EFFETS DE POLYMORPHISME S GENETIQUES DE
LA VOIE DE SIGNALISATION DE LA VITAMINE D
SUR LES NIVEAUX CIRCULANTS DE VIT4AMINE D,
LE RISQUE DE CANCER DU SEIN ET LA DENSITÉ
MAMMAIRE.
Thèse présentée
à la faculté des études supérieures de l'Université Laval
dans le cadre du programme d'epidemiologic pour l'obtention
du grade de Philosophiae Doctor (Ph.D.)
Epidemiologic FACULTÉ DE MÉDECINE UNIVERSITÉ LAVAL QUÉBEC 2010 © Marc Sinotte 2010
RESUME
Objectif: Plusieurs études écologiques, d'observation et expérimentales suggèrent que des apports ou des niveaux circulants élevés de vitamine D sont associés à des risques plus faibles de divers cancers, incluant le cancer du sein. Les effets de la vitamine D passent par son récepteur (VDR) et ultimement permettent la régulation de la croissance, de la différenciation, de l'apoptose et de plusieurs autres mécanismes cellulaires impliqués dans la carcinogenèse. Les variations génétiques (telles que les SNP pour « Single Nucleotide Polymorphisms ») des gènes qui codent pour les diverses protéines impliquées tout au long de la voie de signalisation de la vitamine D sont susceptibles d'influencer les effets de la vitamine D. Dans cette thèse seront examinés trois groupes d'associations : 1) les associations entre deux SNP du VDR et le risque de cancer du sein; 2) les associations entre plusieurs SNP situées tout au long de la voie de signalisation de la vitamine D et la densité mammaire à la mammographie, celle-ci étant un important marqueur intermédiaire du risque de cancer du sein; 3) les associations entre deux SNP du gène de la protéine de transport de la vitamine D (DBP) et la concentration circulante de vitamine D, le niveau circulant de 25-hydroxyvitamine D (25(OH)D) étant le meilleur marqueur du statut global en vitamine D.
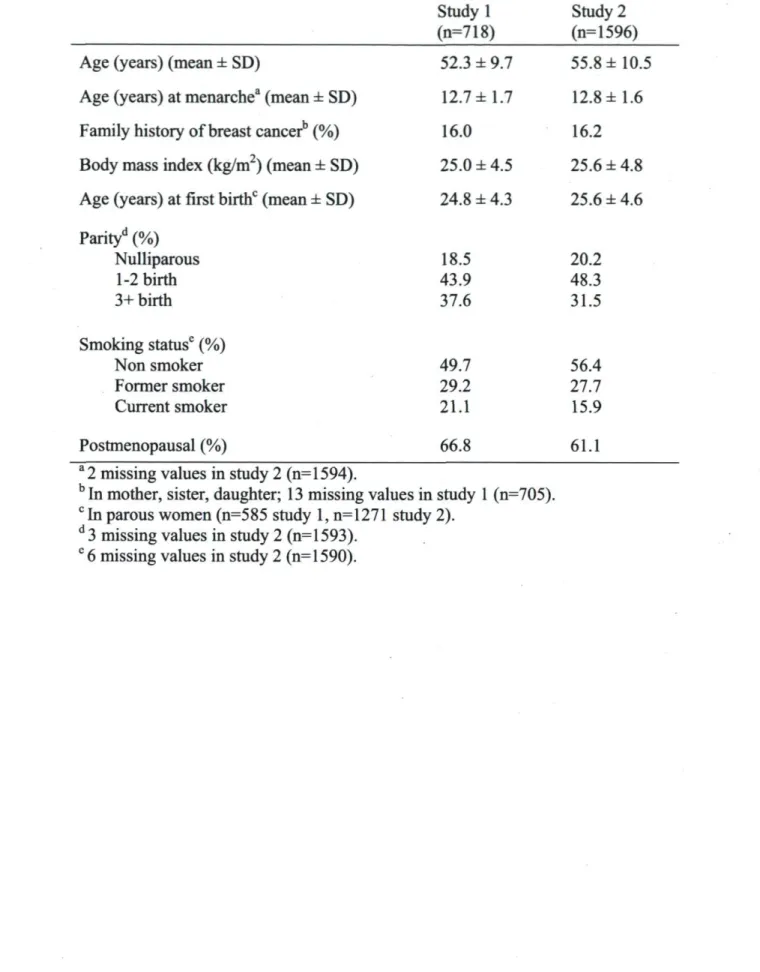
Méthodes : L'association entre deux SNP du VDR (Fokl et Bsml) et le risque de cancer du sein a été étudiée dans deux études cas-témoins indépendantes réalisées chez des femmes de la grande région de Québec (totalisant 937 cas et 1500 témoins ). Les associations entre 13 SNP de la voie de signalisation de la vitamine D et la densité mammaire ont été étudiées chez 741 femmes préménopausées ayant eu une mammographie de dépistage dans une clinique radiologique de Québec. Dans cette dernière population la relation entre deux SNP du gène de la protéine de transport de la vitamine D {DBP-1 et DBP-2) et le niveau sanguin de 25(OH)D a aussi été étudiée.
Résultats : Les génotypes ff (homozygotes rares) du polymorphisme VDR Fokl sont associés à un risque de cancer du sein plus élevé (étude 1: RC=1,22, IC95%=0,76-1,95, étude 2: RC=1,44, IC95%= 1,05-1,99, études combinées: RC=1,33, IC95%= 1,03-1,73). Cette relation est modifiée par l'histoire familiale de cancer du sein dans les deux études tout comme dans leur analyse combinée (Pinteraction^O^l, 0,050, et 0,0059 respectivement).
Les rapports de cotes parmi les femmes sans histoire familiale étaient de 1,00, 1,27 (IC95%=1,02-1,58) et 1,57 (IC95%=1,18-2,10) respectivement pour les porteuses de FF, Ff et ff (Ptendance=0,0013). Dans le cas du polymorphisme VDR Bsml, les génotypes Bb+bb étaient associés à une légère augmentation non significative du risque dans les deux études (RC combiné =1,22, IC95%=0,95-1,57) comparativement aux femmes avec le génotype BB, sans interaction avec l'histoire familiale.
L'étude de l'association entre 13 polymorphismes situés sur les gènes vitamin D receptor (VDR), cytochrome P450, family 27, subfamily B, polypeptide 1 (CYP27B1) et vitamin D-binding protein (DBP) n'a pas permis de détecter d'associations statistiquement significatives entre ces derniers et la densité mammaire (Ptendance>0,15) chez les femmes préménopausées.
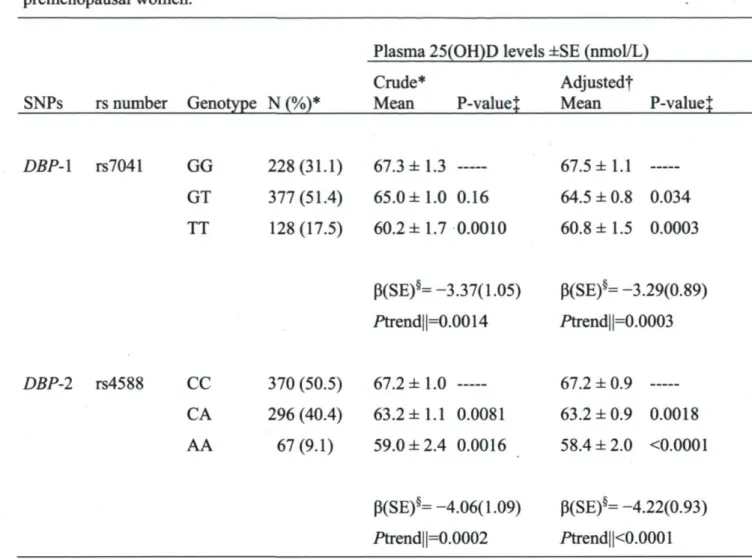
Les deux polymorphismes DBP-l et DBP-2 sont associés aux niveaux de 25(OH)D. Chaque copie additionnelle de l'allèle rare de DBP-l ou DBP-2 est associée à une baisse
des niveaux de 25(OH)D (P=-3,29, Pte„dance=0,0003; p=-4,22, Ptendance=<,0001,
respectivement). Ces associations semblent plus apparentes lorsqu'il y a beaucoup de vitamine D à transporter (P=-3,78 vs p=-l,74 pour DBP-l et B=-5,73 vs p=-3,03 pour DBP-2, respectivement en été/automne vs en hiver/printemps), l'effet modifiant n'étant
cependant pas significatif (Pinteraction=0,27 et 0,16 respectivement). Enfin, ces deux SNP expliquent autant de variation des concentrations de 25(OH)D que les apports totaux en vitamine D (iM.3% pour DBP-l et r2=2,0% pour DBP-2 vs r ^ l ^ / o pour les apports en vitamine D).
Conclusion : Dans leur ensemble les résultats présentés dans cette thèse supportent l'idée que la vitamine D et plusieurs polymorphismes de sa voie de signalisation sont susceptibles de jouer un rôle non négligeable dans l'étiologie du cancer du sein. On a en effet mis en évidence que les porteuses de certains polymorphismes du récepteur à la vitamine D sont plus à risque de cancer du sein. On a également démontré que des variations génétiques du gène de la protéine de transport de la vitamine D sont associées à un faible niveau sanguin de 25-hydroxyvitamine D (25(OH)D). Dans le contexte où les suppléments en vitamine D sont pressentis comme moyen pour réduire l'incidence du cancer du sein, ces résultats sont particulièrement importants puisqu'ils suggèrent que certains sous-groupes de la population
pourraient bénéficier différemment d'un apport de cette vitamine. Cette hypothèse devrait faire l'objet de recherches ultérieures.
