Sex dependent effects on breathing and apnea following
chronic caffeine and acute progesterone administration
in newborn rats
Mémoire
Nagapraveena Uppari
Maîtrise en neurosciences - avec mémoire
Maître ès sciences (M. Sc.)
Sex dependent effects on breathing and apnea
following chronic caffeine and acute progesterone
administration in newborn rats
Mémoire
NagaPraveena UPPARI
Sous la direction de :
Aida Bairam, directrice de recherche
Vincent Joseph, codirecteur de recherche
Résumé
La caféine, une puissante stimulant respiratoire, est largement utilise pour traiter l'apnée du prématuré. Cependant, elle réduit mais n'élimine pas l'apnée et on ignore si ces effets sont spécifiques au sexe. Récemment, la progestérone qui est également un stimulant respiratoire a été proposée pour traiter l'apnée qui persiste à la caféine. Nos données antérieures chez le rat nouveau-né indiquent que la progestérone améliore la réponse ventilatoire à l'hypoxie et réduit la fréquence de l'apnée. Ici, nous avons testé l'hypothèse que l'ajout de progestérone améliore la respiration et diminue la fréquence de l'apnée de manière sexe-dépendant chez le rat de 12 jours de vie et sous traitement quotidien à caféine, telle qu'elle est utilisée en clinique. Nos résultats démontrent que chez le rat mâle préalablement traité avec de la caféine, la progestérone diminue la ventilation et augmente la fréquence de l’apnée en condition de normoxie et en réponse à l’hypoxie. Par contre, chez la femelle, la progestérone diminue la fréquence respiratoire seulement en réponse à l'hypoxie. Cependant, l’administration de la caféine en normoxie comme en hypoxie a réduit de maniére plus importante la fréquence de l’apnée chez le rat femelle que chez le mâle. Ces résultats indiquent une meilleure efficacité de la caféine dans le contrôle de l'apnée chez le rat nouveau-né femelle. Les mécanismes sous-jacents de ces observations sexe spécifique de la caféine sur les irrégularités respiratoires doivent être étudiés plus en détail. De plus, nos résultats suggèrent que l'administration chronique de la caféine à des rats nouveau-nés peut induire des changements au niveau des neurotransmetteurs qui modifient le rôle de la progestérone comme stimulant respiratoire.
Abstract
Caffeine is a potent respiratory stimulant and a widely used treatment for apnea of prematurity. However, caffeine reduces, but does not eliminate apnea and it is not known if these effects are sex-specific. Recently, progesterone which is also a respiratory stimulant, has been proposed to treat apnea persistent with caffeine. Our previous data in newborn rats indicate that progesterone enhances the ventilatory response to hypoxia and reduces apnea frequency. Here, we tested the hypothesis that the addition of progesterone to daily caffeine administration, as used in clinic, enhances respiration and decreases apnea frequency in a sex-dependent manner in 12 day old rats. Our results show that, in rats treated with caffeine progesterone depressed ventilation under normoxia and in response to hypoxia. Whereas in female rats, progesterone decreased the respiratory frequency only in response to hypoxia. However, during normoxia and hypoxia, chronic caffeine administration reduced apnea frequency more in female than male rats. These results indicate a better efficiency of caffeine in controlling apnea in newborn female rats. The underlying mechanisms of these sex-specific effects of caffeine on breathing irregularities need further investigation. In addition, our results suggest that chronic caffeine administration to newborn rats may induce changes in neurotransmitters milieu that consequently affect the role of progesterone as a respiratory stimulant.
Table of contents
Résumé ... iii
Abstract ... iv
Table of contents ... v
List of figures ... ix
List of tables ... xii
List of abbreviations ... xiv
Acknowledgements ... xvii
Avant-propos ... xix
Preface ... xxi
Introduction ... 1
A. Structures involved in control of breathing ... 1
A.1 Hypoxic ventilatory response (HVR) in newborn ... 3
A.2 Implication of neurotransmitters in control of breathing ... 3
A.2.1 Gamma-aminobutyric acid (GABA) ... 3
A.2.2 Adenosine ... 4 B. Breathing Instability ... 4 B.1 Types of Apnea ... 5 B.2 Apnea Treatment ... 6 B.2.1 Xanthine Therapy ... 6 B.2.2 Failure of Methylxanthines ... 6 C. Progesterone ... 7 C.1 Progesterone receptors ... 7
C.1.1 Nuclear Progesterone Receptors (nPR) ... 7
C.1.2 Membrane Progesterone Receptors (mPR) ... 8
C.2 Progesterone as a respiratory stimulant ... 11
C.2.1 In adults ... 11
C.2.2 In newborn ... 11
D.1 In Neonates ... 12
D.2 In newborn animals ... 13
E. Hypothesis ... 14
E.1 Objectives ... 14
Materials and Methods ... 15
F Animals ... 15
F.1 Neonatal gavage with water or caffeine ... 15
F.1.1 Drug preparation and administration ... 16
F.1.2 Progesterone ... 16
F.1.3 Bicuculline ... 16
F.1.4 Allopregnanolone ... 16
F.1.5 Finasteride: ... 17
F.2 Measurement of Respiratory and Metabolic Parameters ... 17
F.2.1 Plethysmography Recording... 19
F.2.2 Analysis of ventilatory parameters and respiratory irregularities ... 20
F.2.3 Circulating progesterone and allopregnanolone levels ... 20
F.2.4 Statistical analysis ... 21
CHAPTER I ... 22
Inhibitory respiratory responses to progesterone and allopregnanolone in newborn rats chronically treated with caffeine. ... 22
1.1 Abstract... 23
1.2 Résumé ... 24
1.3 Introduction... 25
1.4 Materials and methods... 26
1.4.1 Animals ... 26
1.4.2 Neonatal gavage with water or caffeine ... 26
1.4.3 Respiratory measurements ... 27
1.4.4 Circulating progesterone and allopregnanolone levels ... 30
1.5.1 Physiological and ventilatory parameters under normoxic conditions ... 31
1.5.2 Effects of progesterone injection on apnea frequency under normoxia in control and caffeine-treated rats ... 33
1.5.3 Effects of progesterone injection on hypoxic ventilatory response in control and caffeine-treated rats ... 34
1.5.4 Apnea frequency in hypoxia ... 38
1.5.5 Effects of bicuculline in control and caffeine-treated rats ... 39
1.5.6 Effects of allopregnanolone in control and caffeine-treated rats ... 43
1.5.7 Effects of progesterone plus finasteride treatment in control and caffeine-treated rats ... 46
1.6 Discussion ... 50
1.6.1 Interplay between caffeine and gamma-aminobutyric acid (GABAA) receptor function ... 50
1.6.2 Indirect effect of progesterone on GABAA receptors ... 52
1.6.3 Progesterone reverses the effect of caffeine on apnea frequency in newborn rats .. 53
