18 31.80 cdefgh 36 56.61 \defgh
~~
3 21.97 bed 24 31. 79 edefgh 9 56.50 efgh
,1
(
1 21.~6 bed 34 31. 74 cdefgh 15 56.39 efgh17 21. 3 bcd 16 31. 73 Cd~f9h 33 56.33 efgh ~~~ 14 21.79 bcd 10 31. 71 cd fgh 17 56.28 efgh
"
33 21.74 bcd 23 31.67 cdefgh 21 56.25 f'gh 12 21.7Cf' bcd 30 31.48 cdefgh 16 56.23 fgh 20 21. 70 bed 12 31.47 cdefgh 7 55-.80 fgh 9 21.56 bcd 17 31.37 cdefgh 14 55.60 fgh 30 21.54 bcd 29 31.25 cdefgh 5 55.60 fgh..
21 21.44 bcd 21 31.06 cdefgh 1 55.35 fgh 25 21.43 bcd ,9 ~0.944
efg h 13 55.27 fgh .. 16 21.43 bcd 1\ 22 30.82 efgh 12 5.-26
fgh 24 1 21.41 bcd 19 30.77 ~fgh' 2 55.25 fgh ! 6 1 21.35 bcd 1 30.76 efgh 6 55.24 fgh 10 21.31 bcd 6 30.62 fgh 24 54.18 gh 19 21.28 bcd 30 30.57 fgh 4 54.76gh-4
5 21. 75 cd 31 ~30.56 ~h 23 54.74 gh \ 32 21.18 cd 32 \ 30.01 gh 10 54.67 gh -8 21.09 cd 25 30.03---- gh I l 54.41 gh 31 20.89 d 8 29.79 h 8.
53.87 h•
o
"
,'e~
, ~
,TABLEAU 7. Resumé des résultats de l'analyse de la variance-pour l~s variables: nombre de feuilles, CF,
nombre d'épis par plante et hauteur
, f
'---Nombre de feuilles
CE:,' de 1a cinquième
feuille ~
Nombre d'épis par plante Hauteur 1
/
Bloc Valeur F 0.27 NS 2.65*
3.71**
2.52 NS J. Lignée r Valeur F ,---tl.96**
7.81**
3.22**
,
10.10**
"~ . ) 1- .~, ',""" ~',',.,'-. "",~.,. ",.'.' r'I--~~~'"._ ....
-
... ~ ~ ~ Ecart-type ,0 __ 6290, -53.15 0.4781 10.15' c. .... 1;'..!n.~ ~i. MaximUm 13.46 572.02 4.00 126.06 ( 1 (.
Minimum 9.0 269.10 1.00 73.83 \ '"\~
Moyenne CV 0 11.47 - 5.48 371.58 14.30 1. 71 27.95 103.94 9.76G
... ".
,'"
,j::>'-"
-'II1
(
(
C\' \
/
25(c) Les caractères morphologiques
1
En étudiant les résultats des
an~lYSëS
de la yariance pour les divers carlctères morpho:ogiques (Tableau 7), on observe d'abord qu'il y a des différences hautement significatives entre lé~ lignées pour tous. L'effet des blocs n'est pas significatif pour le nombre de feuilles et pour la hauteur, et il est sign~ficatif pour le CF et hautement significatif po~r le 1Prnbre d'ép~s par plante..
Il faut aussi noter l'écart qui existe entre les maxima et minima
, \
des différents caractèresl
• Cet écart élevé et le faible coefficient
de variabilité confirment la signification des résultats de l'analyse de la variance.
(d) Analyse du facteur étudié, le PHR
/
\ Pour le PHR on const~te dl abord que la différence entre les lign~es est hauternjnt significative et celle entre les blocs est significative
(Tableau 8). Le coeffic~ent de variation est très fdible (8.06%). La moyenne est de 41. 76 pol1rcent, ce qui est près du pOift de maturité
> phys~ologique décrit par Hanr-'ay (1963). De plus on p~ut retrancher 2
- : 1
unités pour obtenir l'humidité du grain ce qui ramène 'le PHR
f
39.76%.Dans l" ensemble, on peut considérer que la plupaFt des lignéE7,~ étaient près de la maturité physiologique lors de la récolte.
TABLEAU 8. \. Analyse de la variance pour le fa-cteur PHR
,,;:iource i d.l.
.