ABSTRACT
Objectives : Several ecological, observational and experimental studies have shown that higher intakes or levels of vitamin D are associated with lower risk of breast cancer. Protective effects of vitamin D are though to be mediated through its receptor (VDR) and ultimately regulate growth, differentiation, apoptosis and other cell mechanisms involved in carcinogenesis. Genetic variations (as SNPs for "Single Nucleotide Polymorphisms") on genes located on the vitamin D pathway can possibly mediate the effects of vitamin D. In this thesis we will study three groups of associations : 1) associations between two VDR SNPs and breast cancer; 2) associations between several SNPs located along the vitamin D signalling pathway and mammographie breast density, which is an important intermediate marker of breast cancer risk; 3) associations between two SNPs on the vitamin D binding protein gene (DBP) and circulating levels of 25-hydroxyvitamin D (25(OH)D), the best
marker of human vitamin D status.
Methods : Several polymorphisms (SNPs) located on the vitamin D pathway were genotyped in three different samples of women from Québec city area. The association between two VDR SNPs (Fokl and Bsml) and breast cancer was assessed in two independent case-control studies from Québec City area (for a total 937 cases et 1500 controls). Associations between 13 SNPs on genes of the vitamin D pathway and breast density were investigated in 741 premenopausal women from a Québec breast screening clinic. In the same population, the relation between two SNPs on the vitamin D binding protein gene (DBP) and 25(OH)D was also investigated.
Results : Compared to common homozygotes, VDR Fokl ff carriers (rare homozygotes) were associated to higher breast cancer risk (study 1: OR=1.22, 95%CI=0.76-1.95, study 2: OR=1.44, 95%CI=1.05-1.99 and combined studies: OR=1.33, 95%CI= 1.03-1.73). This association was significantly modified by family history of breast cancer in the two studies as well as in the combined analysis (Pintcraction=0.031, 0.050, and 0.0059 respectively). Among women without family history, odds ratios were 1.00, 1.27 (95%CI= 1.02-1.58) and
1.57 (95%CI=1.18-2.10) respectively for FF, Ff and ff carriers (JW=0.00 13). In the case of Bsml, Bb+bb genotypes were associated with a weak non significant increased risk in the
two studies (combined OR=1.22, 95%CI=0.95-1.57) compared to women with the BB genotype, without interaction with family history.
No statistically significant associations have been detected (Pmn£0*15) between 13 polymorphisms located on vitamin D receptor (VDR), cytochrome P450, family 27, subfamily B, polypeptide 1 (CYP27B1), vitamin D binding protein (DBP) genes and breast density in premenopausal women.
Both polymorphisms DBP-l and DBP-2 were associated with 25(OH)D. Each additional copy of DBP-l or DBP-2 rare allele is associated with lower 25(OH)D levels (p=-3.29, 4Ptrend=0.0003; P=-4.22, 4Ptrend<0.0001, respectively). These associations seemed more apparent when more vitamin D needed to be transported (P=-3.78 vs B=—1.74 for DBP-l and P=-5.73 vs P=-3.03 for DBP-2, respectively for summer/autumn vs winter/spring) although this effect modification was not statistically significant i/Pinteraction=0.27 and 0.16 respectively). Moreover these DBP polymorphisms explained as much of the variation in circulating 25(OH)D as did total vitamin D intake (f==1.3% for DBP-l and r2=2.0% for DBP-2 vs r2<1.2% for vitamin D intake).
Conclusion : The findings reported in this thesis support the idea that vitamin D and several polymorphisms of its signalling pathway are probably playing a significant role in the etiology of breast cancer. Indeed we showed that some VDR SNPs carriers are at higher risk of breast cancer. We also demonstrated that two SNPs of the vitamin D binding protein gene (DBP) are associated to low levels of circulating levels of 25-hydroxyvitamin D (25(OH)D). In the context where vitamin D supplementation has been recently recommended as a way to reduce breast cancer incidence, these results suggest that some groups of the population might differently beneficiate from such an intervention. This hypothesis needs further investigation.
AVANT-PROPOS
Durant mes études j'ai eu la chance de participer à plusieurs projets et je fus impliqué à toutes les étapes de la recherche incluant le développement de questions de recherche, l'élaboration d'un protocole d'étude et d'une demande de subvention, la réalisation d'une collecte et de plusieurs analyses de données. J'ai pu aussi rédiger et participer à la rédaction d'articles scientifiques. Dans l'ensemble, ces activités m'ont permis d'acquérir une formation complète pour la réalisation d'études épidémiologiques. Cette thèse présente les résultats d'un sous ensemble de ces travaux.
Durant mes études de doctorat j'ai partagé mon temps entre deux thèmes de recherche favorisant l'acquisition d'habilités distinctes. Premièrement, je me suis impliqué dans le développement d'un projet de recherche et la préparation d'une demande de subvention intitulée: Hormonally-Active Environmental Contaminants and Breast Cancer Cell Proliferation. Plusieurs contaminants environnementaux ont des activités hormonomimétiques reconnues et certains ont été individuellement associés au risque ou à l'agressivité du cancer du sein. Ces contaminants constituent possiblement des facteurs de risque environnementaux modifiables. Toutefois seule une infime minorité de ces contaminants a été étudiée individuellement, et l'effet conjoint de l'exposition à plusieurs contaminants mérite une attention particulière puisqu'elle reflète davantage la réalité. Le projet de recherche portait d'une part, sur la mesure de 145 contaminants bioaccumulables grâce à une nouvelle méthode analytique chez des femmes ménopausées de la région de Québec, et d'autre part sur l'évaluation de l'activité hormonale individuelle ou conjointe de ces contaminants dans des cultures cellulaires de cellules mammaires. Une analyse du lien entre les niveaux de ces contaminants et la densité mammaire, un important marqueur de risque du cancer du sein, était aussi prévue. Cette demande de subvention ayant été accordée, j'ai été amené à contribuer à plusieurs étapes de la réalisation du projet par exemple: le développement du questionnaire, la rédaction des formulaires de consentement, la planification et la supervision du recrutement des participantes et des témoins, le développement des outils de suivi des échantillons, de même que la planification et la réalisation du contrôle de qualité pour les analyses de nouveaux marqueurs biologiques. J'ai, pour l'instant, contribué à analyser une partie des données, rédigé des résumés et
publié un article scientifique dans le cadre de ce projet (référence 5 ci-bas). Cet article tend à démontrer que les femmes de la région de Québec ont des concentrations plasmatiques de polybromodiphényléther (PBDE), dont on se sert comme ignifuge en Amérique du nord, beaucoup plus élevées qu'en Europe où ces substances ont été réglementées ou bannies. On y démontre également que les expositions à ces substances bioaccumulables sont vraisemblablement plus récentes et moins liées à la chaîne alimentaire que les BPC. D'autres analyses se poursuivent, par exemple pour évaluer la relation entre le niveau sérique de 12 hormones sexuelles (dont l'estradiol, l'estrone, la testosterone, la déhydroépiandrostérone) et la densité mammaire telle que visualisée à la mammographie. Deuxièmement, j'ai fait des recherches sur le lien entre la vitamine D et le cancer du sein, un sujet d'importance grandissante. Après avoir mis en évidence que certains polymorphismes génétiques du récepteur à la vitamine D étaient associés au risque de cancer du sein dans une banque de données sur les contaminants organochlorés, j'ai voulu tenter de répliquer ces résultats dans une autre étude indépendante. Ce sont précisément ces replications, souvent absentes, qui donnent davantage de crédibilité aux résultats d'association génétique que l'on rapporte dans le premier article de la thèse (référence 1 ci-dessous). Suivant l'obtention de ces résultats positifs sur deux polymorphismes du récepteur à la vitamine D (VDR), la suite logique fut de rechercher les effets de ceux-ci dans une banque de données sur la densité mammaire à la mammographie. Dans cette banque j'ai également étudié d'autres polymorphismes de la voie de signalisation de la vitamine D (référence 2 ci-bas). Enfin, c'est grâce à cette dernière banque de données que j'ai pu mettre en évidence pour la première fois que deux polymorphismes du récepteur à la vitamine D (DBP) contribuent à expliquer les variations dans les concentrations circulantes de vitamine D, et ce de manière équivalente ou supérieure aux apports totaux (i.e. nutrition et suppléments) de cette dernière vitamine (référence 3 ci-dessous).
On doit également ajouter une compétence supplémentaire touchant le développement du sens critique par l'analyse et la présentation hebdomadaire de divers articles scientifiques dans le cadre du club de lecture de notre équipe. Cette activité hautement formatrice et toujours stimulante touchait de près ou de loin nos sujets d'intérêt immédiat.
Puisque les données utilisées pour la réalisation des articles sur les polymorphismes génétiques sont issus de projets de recherche initiés préalablement à la réalisation de cette
thèse, il va sans dire que les résultats originaux qu'on peut en tirer découlent de la vision et de l'intuition des chercheurs à l'origine de ces projets, sans qui il aurait été impossible de maximiser l'utilisation des données récoltées il y a plusieurs années. Puisque les questions de recherche à la base des articles de cette thèse découlaient souvent des échanges et discussions entre les membres de l'équipe, cela renforce chez moi la conviction qu'il faut favoriser les interactions entre les individus de manière structurée pour stimuler les réflexions tout en permettant la créativité. Cette dernière constatation est certainement un acquis initialement insoupçonné, mais combien important, associé à la réalisation de ce projet.
Ci-dessous vous trouverez la liste des publications que j'ai produites ou auxquelles j'ai participé durant le déroulement de mes études de doctorat. Elle regroupe mes contributions au vaste champ de l'épidémiologie.