1.6.4 Other putative mechanisms ... 53
1.7 Conclusion and perspective ... 54
1.8 Acknowledgements ... 55
CHAPTER II ... 56
Respiratory responses to progesterone and allopregnanolone following chronic caffeine treatment in newborn female rats. ... 56
2.1 Abstract... 57 2.2 Résumé ... 58 2.3 Introduction... 59 2.4 Methods ... 60 2.4.1 Animals ... 60 2.4.2 Plethysmography ... 60
2.4.3 Drug preparation and dose ... 61
2.4.4 Respiratory analyses and statistics ... 62
2.4.5 Statistical analyses ... 62
2.5.1 Effects of progesterone in P12 female rats treated with water (control) or caffeine 63 2.5.2 Effects of allopregnanolone in P12 female rats treated with water (control) or
caffeine ... 68
2.5.3 Antagonisms of GABAa receptors blocked the effects of allopregnanolone on breathing frequency and apnea duration in P12 female rats treated with water (control) or caffeine ... 73
2.6 Discussion ... 74
2.6.1 Progesterone and basal breathing in P12 female rats ... 75
2.6.2 Progesterone and hypoxia in P12 female rats ... 76
2.7 Clinical usefulness of our observations ... 77
2.8 Acknowledgements ... 78
CHAPTER III... 79
3 General Discussion ... 79
3.1 Newborn rats as animal model... 79
3.2 Age and sex dependent differences with caffeine administration in newborn rats... 80
3.2.1 Acute administration ... 80
3.2.2 Chronic administration ... 80
3.3 Sex dependent differences with progesterone administration in 12-day old rats ... 81
3.3.1 Coherence between adenosine and GABA ... 82
3.3.2 Interconnection between progesterone and GABA... 83
3.4 Consideration for techniques and drugs: ... 86
Conclusion and Perspectives ... 88
List of figures
Figure 1. Elaboration of the structures involved in the control of breathing ... 2 Figure 2. Two representative examples of desaturation, secondary to apnea or respiratory
pauses, in preterm infants monitored via respiratory inductance plethysmography ... 5
Figure 3. Progesterone metabolism and receptor actions in brain ... 9 Figure 4. Biosynthetic Pathway of Progestogens ... 10 Figure 5. Percentage of infants diagnosed with AOP in different gestational age group .... 12 Figure 6. Schematic representation of whole-body plethysmography ... 18 Figure 7. Frequency of apneas recorded under normoxia after an i.p. injection of saline or
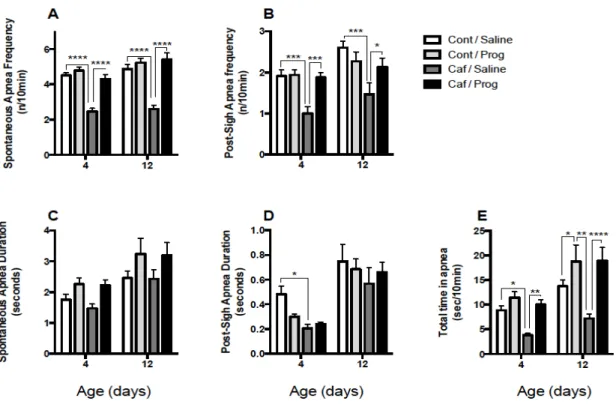
progesterone (Prog) in 4 and 12 days old rats that received a daily gavage of water (Cont) or caffeine (Caf) between postnatal days 3-12 ... 34
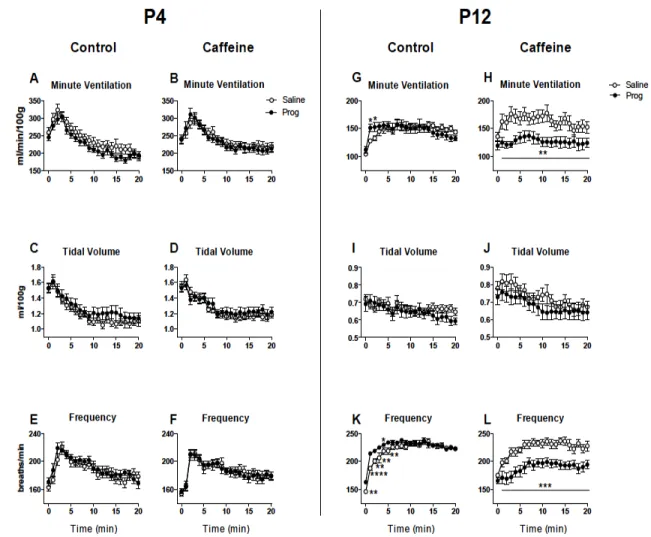
Figure 8: Hypoxic ventilatory response after an i.p. injection of saline or progesterone (Prog)
in 4 and 12 days old rats that received a daily gavage of water (Control) or caffeine between postnatal days 3-12... 36
Figure 9. Changes of CO2 production rate during hypoxic exposure and hypoxic ventilatory
response as and after an i.p. injection of saline or progesterone (Prog) in 4 and 12 days old rats that received a daily gavage of water (Control) or caffeine between postnatal days 3-12 ... 37
Figure 10. Frequency of apneas recorded during the last 10 minutes of the hypoxic exposure
after an i.p. injection of saline or progesterone (Prog) in 4 and 12 days old rats that received a daily gavage of water (Cont) or caffeine (Caf) between postnatal days 3-12. ... 39
Figure 11. Effects of the GABAA receptor antagonist bicuculline (DMSO=0, and 1, 2, 3
mg/kg – ip) on variables recorded in 12 days old rats that received a daily gavage of water (Cont) or caffeine (Caf) between postnatal days 3-12. ... 41
V.E V . CO2 V . E V . O2
Figure 12. Effects of the GABAA receptor antagonist bicuculline (DMSO=0, and 1, 2, 3
mg/kg – ip) on apnea in 12 days old rats that received a daily gavage of water (Cont) or caffeine (Caf) between postnatal days 3-12. ... 42
Figure 13. Effects of allopregnanolone (Allop, 10 mg/kg) or vehicle on variables recorded
in 12 days old rats that received a daily gavage of water (Cont) or caffeine (Caf) between postnatal days 3-12. ... 44
Figure 14. Effects of allopregnanolone (Allop, 10 mg/kg) or vehicle on apnea ) in 12 days
old rats that received a daily gavage of water (Cont) or caffeine (Caf) between postnatal days 3-12. ... 45
Figure 15. Effects of progesterone (4 mg/kg), or progesterone+finasteride (4 mg/kg +10
mg/kg ) on variables recorded in 12 days old rats that received a daily gavage of water (Cont) or caffeine (Caf) between postnatal days 3-12. ... 48
Figure 16. Effects of progesterone (4 mg/kg), or progesterone+finasteride (4 mg/kg +10
mg/kg) on apnea in 12 days old rats that received a daily gavage of water (Cont) or caffeine (Caf) between postnatal days 3-12. ... 49
Figure 17. Effects of progesterone on hypoxic ventilatory response in P12 female rats treated
with water (control) or caffeine. Respiratory frequency, tidal volume and minute ventilation in rats daily gavaged with water or with caffeine between the postnatal days 3-12. ... 65
Figure 18. Effects of progesterone on hypoxic metabolic response in P12 female rats treated
with water (control) or caffeine. ... 66
Figure 19. Effects of progesterone on frequency and duration of apnea during normoxia and
hypoxia in P12 female rats treated with water (control) or caffeine after an i.p. injection of saline or progesterone in 12 days old female rats that received a daily gavage of water (Control) or caffeine between postnatal days 3-12. ... 67
Figure 20. Effects of allopregnanolone on hypoxic ventilatory response in P12 female rats
treated with water (control) or caffeine. Respiratory frequency, tidal volume and minute ventilation in rats daily gavaged with water (control) or with caffeine between the postnatal days 3-12. ... 70
Figure 21. Effects of allopregnanolone on hypoxic metabolic response in P12 female rats
Figure 22. Effects of allopregnanolone on frequency and duration of apnea during normoxia
and hypoxia in P12 female rats treated with water (control) or caffeine. ... 72
Figure 23. Effects of bicuculline or bicuculline+allopregnanolone on ventilatory variables,
frequency and duration of apneas in P12 female rats treated with water (control) or caffeine. ... 74
Figure 24. Maturation of hypoxic ventilatory response in human and rats. ... 80 Figure 25. Effect of caffeine during development on mRNA expression levels of genes
coding for A1 and A2A adenosine receptors (ADORA1 and ADORA2) and for a1 and
a2 subunits of GABAA receptor (GABRA1 and GABRA2). ... 83
Figure 26: Time courses of the effects of caffeine on cerebrocortical steroid concentrations.
