Rôle de la méthylation de l’ADN dans la régulation de l’expression des gènes 15-LOX-1 et 15-LOX-2 dans le cartilage
Texte intégral
Figure


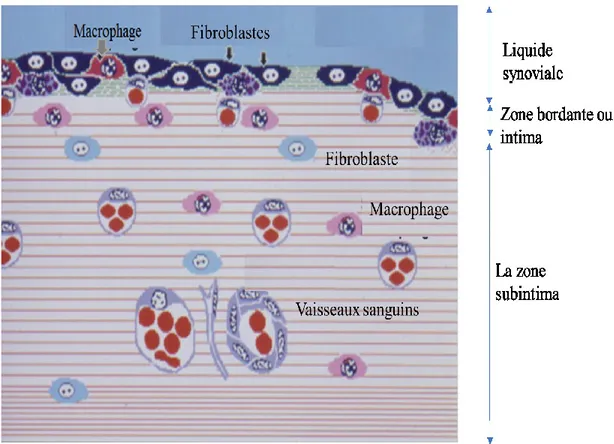

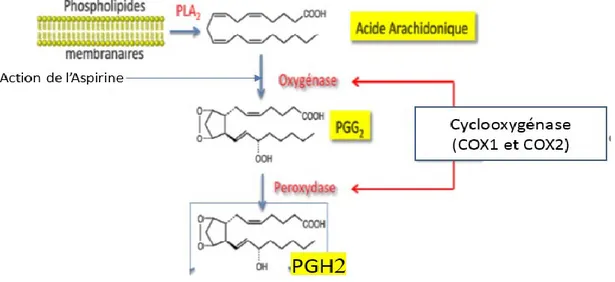

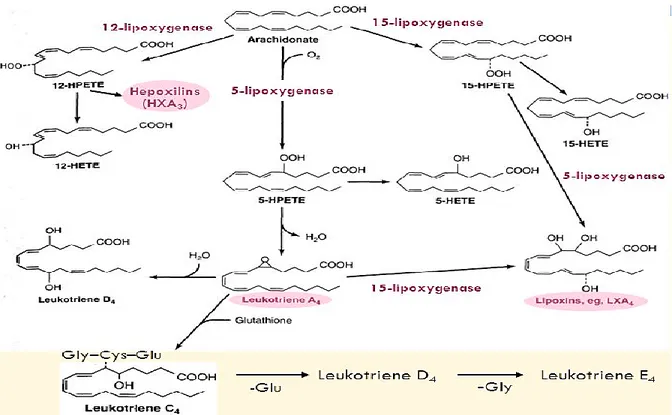
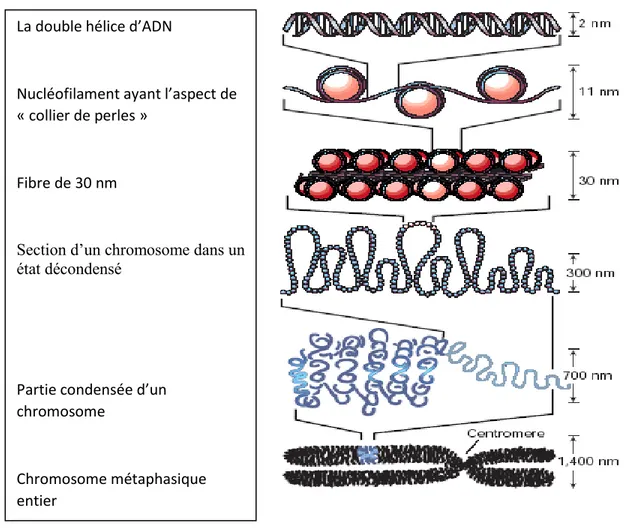
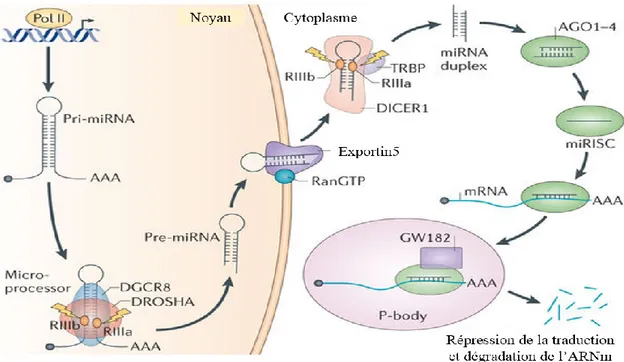
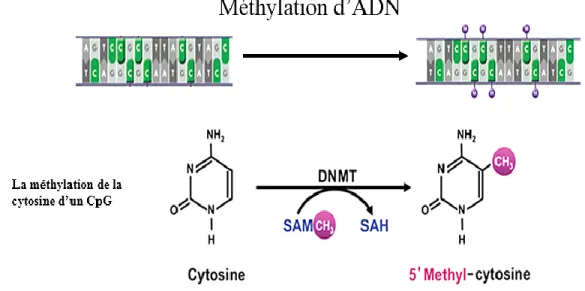
Documents relatifs
Par une méthode de criblage génique (g e n e arra y ) nous avons démontré, en premier lieu , que la répression de MTl-MMP provoque une inhibition dans l'
Bien que les résultats d’expression semblent signifier que les gènes étudiés sont seulement exprimés à des niveaux très faibles dans le placenta des
Chez le blé hexaploïde (Triticum aestivum L.) , le processus de vernalisation est régi par une des voies génétiques qui impliquent trois gènes principaux qui
La recherche des gènes impliqués dans la régulation de la prolifération et de la maturation des chondrocytes se heurte aux difficultés d'extraction d'ARN à
Sinigoj est quant à elle totalement tournée vers LMDZ et l’analyse de la variabilité interannuelle de la FTH, en suivant l’approche des régimes dynamiques appliqués aux moments de
Bien que le critère « présence/absence » d’une séquence consensus dans la région promotrice puisse fournir des indications quant aux gènes pouvant être directement régulés
Inhibition of p38 mitogen-activated protein kinase interferes with cell shape changes and gene expression associated with Schwann cell myelination.. Salzer JL,
Cependant, le recrutement de TFTC/STAGA sur les promoteurs des gènes spécifiques des photorécepteurs est aug- menté dans les souris SCA7 par rapport aux souris témoins et, en