1. Sinotte, M., Rousseau, F., Ayotte, P., Dewailly, E., Diorio, C , Giguère, Y., Bérubé, S., and Brisson, J. Vitamin D receptor polymorphisms (Fokl, Bsml) and breast cancer risk: association replication in two case-control studies within French Canadian population. Endocr Relat Cancer, 2008.
2. Sinotte, M., Diorio, C , Brisson, J., Bérubé, S., and Pollak, M. Vitamin D pathway polymorphisms in relation to mammographie breast density. Cancer Epidemiol Biomarkers Prev, 17: 2505-8, 2008.
3. Sinotte, M., Diorio, C , Bérubé, S, Brisson, J., et Pollak, M. 2008. Genetic polymorphisms of the vitamin D binding protein and plasma levels of 25-hydroxyvitamin D in premenopausal women. Am J Clin Nutr, 2008.
4. Brisson, J., Bérubé, S., Diorio, C, Sinotte, M., Pollak, M. et Masse, B. 2007. Synchronized seasonal variations of mammographie breast density and plasma 25-hydroxyvitamin d. Cancer Epidemiol Biomarkers Prev 16 929-933.
5. Sandanger, TM., Sinotte, M., Dumas, P., Marchand, M., Sandau, CD., Pereg, D., Bérubé, S., Brisson, J., et Ayotte, P. 2007. Plasma concentrations of selected organobromine compounds and polychlorinated biphenyls in postmenopausal women of Quebec, Canada. Environ Health Perspect 115 1429-1434.
REMERCIEMENTS
Un projet de doctorat comme celui-ci a nécessité pour sa réalisation la collaboration de plusieurs personnes, mais en premier lieu il faut remercier toutes les femmes qui ont accepté de participer aux divers projets de recherche ayant généré les données utilisées dans cette thèse. Sans elles il va sans dire qu'aucune recherche épidémiologique n'aurait été possible.
Ensuite ma gratitude doit aller à mon directeur Jacques Brisson qui m'a soutenu tout au long de ce processus. Nos échanges, entre autres durant les rencontres du club de lecture, ont toujours été chaleureux et stimulants. Je dois également souligner le support continu de ma collègue Sylvie Bérubé qui, avec son expérience pratique et sa disponibilité, a su m'aider à toutes les étapes de la réalisation d'un projet de recherche et favoriser le développement des habiletés qui s'y rattachent. Ma reconnaissance va aussi à Caroline Diorio, ma collègue de bureau, pour nos échanges toujours animés d'un esprit de partage et d'entraide, et ce dans tous les domaines, qu'ils touchent ou non la réalisation de cette thèse. Je dois également remercier mon co-directeur Pierre Ayotte avec lequel j'ai eu tellement de discussions intéressantes lors de la préparation de la demande se subvention du projet de recherche touchant les substances hormono-actives, et au sujet des impacts que pourraient avoir les polluants environnementaux sur la santé. Mes remerciements vont aussi à Madame Danielle Audet, infirmière de recherche, avec laquelle j'ai collaboré lors du recrutement des participantes et dans bien des étapes de ce projet de recherche touchant les substances hormono-actives, pour sa gaieté et son dynamisme contagieux.
Ma reconnaissance va également à tous les membres de l'équipe de statisticiens, pour leur disponibilité infaillible à un moment où l'autre de ce projet. Il faut de plus souligner la bonne humeur généralisée de tous les gens que j'ai eu la chance de côtoyer à l'unité de recherche en santé des populations et qui ont fait de mon passage à l'URESP un séjour toujours agréable.
Enfin je ne pourrais passer sous silence les encouragements soutenus de ma conjointe Linda, et surtout de sa patience particulièrement en fin de parcours alors que nos deux enfants, Maé et Romy, me demandaient quotidiennement:" Est-il fini ton doctorat papa?"
Table des matières
RÉSUMÉ ii ABSTRACT v AVANT-PROPOS vii
REMERCIEMENTS x Table des matières xiii LISTE DES TABLEAUX xv LISTE DES FIGURES xvi CHAPITRE I : Introduction 17
PRÉAMBULE 18 ÉTAT DES CONNAISSANCES 20
La vitamine D, son métabolisme et sa voie de signalisation 20 Cancer du sein, vitamine D et polymorphismes du VDR 29 Densité mammaire, vitamine D et polymorphismes de sa voie de signalisation 32
25-hydroxyvitamine D, protéine de transport et polymorphismes de DBP 35 FORCES ET LIMITES DES ÉTUDES D'ASSOCIATION GÉNÉTIQUE 37
PRÉSENTATION DE LA RECHERCHE 42
Objectifs et stratégies 42 Populations et collectes de données 43
Analyses statistiques des données 48
Considérations éthiques 49 CHAPITRE II : Vitamin D Receptor Polymorphisms (Fokl, Bsml) and Breast Cancer Risk:
association replication in two case-control studies within French Canadian population 50
RÉSUMÉ 51 ABSTRACT 52 INTRODUCTION 53 MATERIAL AND METHODS 54
Study population 1 54 Study population 2 55 Statistical methods 56
RESULTS 57 Associations of Fokl and Bsml with invasive breast cancer 57
Effect modification by family history of breast cancer 58
DISCUSSION 58 Fokl and breast cancer 58
Bsml and breast cancer 60
Limitations 60 Conclusion 61 AKNOWLEDGEMENTS 61
REFERENCES.... 66 CHAPITRE III : Genetic Polymorphisms Involved In Vitamin D Pathway In Relation To
Mammographie Breast Density 71
RÉSUME 72 ABSTRACT 73
INTRODUCTION 75 MATERIAL AND METHODS 75
RESULTS 76 DISCUSSION 77 AKNOWLEDGEMENTS 78
REFERENCES 82 CHAPITRE IV : Association Between Genetic Polymorphisms Of The Vitamin D Binding
Protein And Plasma Concentrations Of 25-Hydroxyvitamin D In Premenopausal Women 85
RÉSUMÉ 86 ABSTRACT 87 INTRODUCTION 88 MATERIAL AND METHODS 89
Study population and recruitment procedures 89
Data collection 89 Assessment of plasma 25(OH)D 89
DNA extraction and SNP genotyping 90
Statistical methods 90
RESULTS 91 Determinants of 25(OH)D levels 91
Vitamin D binding protein SNPs and 25(OH)D levels 92 Vitamin D load and strength of DPB SNP effects 92 Contribution to variation of25(OH)D levels 92
DISCUSSION 93 ACKNOWLEDGEMENTS 96
REFERENCES 101 CHAPITRE V : Conclusion 105
LISTE DES TABLEAUX
Tableau 1 : SNP étudiés dans cette thèse 26 Tableau 2 : Characteristics of women by study 63 Tableau 3 : Fokl and Bsml VDR polymorphisms and breast cancer risk 64
Tableau 4 : Fokl VDR polymorphism and breast cancer risk by family history of breast
cancer 65 Tableau 5 : Single nucleotide polymorphisms evaluated in the present study 79
Tableau 6 : Associations of single nucleotide polymorphisms in vitamin D pathway genes
with mammographie breast density 80 Tableau 7 : Relation between plasma 25(OH)D levels and potentially associated variables
in premenopausal women 97 Tableau 8 : Relation between plasma 25(OH)D levels and vitamin D binding protein
genotypes in premenopausal women 98 Tableau 9 : Relation between plasma 25(OH)D levels and genotypes of vitamin D binding
protein by vitamin D load in premenopausal women 99 Tableau 10 : Relative contribution of DBP-l or DBP-2 SNPs and other potential
explanatory variables to explanation of plasma 25(OH)D levels, globally and stratified
LISTE DES FIGURES
Figure 1 : Modèle conceptuel situant les sources, le transport et le métabolisme de la
vitamine D en relation avec la densité mammaire et le cancer du sein 23 Figure 2 : Exemples de faible (<10%) et de forte (>70%) densité mammaire 32 Figure 3 : Association entre les densités mammaires et le risque relatif de cancer du sein
PRÉAMBULE
Les effets de la vitamine D ont premièrement été remarqués sur la santé osseuse mais ses actions vont bien au delà et sont de plus en plus reconnues. Plusieurs études suggèrent que les apports ou les niveaux circulants de vitamine D sont des facteurs de risque modifiables de plusieurs maladies chroniques, incluant des maladies auto-immunes, infectieuses, cardiovasculaires et de différents cancers, parmi lesquels on cite fréquemment les cancers du colon, de l'ovaire, du pancréas, du poumon, de la prostate et du sein, (revu dans (1)). Une récente méta-analyse de 18 essais cliniques randomisés s'intéressant initialement à l'effet de la vitamine D sur la santé osseuse, avec une dose quotidienne moyenne de 528 UI de vitamine D sur une durée moyenne de 5,7 ans, révèle une baisse significative (RR=0,93, IC95%=0,87-0,99) de la mortalité totale chez les patients ayant reçu la vitamine D par rapport aux non exposés (2).
Le cancer du sein, qui touchera une forte proportion de femmes à un moment ou l'autre de leur vie, est une maladie dont l'incidence est à la hausse. Cette maladie fait l'objet d'intenses recherches quant à son étiologie. La vitamine D pourrait avoir une influence sur le risque de ce cancer. De nombreuses études écologiques et d'observation rapportent une hausse de l'incidence et de la mortalité de ce cancer associée au fait de vivre à des latitudes plus élevées où l'exposition au soleil est plus faible, d'avoir des apports nutritionnels réduits de vitamine D, ou des niveaux circulants plus faibles de cette vitamine (revu dans (1, 3, 4). Un essai clinique randomisé d'une durée de quatre ans, dans lequel les participantes consommaient des suppléments alimentaires contenant 1100 UI de vitamine D, apport suffisant pour rehausser le niveau moyen de 25(OH)D de 24 nmol/L, a récemment rapporté une baisse du risque général de cancer (RR=0,23, IC95%=0,09-0,60) incluant le risque de cancer du sein qui était le cancer le plus fréquent (i.e. 41%, 17/41) (5). De plus, des études épidémiologiques récentes suggèrent que les femmes pré-ménopausées ayant des apports alimentaires ou des concentrations circulantes élevés en vitamine D ont une plus faible densité mammaire à la mammographie (6, 7); la densité mammaire constitue un important facteur de risque du cancer du sein.