... 85
Figure 27. Schematic illustration showing the balance between allopregnanolone and
progesterone receptors for the control of apnea frequency in newborn rats. ... 85
Figure 28: The biosynthesis pathway of Progesterone. ... 87
List of tables
Table 1. Number of animals used in each study ... 15 Table 2. Body weight (BW), number of rats/group (n), and values of rectal temperature (Tr),
respiratory frequency (fR), tidal volume ( ), minute ventilation ( ), O2 consumption
rate ( ), CO2 production rate ( ), ventilatory equivalent for O2 ( ), and
CO2 ( ) and ventilatory quotient ( ) recorded under normoxia after an
i.p. injection of saline or progesterone in 4- and 12-day-old rats that received a daily gavage of water (Control) or caffeine between postnatal days 3-12. ... 32
Table 3. serum levels of progesterone and allopregnanolone measured after an i.p. injection
of saline or progesterone in 12 days old rats that received a daily gavage of water (Control) or caffeine between postnatal days 3-12. ... 33
Table 4. Body weight (BW), number of rats/group (n), respiratory frequency (fR), tidal
volume ( ), minute ventilation ( ), O2 consumption rate ( ), CO2 production rate
( ), ventilatory equivalent for O2 ( ), and CO2 ( ) recorded under
normoxia after an i.p. injection of saline, progesterone, vehicle (DMSO+cyclodextrine see text), or progesterone+finasteride (in vehicle) in 12 days old rats that received a daily gavage of water (Control) or caffeine between postnatal days 3-12... 47
Table 5. Body weight (g), number of rats/group (n), body temperature (°C) during baseline
(B) and hypoxia (H), respiratory frequency, tidal volume, minute ventilation, O2
consumption rate ( ), CO2 production rate ( ), ventilatory quotient ( / ),
ventilatory equivalent for O2 ( / ), and CO2 ( / ) recorded under normoxia
after an i.p. injection of saline or progesterone in 12-day-old female rats that received a daily gavage of water (Control) or caffeine from the postnatal days 3-12. ... 64
Table 6. Body weight (g), number of rats/group (n), body temperature (°C) during baseline
(B) and hypoxia (H), respiratory frequency, tidal volume, minute ventilation, O
VT V. E V.O2 V . CO2 V . E V . O2 V.E V . CO2 V . CO2 V . O2 VT V.E V . O2 V.CO2 V . E V . O2 V . E V . CO2 V.O2 V . CO2 V . CO2 V . O2 V . E V . O2 V . E V . CO2
consumption rate ( ), CO2 production rate ( ), ventilatory quotient ( / ),
ventilatory equivalent for O2 ( / ), and CO2 ( / ) recorded under normoxia
after an i.p. injection of DMSO or Allopregnanolone in 12-day-old female rats that received a daily gavage of water (Control) or caffeine from the postnatal days 3-12. . 69
Table 7. Effects of ip progesterone injection on the ventilatory parameters and apnea time in
male and female 12-day old rats that have been treated with caffeine. ... 81
V.O2 V . CO2 V . CO2 V . O2 V.E V . O2 V . E V . CO2
List of abbreviations
AOP Apnea of prematurity BW Body weightCB Carotid body
CNS Central nervous system CO2 Carbon dioxide
CPAP Continuous positive airway pressure
DMSO Dimethly sulfoxide FR Respiratory frequency GABA Gamma aminobutyric acid HVR Hypoxic ventilatory response i.p Intraperitoneal
NPG Nodose-petrosal ganglion
nPR Nuclear progesterone receptor
NTS Nucleus tractus solitarius
mPR Membrane progesterone receptor
mRNA Messenger Ribonucleic acid
O2 Oxygen
P Postnatal
PAQR AdipoQ receptor family PBC PreBötzinger nucleus PROG Progesterone
"You can't connect the dots looking forward; you can only connect them looking backward. So you have to trust that the dots will somehow connect in your future. You have to trust in something - your gut, destiny, life, karma, whatever. This approach has never let me down, and it has made all the difference in my life."
Acknowledgements
Firstly, I would like to express my sincere gratitude to my supervisor Dre. Aida Bairam for giving me the opportunity to work in their lab and for her continuous support, patience, motivation, and immense knowledge. I am truly thankful to my co-supervisor Dr. Vincent Joseph, without his precious support and encouragement it would not be possible to finish my studies. Both my supervisor’s guidance helped me in all the time of research and writing of this thesis. I could not have imagined having a better advisors and mentors.
Besides my supervisors, I would like to thank the rest of our research department Dr. Richard Kinkead, Dr. Jorge Soliz for their immense support in learning of scientific techniques and maintaining motivating atmosphere in the laboratory.
I would also like to thank research assistants François Marcouiller, Roumiana Gulmotova, Stéphanie Fournier, Rose Tam and animal care technician Mélanie Pelletier who helped me during all my years of research. I thank my fellow labmates Orlane Rossignol, Hayet Kouchi, Ryma Boukari, Luana Tonerio, Cécile Baldy, Sofien Laoufa, Alexandra Jochmans-lemoine, Pablo Iturri Soliz, Alexandra Ribon, Jean Phillipe Rousseau for the stimulating discussions, and for all the fun we have had in the lab. Thanks to all the past and present members of the laboratory for being helpful in some or the other way.
I am truly thankful to my friends Nupur, Nikunj, Manasa, Sidharth, Manju, Prasanna for always being supportive in my good and bad times. All these years in Quebec we got to explore this city and did lot of activities, parties together and I will forever cherish those memories.
I am forever indebtedly thankful to my parents Dad Lakshman Rao and Mom Madhavi for always being there for me, especially my mom without her support and encouragement I
would have not been in this position. She is the inspiration of my life. My heartfelt thanks to my family – brothers Kiran, Deepak, Bunny and sister Nalini, sister-in law Neha, nephew Laksh and our newborn niece Aksha for their immense love and pamper. This journey would not have been possible if not for them, and I dedicate this milestone to my family. An recent extended family, special thanks to my in-laws for their constant love and support.
Last but not the least, words cannot express how grateful I am to my beloved husband Chandu for all of the sacrifices he made on my behalf. He was always by my side in my up and downs. I am lucky to have you in my Life.
Avant-propos
Les résultats scientifiques présentés dans cette thèse sont le fruit d'un travail d'étudiant diplômé au laboratoire de Dre. Aida Bairam au centre de recherche de l'Hôpital St-François D'Assise du CHUQ (Université Laval) puis au centre de recherche de l'Institut de Cardiologie et de Pneumologie de Québec (Université Laval). Ces études ont été financées par une subvention de l'Institut canadien de recherche en santé (IRSC) à Dre. Aida Bairam. J'ai contribué en tant que premier auteur à deux manuscrits.