S.C. Valeur ,F PR ~ F J i Bloc 3 114.1943 3.20 0.02*
Lignée 35 6990.86 16.77 0.01**
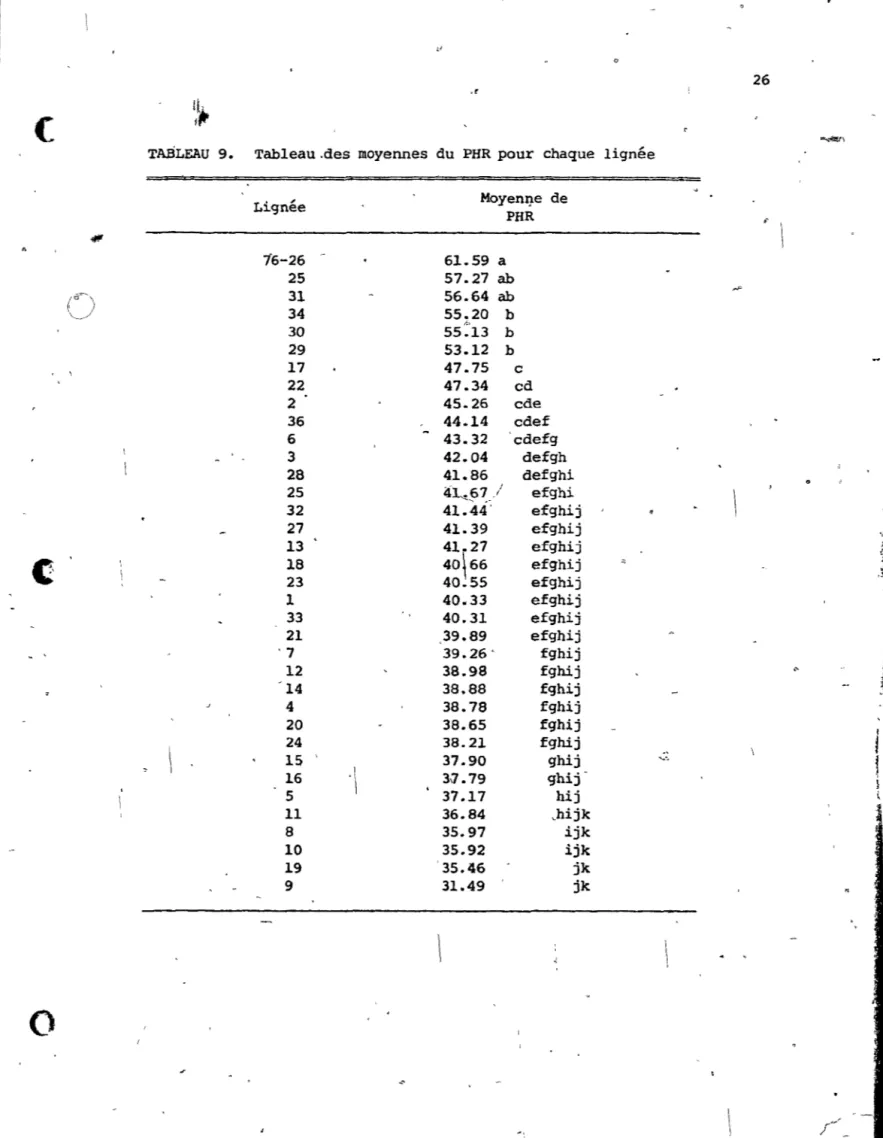
Erreur 105De plus si on anaI\Yse le Tableau 9, on peut voir que les lignées 76-26, 76-11, 76-31, 76-8 suivent les patterns observés plus haut. ,... En
, - T
-
.,
- . \ ' (1 o 26 ,r(
TABLEAU 9. Tableau.des moyennes du PHR pour chaque lignée
Lignée MoyenI].e de PHR " 16-26
-
61.59 a 25 57.27 ab()
34 31 56.64 ab 55.20 b 1.' 30 55.13 b 29 53.12 b , \ 17 47.75 c 22 47.34 cd 2 45.26 cde 36 44.14 cdef 6 43.32 cdefg 3 42.04 defgh 28 41.86 defghi 25 .lÏL.67/ ... ,.~ efghi 32 41.44' efghij 27 41.39 efghij '.~ 13 41.27 efghij,1
18 40\66 efghij 23 40.55 efghij ,~ 1 40.33 efghij " 33 40.31 efghij 21 39.89 efghij :~ '7 39.26 . fghij il' 12 38.98 fghij<-f
'14 38.88 fghij 4 38.78 fghij 20 38.65 fghiji '
24 38.21 fghij 15 37.90 ghij v,1
16'\
37.79 ghij- } 5 37.17 hij \ 11 36.84 chijk 8 35.97 ijk 10 35.92 ijk 19 '35.46 jk 9 31.49 jk~
1o
, - r
(
o
.
- ~effet la lignée 76-26 a un PHR plus élevé que la moyenne, de même que
76-31, alors que 76-11 et 76-8 ont un PHR inférieur à la moyenne.
2.1.3 Corrélationfontre les 'variables mesurées et le facteur étudié ,"
Après avoir dégagé les différences dans le rythme de développement,
on passe à l'étape sui vènte où l'on essaie d'établir une corrélation
(>5 ,
entre le mode d'évolution de la croissance et la maturité des lignées
de màis. Les résult~ts consignés au Tableau 10 montfen-t qu'il Y a des
corrélations positives et hautement signifi~at~ves entre les différentes
o
phases du développement et entre les phases du développement et le PRR.
1-.
Nous notons aussi des corrélations positives hautement significatives
entre la hauteur et le PHR, le nombre de feuilles et le PHR, l~ CF et le
nombre d' épis, la hauteur et le nombre de feuilles (Tableau Il),' Il Y a
également une corrélation né~ative hàutement significative entre le CF
et le nombre de feuilles (Tableau Il).
Il ressort du Tableau 10 une constante: toute~ les corrélations avec le PHR
.~ .~
sont positives, En effet plus les premières phases sont longues' plus le
PHR est élevé. Par ailleurs i l y a q~atre patterns différents de'
développement; donc se baser sur les corrélat~ons
entre les caractères morphologiques ne suffit pas entre toutes ces variables.
<-entr~ les Phas~s o\u ,
à eXP]iquer les relat10ns
l.I.4
Etude de la régression en plaçant le PHR comme variable dépendante Pour pouvoir étudier les relations nous avons utilisé la méthodedu R2 minimum compilée
à
l'aide du système intégré SAS 7J (Tableau 12) •Cette méthode procède par étape, en ajoutant les variables une à la fois
mais en gardant R2 minimum et en respectant la signification de 0.05
pour le , mod~le. Si on utilise cette méthode on ne retient que cinq
variables qui peuvent servir à prédire le PHR au niveau ?e ,0.05, de ~