Les objectifs de cette thèse portent sur les voies par lesquelles la vitamine D pourrait affecter la survenue du cancer du sein. Pour être active la vitamine D, synthétisée dans la peau ou ingérée, doit être transportée par sa protéine de transport jusqu'au foie pour y être hydroxylée une première fois. Par la suite, et de la même manière, elle est transportée jusqu'aux reins et à d'autres organes pour y être hydroxylée une seconde fois avant de
pouvoir finalement se lier à son récepteur et engendrer ses effets biologiques. Tout au long de cette voie métabolique, diverses protéines doivent entrer en jeu, incluant les protéines qui transportent la vitamine D, les enzymes d'hydroxylation et le récepteur de la vitamine D. Il est plausible que des variations dans le code génétique de l'un ou l'autre des gènes impliqués dans la production desdites protéines modulent l'efficacité de cette voie métabolique. Si tel est le cas, à exposition égale à la vitamine D, des variations au niveau des gènes codant la protéine de transport, les enzymes d'hydroxylation, ou encore le récepteur à la vitamine D, pourraient expliquer des différences au niveau de la concentration de vitamine D circulante, du risque de cancer du sein, ou encore du risque d'avoir une forte densité mammaire. Ce sont là les hypothèses que nous avons explorées dans cette thèse. Plus précisément, nous avons étudié trois relations : premièrement, celle entre deux variations génétiques du récepteur à la vitamine D et le risque de cancer du sein; deuxièmement, la relation entre 13 variations génétiques situées sur trois gènes impliqués à diverses étapes du métabolisme de la vitamine D et le pourcentage de densité mammaire; et troisièmement, la relation entre deux variations génétiques de la protéine de transport de la vitamine D et la concentration circulante de cette vitamine.
Le reste de ce chapitre compte trois sections, soit une qui résume l'état des connaissances, une qui discute des forces et faiblesses des études d'association génétique et une dernière qui expose la méthodologie des études réalisées.
ÉTAT DES CONNAISSANCES
La vitamine D, son métabolisme et sa voie de signalisation
Sources de vitamine D et effets de la vitamine D sur la santé
La vitamine D, une vitamine liposoluble, fait partie de la famille des sécostéroïdes et possède une structure similaire aux stéroïdes. Elle est considérée comme une prohormone ayant deux formes principales, la vitamine D3 (ou cholécalciférol) et la vitamine D2 (ou ergocalciférol). La vitamine D3, d'origine animale, est synthétisée par la peau sous l'effet des rayonnements UVB, tandis que la vitamine D2 est d'origine végétale. L'exposition au soleil serait la source principale de vitamine D chez l'humain puisque peu d'aliments en contiennent naturellement des quantités appréciables. En effet, les poissons gras et les huiles de poissons sont pratiquement les seules sources alimentaires significatives de cette vitamine. C'est ce qui explique que plusieurs pays supplémentent divers aliments en vitamine D, comme par exemple le lait au Canada. Des suppléments alimentaires contenant habituellement 400, 800, 1000 et 2000 UI sont également disponibles en pharmacie (revu dans (1)).
Les principales causes de carence en vitamine D seraient le manque d'exposition au soleil, le fait de vivre sous des latitudes nordiques, d'avoir une peau foncée et l'ingestion insuffisante des aliments qui en contiennent. L'obésité, qui engendrerait la séquestration des vitamines liposolubles dans les tissus adipeux, et le vieillissement, qui réduirait la capacité de synthèse de cette vitamine au niveau de la peau, sont aussi des facteurs associés au manque de vitamine D. De plus, certaines maladies entraînant une mauvaise absorption des lipides dans l'intestin sont associées à l'hypovitaminose D (revu dans (8)).
La déficience en vitamine D est généralement définie par un niveau circulant de 25(OH)D sous 50 nmol/L (20 ng/ml), tandis que l'insuffisance se situerait entre 50 et 75 nmol/L (20-30 ng/ml). Les concentrations optimales seraient entre 75 et 150 nmol/L ((20-30-60 ng/ml), alors que les intoxications nécessiteraient des concentrations de 375 nmol/L (150 ng/ml) et plus (revu dans (1)). Les symptômes d'intoxication, tels l'hypercalcémie et l'hyperphosphatémie, sont rares et généralement dus à l'ingestion de doses quotidiennes extrêmement élevées (e.g. 50 000 UI) (revu dans (1)). La prise quotidienne de suppléments de 10 000 UI pendant 5 mois n'aurait pas causé d'intoxication. À noter qu'une intoxication suite à une exposition excessive au soleil est impossible puisque au-delà d'un certain seuil, la vitamine D3 est dégradée au site de production (revu dans (1)).
Dans les sociétés développées, des facteurs de risque importants de l'hypovitaminose D comme l'obésité, le vieillissement de la population et l'exposition réduite au soleil, vont croissants. À titre d'exemple, au Québec, les concentrations moyennes circulantes de 25(OH)D de notre échantillon des femmes préménopausées sont approximativement de 56, 61, 78 et de 62 nmol/L de 25(OH)D pour l'hiver, le printemps, l'été et l'automne, respectivement. Au regard des seuils énoncés plus haut, on constate que même parmi ces femmes relativement jeunes, les concentrations optimales de 25(OH)D seraient tout juste atteintes pendant l'été et qu'elles demeureraient insuffisantes le reste de l'année (9).
Les effets de la vitamine D ont premièrement été remarqués pour la santé osseuse mais ses actions à bien d'autres niveaux sont de plus en plus reconnues. En effet des apports ou concentrations circulantes plus élevés de vitamine D ont été associés à une réduction du risque de plusieurs maladies chroniques incluant des maladies auto-immunes, infectieuses, cardiovasculaires et plusieurs cancers (revu dans (1, 3)). Parmi ces derniers on cite fréquemment les cancers du colon, de l'ovaire, du pancréas, du poumon, de la prostate et du sein. De plus, une récente méta-analyse de 18 essais cliniques randomisés avec une dose quotidienne moyenne de 528 UI de vitamine D sur une durée moyenne de 5,7 ans, révèle en effet une baisse significative (RR=0,93, IC95%=0,87-0,99) de la mortalité totale (2). De même, un essais clinique randomisé d'une durée de quatre ans où les participantes consommaient des suppléments alimentaires contenant 1100 UI de vitamine D, suffisants pour rehausser le niveau de 25(OH)D au dessus de 80 nmol/L, a récemment rapporté une baisse du risque général de cancer (RR=0,23, IC95%=0,09-0,60) parmi lesquels le cancer du sein était le plus fréquent (i.e. 41%, 17/41) (5). Par ailleurs, des études expérimentales in vivo et in vitro démontrent que la vitamine D inhibe la prolifération cellulaire, induit l'apoptose (i.e. mort cellulaire programmée) et arrête la croissance tumorale, tous des processus impliqués dans la carcinogenèse (revu dans (10, 11)).
La convergence des résultats scientifiques sur les effets de la vitamine D sur la santé est suffisamment grande pour que plusieurs experts internationaux suggèrent de prendre action pour rehausser le niveau de vitamine D dans la population, estimant que cela se traduira par une réduction significative du taux de mortalité générale (2) ou par cancer (12). De même, les données scientifiques brièvement décrite ci-dessus laissent penser que l'administration de vitamine D ou de ses analogues pourrait constituer une mesure de prévention de plusieurs cancers. D'ailleurs, depuis 2007, la Société Canadienne du Cancer recommande son utilisation pour la prévention primaire du cancer du sein :
http://www.cancer.ca/ccs/internet/mediareleaselist/0„3172 1613121606 1997621989 la ngld-en.html.
Voie métabolique de la vitamine D
Pour que les effets de la vitamine D surviennent, plusieurs réactions situées tout au long de sa voie métabolique doivent se produire. Cette voie métabolique consiste en une suite de réactions couplées, où le produit d'une réaction devient le substrat d'une réaction subséquente et ainsi de suite. Tel que schématisé à la figure 1 on constate que pour engendrer leurs effets, les prohormones vitamine D3 et vitamine D2 insolubles doivent d'abord être transportées au foie par leur protéine de transport DBP (pour Vitamin D Binding Protein). Elles y subissent une première hydroxylation en 25-hydroxyvitamine D (25(OH)D) grâce à l'enzyme CYP27A1 (pour Cytochrome P450, famille 27, sous- famille A, polypeptide 1) puis reprennent la circulation périphérique pour subir une seconde hydroxylation en 1,25-dihydroxyvitamine D (l,25(OH)2D). Cette dernière hydroxylation se produit principalement dans les reins grâce à l'enzyme CYP27B1 (pour P450, famille 27, sous- famille B, polypeptide 1), mais il y a aussi production locale de l,25(OH)2D avec la même enzyme dans d'autres organes cibles comme le sein notamment (revu dans (1)). C'est alors que la l,25(OH)2D pourra se lier au récepteur VDR (pour Vitamin D Receptor) de la vitamine D afin d'engendrer, entre autres, la production des protéines déclenchant la cascades des effets biologiques de la vitamine D.