La première étude qui constitue le chapitre 2 et présentée à l'appui de cette thèse a été réalisée dans le laboratoire de mon directeur Dre. Aida Bairam et co-directeur de recherche Dr Vincent Joseph au centre de recherche de l'Hôpital Saint François d'Assise affilié au Centre Hospitalier Universitaire de Québec (Université Laval). Ces études ont été réalisées avec le soutien financier des Instituts de recherche en santé du Canada (IRSC) accordé à mon directeur de recherche. Je suis le premier auteur, j'ai participé aux expériences, analyses et interprétations des données générées, j'ai également participé aux corrections du manuscrit.
Inhibitory respiratory responses to progesterone and allopregnanolone in newborn rats chronically treated with caffeine.
NagaPraveena Uppari, Vincent Joseph, Aida Bairam Publication: The Journal of Physiology, Janvier 2016
J Physiol. 2016 Jan 15;594(2):373-89. doi: 10.1113/JP270914. Epub 2015 Dec 14
La deuxième étude qui constitue le chapitre 2 et présentée à l'appui de cette thèse a été réalisée dans le laboratoire de mon directeur Dre. Aida Bairam et co-directeur de recherche Dr Vincent Joseph au centre de recherche de l'Hôpital Saint François d'Assise affilié au Centre Hospitalier Universitaire de Québec (Université Laval). Ces études ont été réalisées avec le soutien financier des Instituts de recherche en santé du Canada (IRSC) accordé à mon directeur de recherche. Je suis le premier auteur, j'ai participé aux expériences, analyses et interprétations des données générées, j'ai également participé aux corrections du manuscrit.
Respiratory responses to progesterone and allopregnanolone following chronic caffeine treatment in newborn female rats
NagaPraveena Uppari, Vincent Joseph, Aida Bairam
Publication: Respiratory Physiology and Neurobiology, June 2017
Respir Physiol Neurobiol. 2017 Jun;240:32-40. doi: 10.1016/j.resp.2017.01.017. Epub 2017 Feb 20.
Preface
The scientific findings presented in this thesis is the product of work as a graduate student in the laboratory of Dre. Aida Bairam at the research center of the St-François D'Assise Hospital of the CHUQ (Laval University) then at the research center of the Institut de Cardiologie et de Pneumologie de Québec (Laval University). These studies were supported by a grant from the Canadian Institute for Health Research (CIHR) to Dre. Aida Bairam. I have contributed as the first author to two manuscripts.
The first study which constitutes chapter 2 and presented in support of this thesis was carried out in the laboratory of my director Dre. Aida Bairam and co-director Dr. Vincent Joseph at Saint François d'Assise Hospital research center affiliated with the Center Hospitalier Universitaire de Québec (Université Laval). These studies were carried out with the financial support of the Canadian Institutes of Health Research (CIHR) granted to my research director. I am the first author, I participated in the experiments, analysis and interpretation of the data generated, I also participated in the corrections of the manuscript.
Inhibitory respiratory responses to progesterone and allopregnanolone in newborn rats chronically treated with caffeine.
NagaPraveena Uppari, Vincent Joseph, Aida Bairam Published: The Journal of Physiology, January 2016
J Physiol. 2016 Jan 15;594(2):373-89. doi: 10.1113/JP270914. Epub 2015 Dec 14
The second study which constitutes chapter 2 and presented in support of this thesis was carried out in the laboratory of my director Dre. Aida Bairam and co-director Dr. Vincent Joseph at Saint François d'Assise Hospital research center affiliated with the Center Hospitalier Universitaire de Québec (Université Laval). These studies were carried out with the financial support of the Canadian Institutes of Health Research (CIHR) granted to my research director. I am the first author, I participated in the experiments, analysis and interpretation of the data generated, I also participated in the corrections of the manuscript.
Respiratory responses to progesterone and allopregnanolone following chronic caffeine treatment in newborn female rats
NagaPraveena Uppari, Vincent Joseph, Aida Bairam
Published: Respiratory Physiology and Neurobiology, June 2017
Respir Physiol Neurobiol. 2017 Jun;240:32-40. doi: 10.1016/j.resp.2017.01.017. Epub 2017 Feb 20.
Introduction
As mentioned by recent nobel prize winner Dr. Gregg L Semenza “Respiration is the
key function vital for the survival of all the living cells and for the organism” (Semenza G.L.,
2007). Preterm infants born less than 34 weeks gestation encounter problems with temperature instability, hypoglycaemia, respiratory distress, feeding issues and apnea (Marchal F et al., 1987). Brainstem development and neural control of respiration are less mature comparatively to full-term infants (Darnall R.A et al., 2006).
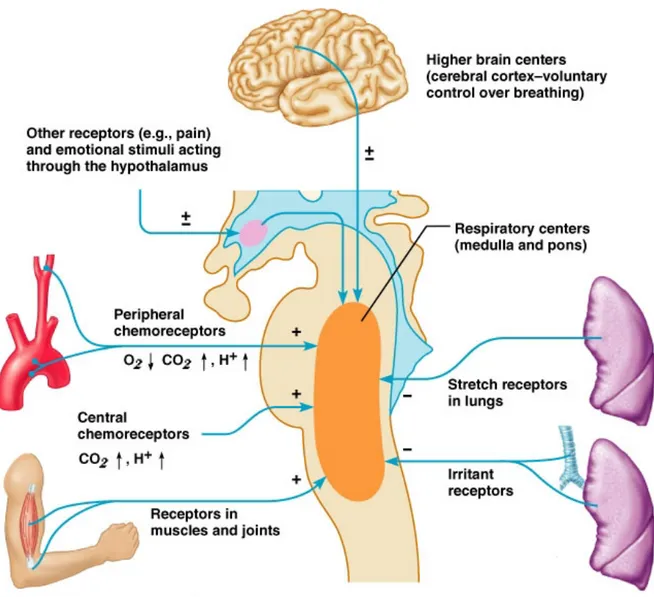
A. Structures involved in control of breathing
The system that generates the respiratory rhythm is located in the preBötzinger complex (brainstem) (Gray P.A et al., 1999; 2001). This respiratory nucleus sends motor commands towards the spinal cord motorneurons to activate the phrenic nerve and respiratory muscles (such as diaphragm and upper airways). It also receives afferents from higher cortical areas, thalamus, muscle proprioeceptors, peripheral and central chemoreceptors. This system produces an alternation between inspiration and expiration through activation of the upper airways, abdominal and intercostal muscles (Feldman and Del Negro., 2006). The chemosensory organs that monitor blood oxygen (O2) and
carbondioxide (CO2) levels are located primarily in the carotid bodies (CB) and in the
brainstem. The CB’s are located at the bifurcation of the carotid arteries and generate signals that relate mostly to O2 pressure in arterial blood. Central chemoreceptors are localized in
the ventral medullary surface and are sensitive to changes in CO2 and pH (Mellen N.M et
Figure 1. Elaboration of the structures involved in the control of breathing
Figure adapted from Feldman and McCrimmon, Fundamentals of Neurosciences, 2nd edition,
Academic Press.