Peau et
système digestif
UVB Nourriture et suppléments Vitamine DCirculation
sanguine
Foie
Circulation
sanguine
Vitamine D + DBP*Vitamine D + CYP27A1 = 25(OH)D
25(OH)D + DBP<
Reins
Circulation
sanguine
25(0H)D + CYP27B1* l,25(OH)2D l,25(OH)2D + DBPSein
25(0H)D + CYP27B1* l,25(OH)2DZ
l,25(OH)2D + VDR* Effets physiologiquesDensité mammaire ? Cancer du sein?
Figure 1 : Modèle conceptuel situant les sources, le transport et le métabolisme de la vitamine D en relation avec la densité mammaire et le cancer du sein.
Les polymorphismes situés sur certains gènes de la voie de signalisation de la vitamine D étudiés dans cette thèse sont marqués d'un astérisque. UVB = rayonnement ultraviolet (315-280 nm). DBP = protéine de transport de la vitamine D. CYP27A1 = enzyme servant à la conversion de la vitamine D en 25(OH)D). CYP27B1 = enzyme servant à la conversion de la 25(OH)D) en (l,25(OH)2D). VDR = récepteur de la vitamine D.
Polymorphismes de la voie de signalisation de la vitamine D
Comme schématisés à la figure 1, les effets ultimes de la vitamine D dépendent de plusieurs molécules telles la protéine de transport (DBP), les enzymes d'hydroxylation et le récepteur de la vitamine D. Les gènes contenant l'information nécessaire à la synthèse de ces molécules composent ce qui est appelé 'la voie de signalisation de la vitamine D'.
Un gène est une portion d'un brin d'ADN; c'est donc une petite séquence de nucleotides, eux-mêmes composés d'un groupement phosphate, d'un désoxyribose (sucre à 5 carbones) et d'une base azotée variable selon le nucleotide. Dans le processus de synthèse d'une protéine, cette portion d'ADN est copiée lors de la transcription en ARN messager. L'ARN messager, qui passe du noyau de la cellule à son cytoplasme, est ensuite traduit en une chaîne d'acides aminés (et en protéines) par l'intermédiaire de l'ARN de transfert. Une suite de trois bases azotées consécutives sur TARN messager forme un codon qui code le recrutement d'un acide aminé donné (revu dans (13)).
On peut concevoir que si la séquence des bases azotées d'un gène est invariable d'un individu à l'autre, toutes les protéines produites selon ce gène seront identiques ou presque. À l'inverse si la séquence varie d'une personne à l'autre, les acides aminés de la protéine tributaire de ce gène pourraient aussi varier d'un individu à un autre et, par conséquent, l'efficacité de la protéine en question pourrait présenter des variations inter-individuelles. Lorsque dans une population il y a des variations entre les individus dans la séquence des bases azotées de l'ADN d'un gène donné, on parle de polymorphismes ou de variations génétiques. On dira que ce gène a plusieurs alleles. Ces variations génétiques (ou ces différents alleles), proviennent de mutations antérieures et des adaptations qui ont pu en découler. Les variations génétiques qui surviennent sur une seule paire de base d'un gène représentent 90% de l'ensemble des variations génétiques humaines. On les appellent
'Single Nucleotide Polymorphisms' (SNP). Les SNP peuvent se retrouver au sein de régions codantes des gènes (i.e. exons) qui seront traduites en séquence d'acides aminés, ou dans des régions non codantes (i.e. introns) qui ne le seront pas. Ils peuvent aussi se retrouver dans deux petites parties flanquant les extrémités du gène qui, bien que non codantes, sont également importantes pour l'initiation (e.g. région promotrice de l'extrémité 5'UTR) et le contrôle (i.e. 3'UTR) de la transcription du gène (revu dans (13,
14)). Ces SNP sont donc susceptibles d'affecter directement ou indirectement la production des protéines.
Ainsi, lorsque des variations génétiques prévalent, un gène donné peut avoir diverses formes appelées alleles, et puisque chaque individu est porteur de deux copies d'un gène, on retrouvera dans la population des groupes d'individus qui seront hétérozygotes, ou encore homozygotes pour l'allèle majeur (i.e. le plus commun) ou l'allèle mineur (i.e. le plus rare). Par exemple, le SNP Fokl du VDR étudié aux chapitres II et III de cette thèse et présenté au tableau 1, contient deux alleles, appelés G ou A qui correspondent aux bases azotées guanine ou adenine respectivement. Dans notre population de témoins (tout comme dans les autres populations d'origine caucasienne), on retrouve environ 37 % d'homozygotes pour l'allèle majeur (GG), 16 % d'homozygotes pour l'allèle mineur (AA), et 47 % d'hétérozygotes (GA). Il est bon de noter que les alleles peuvent alternativement être nommés en référence aux enzymes ayant servi à leur détection. Dans notre exemple les alleles G et A sont synonymes de F et f tout comme les génotypes GG, GA et AA sont synonymes de FF, Ff et de ff respectivement. C'est l'usage courant dans la littérature qui a guidé le choix de la nomenclature utilisée dans cette thèse. Les études d'association génétique tentent de mettre en évidence les relations entre différents alleles ou génotypes et le risque de maladies dans les populations où des polymorphismes existent.
Dans la présente thèse nous avons étudié des polymorphismes du gène codant la synthèse du récepteur à la vitamine D, du gène codant la synthèse d'une enzyme impliquée dans la transformation de la vitamine D, et du gène codant la synthèse de la protéine de transport de la vitamine D. Les astérisques à la figure 1 indiquent où se situent ces polymorphismes sur la voie de signalisation de la vitamine D. Chacun de ces polymorphismes est également présenté au tableau 1 et est décrit plus en détails ci-dessous selon l'ordre d'apparition dans la thèse. Par convention les acronymes utilisés lorsque l'on fait référence aux gènes ou à leurs polymorphismes, plutôt qu'aux protéines qu'ils codent, doivent être écrits en italique.
Tableau 1 : SNP étudiés dans cette thèse.
Alleles
Nom (majeur> Numéro de Position sur le gène
du gène Séquence codant pour : mineur) référence :* (autre appellation) : VDR Récepteur de la vitamine D G > A
O G
T>C
G>A
G>A
C>T
A>C
T>C
CYP27B1 Cytochrome P450, famille 27, A > G sous- famille B, polypeptide 1 C > A
O G DBP Protéine de transport de G > T la vitamine D C > A rsl 1568820 Région promotrice (Cdxl) rs7139166 Région promotrice (-1521) rs4516035 Région promotrice (-1012) rs2238136 Intron 1 (-4817) rsl0735810 Exon2(FoJtI) rs 1544410 Intron 8 (Bsml) rs7975232 Intron 8 (Apal) rs731236 Exon 9 ( r ^ I ) rs703842 Région promotrice (-1918) rs 10877012 Région promotrice (-1260) rs3782130 Région promotrice (-1077) rs7041 Exon 11 ( Asp416Glu) rs4588 Exon 11 (Thr420Lys) Les SNP sont identifiés par leur numéro dbSNP sur le site http://www.ncbi.nlm.nih.gov/SNP/.
VDR
Les effets de la vitamine D nécessitent la liaison entre le récepteur de la vitamine D (VDR) et son ligand l,25(OH)2D, permettant par la suite la régulation de la croissance cellulaire, de la différenciation, de l'apoptose, et de plusieurs mécanismes cellulaires impliqués dans la carcinogenèse (revu dans (15)). Plus précisément le complexe VDR-l,25(OH)2D formerait un hétérodimère avec le récepteur X rétinoïde (RXR) qui à son tour se lierait aux sites de liaison répondant à la vitamine D (VDREs pour Vitamin D Responsive Elements) dans la région promotrice de plusieurs gènes et en régulerait alors l'expression.
L'utilisation d'un modèle animal a permis d'établir que les effets de la vitamine D sur la glande mammaire nécessitent la présence de son récepteur (VDR) (revu dans (11)). En employant des souris transgéniques déficientes pour le gène du VDR (VDR KO), certains auteurs ont démontré que la présence de ce gène réduit la quantité de structures de la glande mammaire affectées par la carcinogenèse. En effet, comparées aux souris normales, les souris VDR KO avaient des glandes mammaires plus lourdes, une augmentation de la ramification et de l'extension canalaire ainsi qu'un plus grand nombre de bourgeons cellulaires (16). De plus durant la grossesse des souris VDR KO, le développement des bourgeons cellulaires était devancé et leur régression apoptotique associée à l'involution
était retardée (17). Une relation dose-réponse entre le VDR et certains de ses effets a aussi été démontrée en croisant des souris VDR KO avec des souris transgéniques MMTV-neu à plus fort risque de cancer de la glande mammaire. La présence d'une seule copie d'un
VDR déficient était suffisante pour raccourcir la période de latence et augmenter l'incidence des tumeurs mammaires (18). Cette information est particulièrement intéressante lorsque l'on sait que chez les femmes nullipares, la glande mammaire est principalement composée de bourgeons cellulaires (i.e. alvéoles au stade non différencié) et lobules de type 1 (constitués de 5 à 6 alvéoles), et que ceux-ci seront progressivement remplacés par des lobules plus gros et plus différenciés (types 2 et 3) en fonction du nombre de grossesses (19, 20). Les bourgeons cellulaires et lobules de type 1 sont les structures ayant le taux de mitoses le plus élevé et selon certains auteurs ils sont vraisemblablement à la base de l'explication biologique reliant la faible parité à l'augmentation du risque de cancer du sein (revu dans (21)). La démonstration que le récepteur de la vitamine D puisse influencer la structure même de la glande mammaire en fait donc un gène candidat de premier ordre pour l'étude de l'étiologie du cancer du sein.
Le gène VDR, exprimé dans les cellules épithéliales mammaires normales ou cancéreuses (22), contient plusieurs polymorphismes dont certains ont été analysés en relation avec le cancer du sein (revus dans (23)). La région 3'UTR et les nombreux SNP qui la précèdent (e.g. Bsml, Apal et Taqï) ont été beaucoup étudiés. Par contre, à l'exception du polymorphisme sur le codon de départ de l'exon 2 (i.e. Fokl), les autres SNP près de la région 5'UTR (renfermant la région promotrice) du VDR l'ont été beaucoup moins.
On a retenu les SNP Bsml (rsl544410) et Fokl (rsl0735810) puisqu'il sont situés suffisamment loin l'un de l'autre sur le gène pour ne pas être en déséquilibre de liaison (i.e. qu'ils n'auront pas tendance à être transmis ensemble), permettant dès lors d'en étudier les effets indépendants sur le cancer du sein (24). Ces deux candidats sont potentiellement fonctionnels, directement ou indirectement. Le polymorphisme Fokl se trouve sur l'exon 2 du gène et ne serait en déséquilibre de liaison avec aucun autre SNP du VDR. Chez les porteuses du génotype ff, la protéine formant le récepteur VDR est plus courte de trois acides aminés, de plus son efficacité réduite a été démontrée (revu dans (24)). Quand au Bsml, bien qu'il soit situé sur un intron (intron 8) et que par conséquent il ne devrait pas être fonctionnel en soi, ce polymorphisme est en déséquilibre de liaison complet avec le polymorphisme Taql, lui-même situé sur un exon (exon 9), et aussi avec la région 3'UTR du gène. Cette dernière région, quoique non transcrite, contient le microsatellite poly(A) et serait impliquée dans la stabilité de l'ARN messager ou dans sa traduction en protéines
moins fonctionnelles (revu dans (24)). Ceci fait de Bsml un marqueur de régions potentiellement fonctionnelles.
Quand aux autres SNP du VDR analysés dans cette thèse (rsl 1568820, rs7139166, rs4516035, rs2238136), et présentés au tableau 1, ils ont été peu étudiés. Cependant leur position dans ou près de la région promotrice du gène fait en sorte qu'ils sont également susceptibles d'être fonctionnels puisque cette région est importante pour la régulation de la transcription du gène et de sa traduction éventuelle en protéines (revu dans (13)).
CYP27B1
Avant de pouvoir se lier à son récepteur, la vitamine D (sous forme de 25(OH)D) doit être transformée en l,25(OH)2D puisque c'est ce metabolite qui constitue le ligand principal du VDR. Cette transformation est catalysée par l'enzyme 25(OH)D 1 a-hydroxylase ou cytochrome P450, famille 27, sous-famille B, polypeptide 1 (CYP27B1), et survient principalement dans le rein (revu dans (1, 25)) bien que les cellules mammaires humaines puissent également faire cette transformation localement (26, 27). On sait d'autre part qu'une déficience de l'enzyme 25(OH)D la-hydroxylase chez les patients atteints de rachitisme héréditaire vitaminodépendant de type I (pseudo carence en vitamine D) entraîne des niveaux très bas de l,25(OH)2D et des symptômes importants associés au manque de vitamine D tels de l'hyperparathyroïdie, des retards de croissance, des anomalies osseuses, etc. Ces anomalies peuvent disparaître complètement suite aux traitements avec de la
l,25(OH)2D (revu dans (25)). De plus l'utilisation de souris transgéniques déficientes pour le gène 25(OH)D la-hydroxylase (CYP27B1 KO) (28) a permis de reproduire les effets caractéristiques du rachitisme héréditaire vitaminodépendant de type I.
Le gène CYP27B1 possède des polymorphismes qui sont susceptibles de réduire les niveaux circulants ou intracellulaires de l,25(OH)2D. Aucun de ces SNP n'a été analysé au regard de la densité mammaire. Nous en avons étudié trois potentiellement fonctionnels puisque localisés dans la région promotrice du gène (cf. Tableau 1)
DBP
La vitamine D étant une hormone liposoluble elle doit se lier à une protéine de transport pour cheminer le long de sa voie métabolique, c'est-à-dire de son lieu de production ou d'absorption (i.e. la peau ou l'intestin) jusqu'à son récepteur se retrouvant dans différents
organes. Environ 90% de la 25(OH)D et de la l,25(OH)2D plasmatique serait lié à la protéine de transport de la vitamine D (DBP) (29). Il existe trois principales variantes phorétiques de cette protéine, aussi appelée protéine Gc (i.e. Gc-1-1, Gc-1-2, Gc-2-2), dont certaines ont été associées à des concentrations plus faibles de protéines de transport (30) ou à des concentrations plus faibles de la 25(OH)D (31).
Des études avec des souris transgéniques déficientes en DBP (DBP KO) ont démontré que chez ces animaux la demi-vie de la vitamine D était réduite, et ses metabolites non actifs (e.g. 24,25(OH)2D) étaient excrétés en plus grande quantité dans l'urine, diminuant par conséquent les niveaux sanguins de 25(OH)D (32).
À notre connaissance les effets des polymorphismes du gène de la protéine de transport de la vitamine D (DBP) n'ont jamais été étudiés en relation avec la densité mammaire, non plus qu'avec les concentrations circulantes de la 25(OH)D chez des femmes préménopausées en santé. Pour nos études nous avons retenu deux SNP potentiellement fonctionnels situés sur l'exon 11 de ce gène, ceux-ci changent la chaîne d'acides aminés donc la structure primaire de la protéine de transport (cf. Tableau 1).
Cancer du sein, vitamine D et polymorphismes du VDR
Dans la première partie de l'état des connaissances ci-haut, nous avons sommairement résumé le métabolisme de la vitamine D chez l'humain en mettant l'accent sur les variations interindividuelles qui peuvent s'exprimer aux diverses étapes de cette voie métabolique. Nous avons aussi tenté d'expliquer les fondements génétiques de ces variations métaboliques en mettant l'accent sur les quelques polymorphismes que nous avons étudiés. Dans les trois prochaines parties de cette section sur l'état des connaissances, nous résumons les connaissances spécifiques à chacune des trois études qui ont fait l'objet de cette thèse. Cette partie traitera du cancer du sein et de sa relation avec la vitamine D et certains polymorphismes de son récepteur. La prochaine traitera de la densité mammaire et de sa relation avec la vitamine D et avec certains polymorphismes de sa voie de signalisation. Enfin la dernière partie portera sur la 25-hydroxyvitamine D (25(OH)D) et de sa relation avec sa protéine de transport et certains polymorphismes de cette dernière. Le cancer du sein est une cause importante de morbidité et de mortalité chez les femmes. Les estimations actuelles indiquent qu'une canadienne sur 9.5 sera atteinte d'un cancer du
sein pendant sa vie et une femme sur 26 mourra des suites de ce cancer. En 2009 on estimait qu'au Canada, environ 5 400 décès lui seraient attribuables (33) Bien que le taux de mortalité normalisé selon l'âge soit relativement stable depuis 1971 et qu'on enregistre une légère baisse du taux de mortalité depuis 1993, les taux d'incidence du cancer du sein continuent pour leurs parts de connaître une augmentation significative. Ceux-ci ont augmenté d'environ 28 % durant les 25 dernières années au Canada. Selon les statistiques de 2004, la progression des taux a été constante et régulière entre 1973 et 2004 et a affiché peu de fluctuations (34). Depuis, le taux d'incidence est demeuré relativement stable (33). Bien que la détection précoce du cancer du sein suite aux mammographies de dépistage puisse avoir joué un rôle dans cette progression, les causes de l'augmentation des taux jursqu'en 2004 demeurent en grande partie inconnues. En effet seulement 25% à 40% des cas seraient attribuables aux facteurs de risque reconnus du cancer du sein (35). Les recherches se poursuivent donc pour découvrir ou documenter d'autres facteurs de risque méconnus du cancer du sein.
Plusieurs études épidémiologiques à devis écologiques ont décrit que l'augmentation de la latitude ou la baisse des niveaux de rayonnements UVB étaient associés à une augmentation de l'incidence du cancer du sein (36-40). De telles observations ont suggéré que la vitamine D, principalement synthétisée par la peau sous l'effet du soleil et qui enregistre une forte variation saisonnière aux latitudes élevées, était impliquée dans ce type de cancer. Des études à devis différents telles les études cas-témoins et de cohortes (41-49) ont également associé de faibles apports ou niveaux circulants de vitamine D et le cancer du sein. Un essai clinique randomisé récent, durant lequel les niveaux de 25(OH)D ont atteint plus de 80 nmol/L, a d'ailleurs montré une baisse du risque de plusieurs cancers (5) parmi lesquels le cancer du sein était le plus fréquent. Bien que critiqués quand à la randomisation de leur étude, les auteurs rapportent que leurs résultats significatifs s'expliquent vraisemblablement par l'utilisation d'une dose suffisante pour rehausser le niveau de base de vitamine D de 24 nmol/L (i.e. + 33%) par rapport aux autres essais cliniques ayant généralement une faible augmentation relative de la vitamine D circulante. Selon eux, dans l'étude récente du Women's Health Initiative (WHI) utilisant une supplementation de 400 UI de vitamine D (50) et ayant produit des résultats négatifs, le niveau de 25(OH)D n'a pu être augmenté que de 42 à 47 nmol /L (i.e. + 12%) On peut ajouter enfin que plusieurs études suggèrent que la vitamine D réduit la progression ou l'agressivité du cancer du sein (45, 51-54).