In newborn mammals, immature control of breathing is characterized by low
chemoreflex responses to natural stimuli (hypoxic ventilatory response, HVR) and high frequency of breathing instability (apnea).
A.1 Hypoxic ventilatory response (HVR) in newborn
Since 1952, it is very well known that in newborn infants and animals the HVR is weak and show a biphasic pattern (Neilson M and Smith H., 1952; Cohen G et al., 1997; Nock M.L et al., 2004). The initial increase of ventilation results from the stimulation of peripheral chemoreceptors. The second phase of decline starts after 2 to 3 min of exposure to hypoxia and originates mainly in the central nervous system where inhibitory mechanisms override the initial peripheral chemoreceptor stimulation (Martin R.J et al., 1998). For instance, this biphasic pattern of HVR was predominant in rats less than 12 days old (Liu Q, T.F. Lowry, and M.T., 2006; Liu, Q. and M.T. Wong-Riley., 2006; Niane, L.M. and A. Bairam., 2012) indicating that, the end of second postnatal week is a critical period of development for brainstem respiratory function.
A.2 Implication of neurotransmitters in control of breathing
The expression levels of several neurotransmitters and their receptors are expected to undergo postnatal changes as synapses gets established, pruned and matured (Niane L.M et al., 2011). Important neurotransmitter systems, including dopamine, glutamate, adenosine, Gamma-aminobutyric acid (GABA), serotonin and acetylcholine undergo rapid and varying changes during late gestation resulting in continuing instability in the control systems involved in breathing. The main inhibitory neurotransmitters that are thought to be involved in the expression of the biphasic HVR and respiratory instability in developing newborn mammals are GABA and adenosine (Liu, Q. and M.T. Wong-Riley., 2006; Bairam A et al., 2013).
A.2.1 Gamma-aminobutyric acid (GABA)
GABA (Gamma-aminobutyric acid) is a major inhibitory neurotransmitter in the central nervous system (CNS). In the rat hippocampus between postnatal days 0 to 8 (P0-P8), approximately 85-93% of CA3 neurons and 25% at P11-P12 exhibit spontaneous giant depolarizing potentials mediated by GABA. These events are no longer present after P12 indicating that GABA transmission has switched to hyperpolarization (Liu, Q. and M.T. Wong-Riley., 2005). Studies conducted in rat brain stem respiratory nuclei (P0-P21) by Liu and his group showed the possible switches in GABAA subunits (α1, α2 and α3). In the
preBötzinger nucleus (PBC), and ventrolateral sub nucleus of the nucleus tractus solitarius (NTSVL), expression of GABA varied at different postnatal ages and the switch from α3 to α1 subunit dominance occurred around P12 (Liu, Q., T.F. Lowry, and M.T. Wong-Riley., 2006; Liu, Q. and M.T. Wong-Riley., 2006; Liu, Q. and M.T. Wong-Riley., 2005). Also data from ex-vivo studies using the brainstem–spinal cord preparation showed a transition from excitatory to inhibitory action of allopregnanolone as early as embryonic day 19 (Ren J and Greer J.J., 2006). This demonstrates the age-dependent changes of GABAA
receptor-mediated activities on respiratory rhythmogenesis during the perinatal period.
A.2.2 Adenosine
Adenosine, a metabolite of cellular ATP, is released from cells in response to inadequate tissue oxygenation. Adenosine mediates its cellular effects via specific membrane receptors. To date, four Ade receptor subtypes have been identified (A1, A2A, A2B, and A3). The A1 and A3 receptors are coupled to the inhibitory G protein (Gi) and mediate inhibition of adenylate cyclase. The A2 receptor family, on the other hand, is coupled to the stimulatory G protein (Gs) and activates adenylate cyclase. Adenosine is a main inhibitory transmitter in the CNS including the brain stem respiratory nuclei (Gauda E.B et al., 2006). There is also evidence that Adenosine is produced in newborn rats during hypoxia in CB (Chen G. and A.N. Van den Pol., 1997). During the neonatal period, A1 receptor mRNA is not present in CB but is found in the nodose-petrosal ganglion (NPG) which contains the chemo-afferent cell bodies, while A2 receptor mRNA expression increases during postnatal development in the NPG (Gauda E.B et al., 2006). In adult rats that have been treated with caffeine as newborn (between postnatal days 3 and 12), there was a 37% increase in excitatory A2a receptor expression in the CB and 13% decrease in the inhibitory A1 receptor in the nucleus tractus solitarius (NTS) (Bairam A et al., 2009).
B. Breathing Instability
Apnea of prematurity (AOP) is one of the most common problems encountered in premature infants (Darnall R.A et al., 2006; Zhao J et al., 2011). As stated by Jing Zhao and team “The most widely used definition of apnea of prematurity (AOP) specifies a pause in
2011). The occurrence of AOP with gestational age are 85% of infants born before 34 weeks: 7% at 34–35 weeks, 15% at 32–34 weeks, 54% at 30–31 weeks, and almost all infants born earlier than 30 weeks or born with birth weight less than 1000g (Martin R.J et al., 2004).
B.1 Types of Apnea
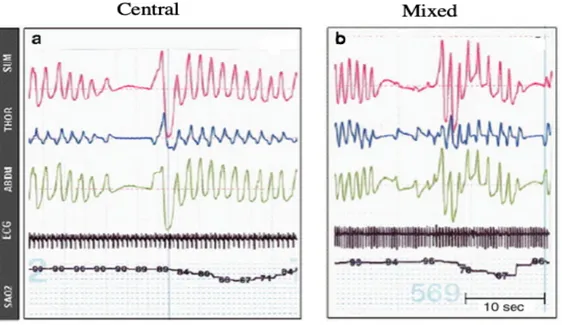
The three main types of apnea are central, obstructive, and mixed. Central apnea is characterized by total cessation of inspiratory efforts with no evidence of obstruction. Obstructive apnea is described by abnormal collapse of the pharyngeal airway and therefore no air exchange with the lungs. Frequently with obstructive apnea, there are vigorous inspiratory efforts causing arousal. Mixed apnea refers to an episode with combinations of both central and obstructive apnea and is probably the most common type of apnea (Martin R.J et., 2004). The percentage occurrences for types of apnea are central apnea – 40%, obstructive apnea – 10% and mixed apnea – 50% (Martin R.J et., 2011).
Figure 2. Two representative examples of desaturation, secondary to apnea or respiratory
pauses, in preterm infants monitored via respiratory inductance plethysmography
(a) represents an isolated central apnea, (b) demonstrates a mixed apnea with obstructed inspiratory efforts preceding resumption of breathing
SUM : ABD+THOR, THOR : Thoracic movements, ABD : Abdominal movements, ECG : Electroencephalogram, SAO2 : Arterial oxygen saturation (Martin R.J et al., 2011).
B.2 Apnea Treatment
There are different modalities such as by prone position, kangaroo mother care, sensory stimulation (tactile and olfactory stimulation) to reduce the frequency of apnea in preterm neonates. Other nonpharmacological approaches, mechanical ventilation include nasal (continuous positive airway pressure, CPAP) or tracheal. These methods stimulate inspiration, reduce obstruction or facilitate air entry. Among treatments available, mechanical ventilatory support and pharmacological support (methylxanthines) had proven to be more effective (Zhao J et al., 2011).