Tel que décrit à la section précédente, puisque les polymorphismes du récepteur (VDR) peuvent en réduire l'efficacité, la protection contre le cancer du sein associée à la vitamine D pourrait varier pour les porteuses des certains génotypes définis par les deux SNP potentiellement fonctionnels que nous avons retenus, c'est à dire Fokl et Bsml. Si la vitamine D affecte le risque de cancer du sein et que ces polymorphismes sont effectivement fonctionnels, on pourrait s'attendre à ce que le risque de cancer du sein diffère selon les polymorphismes (génotypes) Fokl et Bsml du VDR.
Seules quelques études épidémiologiques ont analysé la relation entre ces deux SNP et le risque de cancer du sein. Elles étaient cependant généralement de petite taille (< 500 cas ou témoins) et souvent conduites dans des populations d'ethnicités différentes, ce qui explique peut-être les résultats variables. Dans le cas du Fokl on a recensé huit articles (55-62) dont deux publiés après la fin de nos analyses. Trois de ces articles, émergeant vraisemblablement d'un même échantillon de femmes caucasiennes, rapportaient une baisse de risque associée à l'allèle rare (f) (56, 59, 60). Parmi les études antérieures, deux (58, 61) n'observaient aucune association alors que la plus grosse à l'époque concluait à une hausse de risque associée à l'allèle rare (f) (57).
En ce qui concerne Bsml, nous avions répertorié 11 études avant de procéder à nos analyses, (56, 57, 59-67), et deux par la suite (68, 69). Parmi les premières, seulement deux attribuaient une augmentation de risque à l'allèle B et elles avaient été conduites dans des populations Hispanique (61) et Taiwanaise (67) ayant des fréquences d'alleles très différentes de celles des populations caucasiennes (i.e. 43 % et 91 % de bb respectivement chez les témoins comparativement à 36 % dans notre échantillon). Les premières études, plus petites (56, 60, 66), rapportaient des accroissements de risque associés au génotype bb alors que les études plus grosses et plus récentes (57, 64) ne confirmaient pas les résultats significatifs initiaux.
Ainsi, la relative rareté des études sur la question et les contradictions dans les résultats publiés nous ont incités à réaliser l'étude sur les risques de cancer du sein en fonction des polymorphismes Fokl et Bsml du récepteur (VDR); les résultats sont présentés au chapitre II de la présente thèse. De même, cette étude fut l'occasion d'explorer l'effet modifiant de diverses charactéristiques des femmes incluant les antécédents familiaux de cancer du sein sur la relation entre les polymorphismes du VDR et le risque de cancer du sein; question non encore abordée par la communauté scientifique.
Densité mammaire, vitamine D et polymorphismes de sa voie de
signalisation
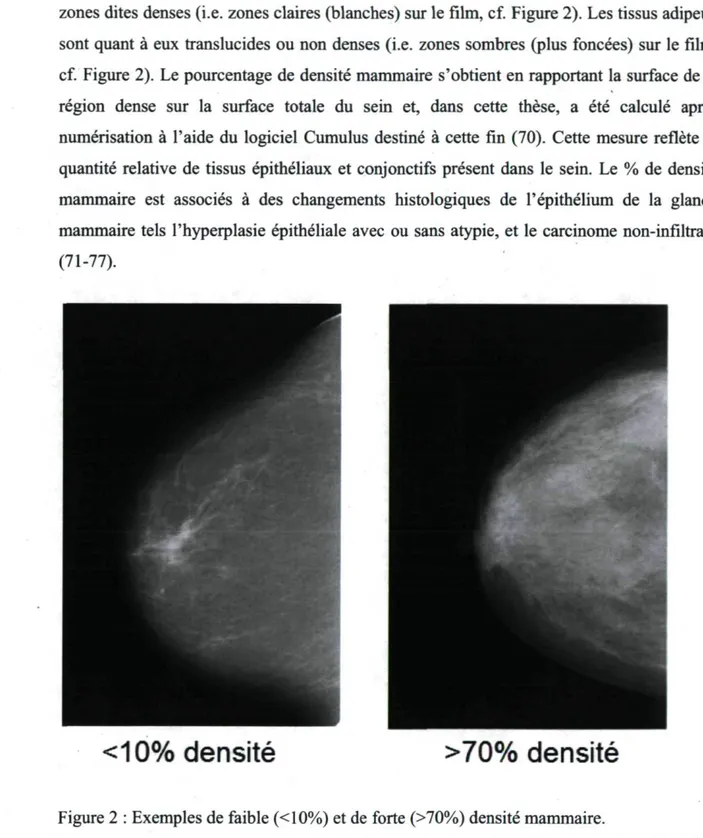
Sur une mammographie, ce sont les tissus épithéliaux et conjonctifs qui produisent les zones dites denses (i.e. zones claires (blanches) sur le film, cf. Figure 2). Les tissus adipeux sont quant à eux translucides ou non denses (i.e. zones sombres (plus foncées) sur le film, cf. Figure 2). Le pourcentage de densité mammaire s'obtient en rapportant la surface de la région dense sur la surface totale du sein et, dans cette thèse, a été calculé après numérisation à l'aide du logiciel Cumulus destiné à cette fin (70). Cette mesure reflète la quantité relative de tissus épithéliaux et conjonctifs présent dans le sein. Le % de densité mammaire est associés à des changements histologiques de l'épithélium de la glande mammaire tels l'hyperplasie épithéliale avec ou sans atypie, et le carcinome non-infiltrant (71-77).
* ^ _ f e
m
^F
<10% densité
>70% densité
Figure 2 : Exemples de faible (<10%) et de forte (>70%) densité mammaire.
Les zones claires sont composées principalement de tissus épithéliaux et conjonctifs. Elles constituent la partie dense du sein. Les zones foncées sont composées principalement de tissus adipeux non radio-opaques. Elles constituent la partie non dense du sein.
Plusieurs études ont démontré que la densité mammaire est fortement associée au risque de cancer du sein (revue dans (78, 79)). Cette relation est illustrée à la figure 3 où l'on retrouve l'augmentation du risque relatif de cancer du sein en fonction du pourcentage de densité évalué par un même chercheur dans trois études cas-témoins différentes (80). Une étude cas-témoin récente montre également que la densité mammaire est un facteur de risque indépendant des concentrations d'hormones circulantes, et non simplement le reflet de celles-ci (81). La densité mammaire réagit relativement rapidement à divers stimuli, comme le démontrent les hausses suivant le début de l'hormonothérapie de remplacement (82) ou la baisse suivant la prise de tamoxifène (83), elle est de plus en plus reconnue comme marqueur précoce du risque de cancer du sein ce qui favorise son utilisation dans l'étude de l'étiologie de ce cancer (84).
0 f - 6-11 5 -* e 4 - t w (/) q iE J' 2 - *
u>+
lf
-. • • I I 1 - 0-< —i t t 1 1 20 40 60 Densités mammaires (%) 80 100Figure 3 : Association entre les densités mammaires et le risque relatif de cancer du sein obtenue par un même chercheur regroupant les données de trois études cas-témoins.
Chaque point représente un risque relatif comparant les femmes n'ayant aucune densité avec celles catégorisées par classe de densité de 5% à l'exception de la première (1-9%) et de la dernière classe (90-100%). Les risques relatifs ont été obtenus par régression logistique et sont ajustés pour l'âge et le poids. La courbe en pointillé est une fonction spline qui modélise les risques relatifs. Les lignes pleines sont les intervalles de confiance à 95% autour de chaque risque relatif prédit par la fonction spline.
Les apports en vitamine D ont été associés à des baisses de densité mammaire dans quelques études (85-88) y compris dans la population Québécoise (6). De plus dans cette dernière on a démontré une association entre les variations saisonnières de 25(OH)D et les variations saisonnières des densités mammaires (7). Par contre d'autres études n'ont pas pu mettre en évidence une relation entre la vitamine D et les densités mammaires (89-93). Il semble plausible que les polymorphismes de la voie de signalisation de la vitamine D influencent les densités mammaires, toutefois cette relation n'a jamais été étudiée jusqu'à maintenant. Compte tenu des associations entre les deux SNP du récepteur à la vitamine D (i.e. Fokl et Bsml) et le cancer du sein dans nos données, on a vérifié leur association avec la densité mammaire. De plus nous avons retenu d'autres gènes candidats potentiellement importants (94) sur la même voie de signalisation dans la mesure où ils se situaient dans la région promotrice ou sur un exon du gène, ou encore lorsqu'ils avaient été associés au risque de cancer du sein ou à ses facteurs de risque. Six autres SNP sur le VDR ont donc été analysés en plus des deux précédents. Les quatre premiers se situent sur le promoteur ou en déséquilibre de liaison avec lui, (i.e. rsl 1568820, rs7139166, rs4516035, rs2238136). Les deux autres, localisés près de la région 3'UTR du gène sur l'exon 9 (Taql : rs731236) ou en déséquilibre de liaison avec lui (Apal : rs7975232), ont déjà été associés au risque de cancer du sein (23).
Tel que décrit précédemment le ligand principal du VDR est la 1,25-dihydroxyvitamine D (l,25(OH)2D), et sa production nécessite l'hydroxylation de la 25(OH)D grâce à l'enzyme émanant du cytochrome P450, famille 27, sous-famille B, polypeptide 1 (CYP27B1). Cette enzyme, impliquée dans la production locale de 1,25(OH)2D dans les cellules mammaires (26, 27), est susceptible d'influencer la densité mammaire et possède des polymorphismes génétiques parmi lesquels nous avons étudié trois SNP localisés dans la région promotrice du gène (rs703842, rsl0877012, rs3782130).
Enfin, le gène codant pour la protéine de transport (DBP) de la vitamine D et de ses metabolites se retrouve en amont des SNP précédents sur la voie de signalisation et, tel que démontré par des études avec des souris transgéniques, cette protéine est nécessaire au maintien de niveaux stables et au fonctionnement adéquat de la vitamine D (32). Les SNP DBP-l (rs7041) et DBP-2 (rs4588) situés sur l'exon 11 ont été analysés en relation avec la densité mammaire. Ce dernier polymorphisme a récemment été associé aux concentrations
circulantes de 25(OH)D chez des patients atteints de la maladie de Graves (95), de même que parmi des femmes témoins postménopausées (55).