B.2.1 Xanthine Therapy
Methylxanthines have been the keystone of pharmacologic support to treat apnea. Methylxanthine compounds such as caffeine, theophylline, and aminophylline are powerful CNS stimulants and likely reduce apnea by multiple physiological (increasing ventilation, enhancing the respiratory response to O2 and CO2, improving lung compliance) and pharmacological mechanisms (antagonism of adenosine A1 and A2a receptors in the CNS) (Marchal, F et al., 1987; Hascoet, J.M et al., 2000; Bairam A et al., 2009). Although all methylxanthines compounds are effective for the treatment of apnea, caffeine was considered to be safer since it had a wider therapeutic range than theophylline and longer plasma half-life (100 hours compared to 30 hours for theophylline) (Mueni E et al., 2009; Carnielli, V.P et al., 2000; Bairam A et al., 1987; De Carolis MP et al.,1991). Caffeine reduced the incidences of cerebral palsy and cognitive delay and improved survival without neurodevelopmental disability at a corrected age of 18 to 21 months, providing strong evidence that the overall benefits of caffeine therapy can outweigh any potential risks up to 2 years after very preterm birth (Gray P.H et al., 2011; Schmidt B et al., 2007).
B.2.2 Failure of Methylxanthines
Caffeine’s efficiency to treat apnea is well known and is currently the main therapy worldwide. In addition to reducing the cost of medical care, early onset of caffeine treatment is associated with a greater reduction in the time during which breathing must be supported by artificial ventilation. Nevertheless, even when therapeutic plasma levels of caffeine are achieved a substantial percentage of pathological apneas persist. Such persistence often
necessitates mechanical ventilation, particularly in very preterm neonates (Martin R.J et al., 2004; Hascoet J.M et al., 2000; Poets C.F., 2010, 2010; Erenberg A et al., 2000).
To decrease morbidity and usage of mechanical support with caffeine treatment it has been suggested that progesterone, which is well-known as a respiratory stimulant in adults, could be an alternative therapy for the treatment of apnea in premature infants (Finer N.N et al., 2006).
C. Progesterone
Steroid circulating hormones including estrogen, progesterone, androgen, glucocorticoid and mineralocorticoid are mainly synthesized in adrenal glands, the gonads and the feto-placental unit.They play a pivotal role in the differentiation of the CNS acting via their cognate receptors (figure 3). In the central and peripheral nervous system, steroid hormones are synthesized de novo from cholesterol and are therefore recognized as “neurosteroids” (figure 4) (Brinton R.D et al., 2008; Schumacher M et al., 2014). Furthermore, allopregnanolone a natural metabolite of progesterone, had no affinity for the intracellular progesterone receptor (PR), but it is a potent allosteric modulator of GABAA receptors (Hosie A et al., 2006).
C.1 Progesterone receptors
Two major PR forms are known, namely nuclear and membrane receptors.
C.1.1 Nuclear Progesterone Receptors (nPR)
Nuclear PR are a class of proteins belonging to the larger superfamily of steroid receptors that act as powerful transcription factors. The hypothalamus was the first region where PR were localized in the CNS and it was later found that expression of these receptors are not limited to hypothalamus, but they are widely distributed throughout the brain regions of adult and newborn rats, in the hippocampus, cerebral cortex, cerebellum and subcortical regions (Brinton R.D., et al., 2008; Schumacher M et al., 2014). nPR has two major isoforms PR-A and PR-B. Both of the classical isoforms are expressed in hippocampus, hypothalamus, frontal cortex, amygdala, cerebellum and thalamus. nPR are also localized in brain stem nuclei such as the NTS and locus coeruleus (Brinton R.D et al., 2008, Haywood S.A et al., 1999, Helena C et al., 2009). In the peripheral chemoreceptors of adults, newborn
andfetal rats there is a dense immunostaining for nPR, localized in clusters of chemosentitive cells. nPR mRNA was also found in CB of adult rats (Joseph V et al., 2006).
C.1.2 Membrane Progesterone Receptors (mPR)
Membrane PR belong to the progestin and adipoQ receptor family (PAQR) that mediate cell surface-initiated progesterone actions in the reproductive system involving action of intracellular signaling pathways. The mPRs were first characterized in fish ovaries (Zhu Y et al., 2003) and five isoforms (mPRα, mPRβ mPRγ mPRδ and mPRε) were identified in humans and vertebrates. In adult, some of these isoforms are present in the spinal cord or in the brain (Zhu Y et al., 2003; Intlekofer K.A. and S.L. Petersen., 2011; Labombarda, F., et al., 2010; Pang Y J. Dong, and P. Thomas., 2013). In newborn mPR mRNA was found in cerebellum (Sakamoto H et al., 2008).
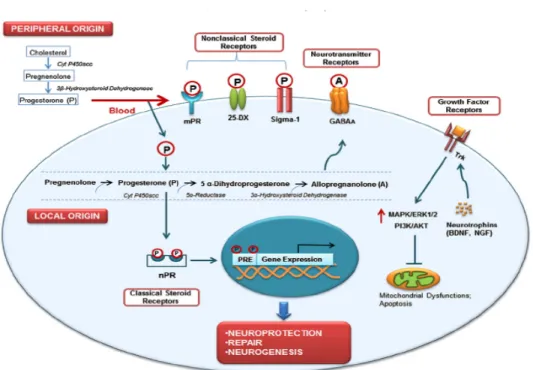
Figure 3. Progesterone metabolism and receptor actions in brain
Progesterone is synthesized not only in the ovaries, adrenal glands and placenta (peripheral origin), but also in both neurons and glia (central origin). Progesterone of peripheral origin reaches the central nervous system through the bloodstream and binds to several non-classical (25-Dx, Sigma-1, mPR) and non-classical (nPR) progesterone receptors to regulate intracellular signaling pathways and, transcription of target genes. Metabolism of progesterone results in generation of its active metabolite allopregnanolone, which then binds to neurotransmitter receptors (GABAA), acting as an allosteric modulator. By binding
to its nuclear receptor, progesterone regulates the expression level of several target genes which are involved in neuroprotection, neuronal repair and neurogenesis. It specifically upregulates the expression of growth factors (BDNF, NGF, etc.), which then bind to their specific Trk receptors. Activated Trk receptors further activate PI3K/AKT and MAPK signaling to enhance neuroprotection and repair by upregulating anti-apoptotic proteins (such as BCl-2) and inhibiting mitochondrial dysfunction-mediated apoptosis. 25-DX: membrane receptor, mPR: membrane progesterone receptor; nPR: nuclear progesterone receptor; PRE: progesterone response element; BDNF: brain-derived neurotrophic factor; NGF: nerve growth factor; TrK: tropomysin related kinase; MAPK: mitogen activated protein kinase; Erk1/2: extracellular signal-regulated kinase1/2; PI3K/AKT: phosphoinositide 3-kinase/protein kinase B (Donald G. Stein., 2012).
Figure 4. Biosynthetic Pathway of Progestogens
The cholesterol molecule shows the numbering of the 27 carbons of the sterane nucleus, its functional groups and the 6 carbon side-chain (in red) which is characteristic of sterols. Cholesterol is converted to pregnenolone inside the mitochondria by the side chain cleavage cytochrome P450scc. Pregnenolone is then converted to progesterone by the 3β -hydroxysteroid dehydrogenases (3β-HSD). The conversion of progesterone to 5α-dihydroprogesterone is catalyzed by one of the two 5α-reductase isoforms. Both progesterone and 5α-DHP bind to the classical intracellular progesterone receptors (PR), which regulate gene transcription within the cell nucleus or interact with kinases and components of intracellular signaling pathways within extra-nuclear compartments. 5α-DHP can be metabolized to allopregnanolone (3α,5α-tetrahydroprogesterone) by aldo-keto reductases (ARK; 3α-HSD = 3α-hydroxysteroid dehydrogenase). Because of its hydroxyl group at C3, allopregnanolone has no affinity for the intracellular PR, but is a potent allosteric modulator of GABAA receptors (Schumacher M et al., 2014).
C.2 Progesterone as a respiratory stimulant C.2.1 In adults
Progesterone is a well-known respiratory stimulant in mammals including human, acting peripherally (CB) and centrally (through brainstem and hypothalamic structures) to increase ventilation in response to hypoxia (HVR) (Behan M. and J.M Wenninger., 2008). Administration of progesterone: (a) reduces CO2 retention in patients suffering from chronic obstructive pulmonary disease (Dolly F.R and Block A.J., 1983; Skatrud J.B and Dempsey J.A., 1983; Al-Damluji S., 1986); (b) induces hyperventilation (Slatkovska L et al., 2006); (c) reduces upper airway resistance during sleep (Driver H.S et al., 2005); and in healthy men, medroxyprogesterone increased minute ventilation as well as the hypoxic and hypercapnic ventilatory responses (Skatrud J.B et al., 1978). Postmenopausal women have a greater apnea-hypopnea index than pre-menopausal women (Young T et al., 2003). In postmenopausal women, combined hormone therapy (estrogen and progesterone) reduced sleep apnea (Shahar E et al., 2003).
C.2.2 In newborn
Newborn mammals are exposed to high levels of progesterone in utero (synthesized by the placenta, the adrenals or by neural networks) and there is a significant decrease of progesterone levels after birth (24–48 hours) (Runnebaum B et al., 1975; Trotter A et al., 1999). In humans and rats, progesterone is also produced by the adrenals in both females and males (Romeo R.D et al., 2005; Romeo R.D., 2010; Brunton P.J and Russell J.A., 2011).
In ten-day-old rats, chronic exposure to progesterone through the milk of lactating mothers increases the HVR and decreases the frequency of apnea during normoxia and hypoxia (Lefter R et al., 2007). Acute progesterone administration in a dose range between 0-8 mg/kg did not influence the normoxic ventilation or metabolic rate in both 4 and 12-day-old rats. However, progesterone with dosage of 4 mg/kg enhanced HVR in 4-day old but not in 12-day-old rats (Hichri O et al., 2012). At 4 mg/kg progesterone increases HVR in 1-, 4- and 7- days old but not in 12-day old rats showing age dependent effect. Interestingly, there was a decrease in apnea frequency only in 1-day old rats suggesting that different mechanisms are
involved in mediating the effects of progesterone on HVR and apnea frequency (Bairam A et al., 2013).
D. Sex-specific differences in respiratory studies D.1 In Neonates
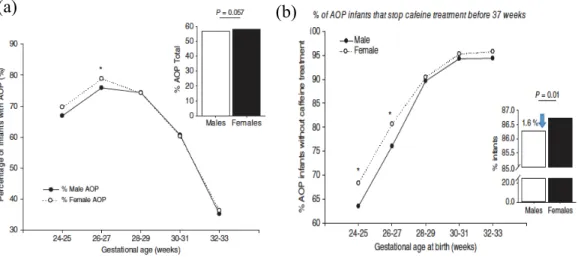
The human females appear to have a greater resistance to suffocation and asphyxiation than males in case of hypoxic/ischemic injuries at birth (Mage D.T and Donner E.M., 2004), respiratory distress syndrome (Condo V et al., 2017). There is also higher mortality rate from Sudden Infant Death Syndrome (50%) in males compared to females (Kinney H.C and Thach B.T., 2009; Mage D.T and Donner E.M., 2004; 2006; 2009 and 2014). Addressing AOP, Bairam and colleagues published data showing that the percentage of infants with apneas was higher in females born at <27 weeks of gestation. However, males were treated with caffeine longer compared to females (Figure 4) (Bairam A et al., 2018).
Figure 5. Percentage of infants diagnosed with AOP in different gestational age group
(a) The percentage of infants with apnea is higher in females than males before 27 weeks of gestation (b) percentage of AOP infants for which caffeine treatment was discontinued between 24 and 33 weeks of gestational age at birth, or before 37 weeks of gestation (Bairam A et al., 2018).
D.2 In newborn animals
Studies in newborn mice, at postnatal day 11, suggest that females are protected against the effects of hypoxic ischemia since there is a 43% reduction of hippocampal volume in males whereas 36% was observed in female mice (Hill C.A and Fitch R.H., 2012), and disrupted myelination was noticed only in males (Mayoral S.R et al., 2009) furthermore, male rats had more impairments in associative learning, spatial memory and long-term social memory compared to female rats (Lan W.C et al., 2011). Physiological studies show that males subjected to gestational stress have more apneas than females (Fournier S et al., 2013). Recent studies showed that caffeine had sex specific effects in newborn rats which varied with the mode of administration ie., acute and chronic, during normoxia and hypoxia. Interestingly, the frequency of apneas were lower in female rats compared to male rats with or without caffeine (Kouchi H et al., 2017).
Caffeine and progesterone are both able to reduce the frequency of apneas, acting through different pathways. In a clinical setting, caffeine is the first line of intervention to decrease the frequency of apnea in preterm neonates. If not efficient, complementary approaches can be used. Because potential interactions between drugs are a concern, this should be investigated by preclinical studies.
E. Hypothesis
We hypothesized that progesterone administration following a chronic treatment with caffeine further stimulates breathing and reduces apnea.
E.1 Objectives
Our objectives are to determine the effect of a single administration (intraperitoneal (i.p) injection) of saline or progesterone (4 mg/kg) in male and female rats daily gavaged with caffeine from postnatal days 3 to 12 (15 mg/kg/day) on:
1) Basal ventilation (normoxia)
2) In response to hypoxia (FiO2 = 12%; 20mins)
3) Apnea frequency during normoxia and hypoxia Two studies were hence performed
In the first study, we used male rats to asses :
- Age dependent effects (4 and 12 days old rats)
- Dose dependent effects of bicuculline – a GABAA receptor antagonist -
on ventilation and apnea
- Effect of allopregnanolone on ventilation and apnea
- Effect of finasteride (blocking the conversion of progesterone to allopregnanolone)
Materials and Methods
F AnimalsAll experiments complied with the guidelines of the Canadian Council on Animal Care and were approved by the animal care committee of Université Laval. Sprague-Dawley rats were used, supplied with food and water ad libitum, and maintained in standard animal care conditions (21°C, 12h dark–light cycle: lights on at 07:00 and off at 19:00). Twenty-four hours following birth, litters were culled to 12 pups, if possible with similar number of males and females. All pups with their mother were euthanized at the end of experiments by an overdose of anaesthetic drugs (ketamine 200 mg/kg - xylazine 20 mg/kg, corresponding to 2 times the normal dose for anaesthesia) injected i.p (0.2 ml/10g). The numbers of rats used in 1st and 2nd study are presented in table 1 below.