En somme les 13 SNP décrits ci-dessus sont susceptibles d'influencer la densité mammaire à la mammographie via leur effet potentiel sur les protéines impliquées tout au long de la voie de signalisation de la vitamine D et font l'objet du chapitre III.
25-hydroxyvitamine D, protéine de transport et polymorphismes de DBP
Tel que décrit précédemment, la 1,25-dihydroxyvitamine D (l,25(OH)2D) est le metabolite actif de la vitamine D liposoluble, et celui qui se lie avec le plus d'affinité au VDR. Par contre la l,25(OH)2D reflète moins bien le statut vitaminique que la 25-hydroxyvitamine D (25(OH)D) parce que sa concentration circulante est étroitement contrôlée par des mécanismes de rétroaction impliquant entre autres l'hormone parathyroïdienne (PTH), le calcium, le phosphore, et que par conséquent elle varie peu dans la circulation (1, 96). La 25(OH)D est plus influencée par les variations d'exposition provenant de l'alimentation, des suppléments, ou de l'exposition au soleil, et peut être transformée localement en l,25(OH)2D, par conséquent elle est reconnue comme le meilleur marqueur du statut en vitamine D.La protéine de transport de la vitamine D (DBP) est une protéine faisant partie de la famille des albumines et des alpha-fétoprotéines. Ces protéines hydrosolubles sont essentielles au transport et à l'incorporation cellulaire des hormones ou vitamines liposolubles (revu dans (97)). On estime que plus de 99% de la 25(OH)D serait liée aux protéines plasmatiques dont environ 90% à la protéine de transport de la vitamine D (DBP), la majorité de la partie restante étant liée à l'albumine (29, 98). DBP est synthétisée dans le foie, où se produit également l'hydroxylation de la vitamine D en 25(OH)D. Le rôle exact que joue la protéine DBP dans les effets de la vitamine D est encore incertain, toutefois il semble que ses niveaux ne soient pas influencés par les niveaux de la vitamine D ou des metabolites de la vitamine D, ni par d'autres hormones calciotropiques (e.g. PTH, calcitonine), ou encore par les variations saisonnières d'ensoleillement (revu dans (99)). Par contre les niveaux de DBP augmentent de manière significative durant la grossesse et l'hormonothérapie de remplacement (100-102), et diminuent en présence de maladies du foie (103, 104), du syndrome néphrotique et de malnutrition, probablement résultant d'une diminution de la synthèse hépatique ou d'une proteinuric excessive (revu dans (99)).
Tel que décrit précédemment il existe trois phénotypes principaux de la protéine de transport et, dans les populations caucasiennes, le phénotype Gc2-2 a été associé à des niveaux de DPB plus faibles (30) de même qu'à des niveaux de 25(OH)D inférieurs (31). En plus de cet effet potentiel sur les niveaux circulants, les divers phénotypes de DBP pourraient influencer l'entrée de la 25(OH)D dans les cellules mammaires par l'intermédiaire d'un mécanisme d'endocytose similaire à celui mis en évidence dans le rein (97, 105). Dans ce cas ce serait le complexe 25(OH)D-DBP, plutôt qu'uniquement l'hormone sous forme libre, qui pénétrerait dans la cellule mammaire. Il est donc plausible que les phénotypes de la protéine de transport affectent la performance de l'endocytose de la 25(OH)D.
Les différences de poids moléculaires des divers phénotypes de la protéine de transport de la vitamine D reflètent vraisemblablement des différences structurelles tributaires des variations génétiques (SNP) du gène de la protéine de transport. La présence de SNP fonctionnels sur le gène codant la DBP est donc susceptible d'influencer le transport ou le devenir de la vitamine D. Puisque la protéine DBP circule à des concentrations environ 100 fois plus élevées que la 25(OH)D et que sa demi-vie est courte (2-3 jours) relativement à celle de la 25(OH)D (1-2 mois) (97, 99, 106), toute réduction de la stabilité ou de l'affinité de la protéine de transport DBP pourrait affecter les concentrations circulantes de 25(OH)D ou son incorporation cellulaire et, par conséquent, la manifestation de maladies liées à l'hypovitaminose D.
L'absence d'étude sur le sujet a motivé l'analyse de la relation entre deux SNP, DBP-l (rs7041) et DBP-2 (rs4588) situés sur l'exon 11 du gène de la protéine de transport, et les concentrations circulantes de 25(OH)D. Les résultats de cette étude sont présentés au chapitre IV.
FORCES ET LIMITES DES ÉTUDES D'ASSOCIATION
GÉNÉTIQUE
Avant de passer à la description de la méthodologie utilisée pour chacune des études présentées dans le corps de la thèse, la présente section discute des forces et des limites des études d'association génétique puisque les études menées dans le cadre de ce doctorat sont toutes de ce type.
On sait qu'une histoire familiale de cancer du sein est associée à une augmentation du risque de cette maladie. L'hérédité semble donc jouer un rôle significatif dans son étiologie. En effet, comparé à celui d'une femme n'ayant pas d'histoire familiale de cancer du sein, le risque relatif (RR) d'une femme dont la mère a eu ce cancer diagnostiqué après l'âge de 60 ans est de 1.4, alors que ce RR passe à 4-6 pour une femme ayant 2 parents du 1er degré atteints (par exemple une mère et une sœur ou deux sœurs) (107).
Les études génétiques familiales ont permis de mettre en évidence les associations entre des gènes spécifiques à forte penetrance comme BRCA1 et BRCA2 et le cancer du sein. A titre d'exemple, une femme de 40 ans porteuse d'un mutation sur le BRCAl a un risque de cancer du sein environ 15 fois plus élevé que celui d'une femme non porteuse n'ayant aucune histoire familiale de la maladie (107). Toutefois les cancers 'héréditaires' découlant de ce type de gènes à transmission autosomale dominante ne représenteraient que 5 à 10% de tous les cancers du sein diagnostiqués (revu dans (107)).
Ces résultats suggèrent que des gènes de prédisposition ayant une plus faible penetrance, des facteurs non génétiques tels l'environnement et les habitudes de vie, ou encore des interactions entre les deux, pourraient être impliqués dans l'étiologie d'une forte proportion de cancers du sein. Toutefois, lorsque l'on cherche à identifier des gènes à faible penetrance liés au risque de cancer du sein, les études génétiques familiales souffrent généralement d'un manque de puissance (14, 108). À cette fin, les études d'association génétique conduites dans la population générale sont plus appropriées puisque les effets des gènes à faible penetrance ne donnent pas lieu à des agrégations familiales fortes. En général, dans les études d'association génétique, on compare la fréquence des génotypes (c'est-à-dire des variations génétiques (e.g. SNP) sur un locus d'un gène donné) entre des cas et des témoins (109-112). Lorsque la distribution des génotypes diffère entre ces deux groupes, il y a association allélique entre le SNP et la condition étudiée. En somme ces études d'association recherchent des effets moins spectaculaires (risques relatifs souvent inférieurs
à 2) que ceux associés aux gènes à forte penetrance. Toutefois puisque certains SNP sont communs, la fraction attribuable qui découle d'une association avec un tel SNP peut être élevée et présenter un intérêt supérieur du point de vue de la santé publique.
Les études d'association génétique ont souvent donné des résultats qui n'ont pas été reproduits ultérieurement dans d'autres échantillons ou populations. Plusieurs difficultés classiques s'appliquant à 1'epidemiologic peuvent potentiellement expliquer cet état de fait (14, 112-114), cependant quelques particularités associées à 1'epidemiologic génétique méritent une brève description. Ci-dessous nous allons revoir les principales raisons évoquées pour expliquer cette reproductibilité difficile et décrire par la suite les stratégies employées pour tenter d'y remédier :
1) Erreur de type 1 : La présence d'erreur de type 1 (i.e. faux positif), possiblement exacerbé par le biais de publication, peut expliquer en partie la faible reproductibilité de certaines études. En effet, une étude d'association analysant plusieurs SNP, et ne corrigeant pas pour les comparaisons multiples, est susceptible de rapporter des résultats statistiquement significatifs attribuables essentiellement au hasard. Le fait de restreindre le choix des gènes étudiés uniquement aux gènes ayant une relation biologique plausible avec l'issue d'intérêt (i.e. approche des gènes candidats), de même que la replication des résultats obtenus antérieurement, permettent de réduire ce type d'erreur.
2) Erreur de type 2 : Le manque de puissance entraînant des faux négatifs est souvent invoqué pour expliquer la non replication de plusieurs résultats. Les études de replication conduites dans la même population avec des effectifs importants et une puissance suffisante devraient permettre de vérifier la reproductibilité de résultats initiaux. La puissance de ces études dépend de la taille de l'échantillon et de l'effet attendu, mais également de la fréquence de la variation génétique étudiée dans la population (115). Une puissance adéquate est plus facile à atteindre pour des SNP dit communs, par exemple lorsque l'allèle rare présente une fréquence supérieure à 5% (111).
3) Effets modifiants : Lorsqu'il s'agit d'associations qui varient substantiellement d'un sous-groupe de la population à un autre (c'est-à-dire en présence d'interactions importantes), la replication des résultats présente un obstacle additionnel. Compte tenu de la variabilité ethnique, génétique et comportementale de diverses populations, il est possible qu'une replication réalisée dans une population différente de celle faisant l'objet de l'étude initiale, nécessite des effectifs différents (supérieurs ou inférieurs)