4 days old 12 days old Total
Male Female Male Female
1st Study 68 - 174 - 242
2nd Study - - 121 121
Table 1. Number of animals used in each study F.1 Neonatal gavage with water or caffeine
All gavages were given between 8:00 and 9:00 am daily. Pups from each litter were randomly assigned to receive a daily oral gavage of water (control) or caffeine (15 mg/kg; Sandoz, Boucherville, QC, Canada, 0.05 ml/10 g body weight) between the postnatal days 3 to 12. A polyethylene tubing (PE10) was gently inserted into the esophagus up to the stomach, and the appropriate volume was injected slowly over a 30 seconds period (Montandon et al., 2006). This dose of caffeine generated plasma caffeine levels in the range
of 7-17 mg/l (Montandon et al., 2006), which matches the plasma caffeine range in premature infants treated for apnea (Bairam et al., 1987; Zhao et al., 2011; Spitzer, 2012). In control rats, the plasma caffeine level is below the detection threshold (Montandon et al., 2006).
F.1.1 Drug preparation and administration
In the 1st study the effects of progesterone were assessed at the age of 4 and 12 days
old in control and caffeine-treated male rats. Whereas in the 2nd study 12 days old female
rats were used. The following drugs were used in the 1st and 2nd studies:
F.1.2 Progesterone
On the day of experiment, the rats received an i.p. injection of saline or progesterone (Sigma Aldrich, CAT# P0130) at 4 mg/kg based on dose-response study (Hichri et al., 2012).
F.1.3 Bicuculline
The effect of chronic caffeine administration on GABAA function were assessed
using bicuculline (GABAA antagonist - Sigma Aldrich, CAT# 14340) and in these
experiments, 12 days old rats received a random i.p. injection (0.1 ml/30 g body weight) of either vehicle, or 0, 1, 2, or 3 mg/kg of bicuculline dissolved in water with 2% dimethyl sulfoxide (DMSO, Sigma Aldrich, CAT# DMS666) and 10% 2-hydroxypropyl-β-cyclodextrin (Sigma Aldrich, CAT# H107) (Ren, J. and J.J. Greer, 2006).
F.1.4 Allopregnanolone
The effects of allopregnanolone, the neuroactive metabolite of progesterone and allosteric modulator of GABAA, on ventilatory inhibition in rats treated with caffeine with
the dose of 10mg/kg were studied. (Sigma Aldrich, CAT# P8887) (Ren, J. and J.J. Greer, 2006).
F.1.5 Finasteride:
The contribution of the endogenous conversion of progesterone to allopregnanolone was assessed by finasteride (10 mg/kg dissolved in 2% DMSO and 10% cyclodextrin; Sigma Aldrich, CAT# F1293), a 5α-reductase inhibitor that blocks the conversion of progesterone to allopregnanolone.
All the drugs were freshly prepared on the day of experiment and respiratory recordings with whole body plethysmography were started 30 minutes post injection.
F.2 Measurement of Respiratory and Metabolic Parameters
The respiratory frequency (FR – expressed as breaths/min) and the tidal volume (VT - mL) are recorded by whole-body plethysmography. They make it possible to calculate the minute ventilation (VE – mL/min) (VE = FR * VT). For the recording of respiration, the animal is introduced into a plethysmography chamber (20 cm3) where it is free of any restrain
and sedation, thus reducing respectively the effects of stress and pharmacological substances on the respiratory system of the animal. The principle of the recording underlies that the air inspired by the animal is warmed and moistened by its passage in the airways and the alveoli. This creates a pressure difference inside the plethysmography chamber that will be measured by a differential pressure sensor (Drorbauh J.E and Fenn W.O., 1955). The inverse transformation occurs when the air is exhaled creating a pressure deviation opposite to that of inspiration. In this project, as the system will be open, and not closed, a difference in flow rather than pressure will be caused by breathing. By integrating the flow rate value, it is possible to calculate the equivalent pressure change. The pressure values of the chamber of the animal are compared to the values of a reference chamber in order to make the measuring system independent of sudden pressure changes occurring at the recording site (Bartlett D and Tenney S.M., 1970). The recording is used to determine the frequency and type of apnea. As shown in figure 5, the workstation consists of a flowmeter (model 4140, TSI, Shoreview, MN) to measure the airflow into the chamber, a water vapor pressure analyzer (pH2O) (RH
300, Sand Systems International, Las Vegas, NV, U.SA) and a differential pressure sensor (Emka Technologies, Paris, France). These measurements are necessary for calculating the tidal volume, which must also be divided by the mass of the animal in order to avoid differences in inter- and intra-individual growth. The pump (Qubit Systems, Kingston, ON,
Canada) at the end of the circuit will sample outgoing air. Finally, incoming and outgoing O2 and CO2 analyzers (AEI, technologies, Pittsburgh, PA, USA) collect the data necessary
for calculations of metabolic rate.
Figure 6. Schematic representation of whole-body plethysmography.
(Seaborn T et al., 2013) Mathematical formula allowing the calculation of the tidal volume. Values to calculate the tidal volume
VT (BTPS) = PT/PK x VK x [TR (PB-PC)/ (TR [PB-PC] – TC [PB-PR])]
The symbols in the equation are defined as follows:
VT = tidal volume; BTPS = volume expressed in terms of vapor pressure of water and corrected for body temperature of the animal/organism and is standardised at 37°C; PT = amplitude of the respiration pressure change; PK = amplitude of the calibration of pressure variation; VK = calibration volume (ml); TR = internal temperature of the animal (K); TC =
saturated vapor pressure in the plethysmography chamber (mmHg); PR = saturated vapor pressure at the internal temperature of the animal (mmHg).
Oxygen consumption (VO2) and CO2 production (VCO2) rates were calculated using the
following equations (Lighton, 2008). Metabolism values are adjusted for animal mass to compensate for intra- and inter-individual growth differences.
O2 consumption = Flow x [(O2in –O2out) – O2out x (CO2 out – CO2in)]
(1–O2out)
CO2 production = Flow x [(CO2out –CO2in)-CO2out x (O2 in – O2out)]
(1–CO2out)
F.2.1 Plethysmography Recording
The newborn is first weighed and then placed in the plethysmography chamber for 10 minutes for an initial adaptation period. Its oral temperature is then taken with a thermometer (Thermocouple Thermometer, Barrington, IL, USA) and the newborn is reintroduced into the chamber for another 10 minutes for a second adaptation period. Basal breathing is recorded for 10 minutes (normoxia, O2 = 21%). then, the animal is exposed to
hypoxia (FiO2 = 12 ± 0.2 %), acheived by mixing ambient air and nitrogen for 20 minutes.
The desired concentration of O2 in the chamber is reached in ≈ 2 minutes. Each hypoxia lasts
All signals are recorded on the computer (IOX, Emka Technologies, Paris, France). The average value of the various parameters is reported minute by minute. The subsampling pump is switched on for a few minutes at the end of the periods of normoxia and hypoxia to measure the O2 and CO2 values coming out of the chamber. These values allow the
F.2.2 Analysis of ventilatory parameters and respiratory irregularities
For recordings under normoxia, the periods when the animal is agitated are rejected. The average of the various parameters (respiratory rate, tidal volume and minute ventilation) is then calculated with the remaining periods. The respiratory rate (FR) is expressed as breaths / min, the tidal volume (VT) is expressed as ml / 100 g and the minute ventilation (VE = FR * VT) is expressed as ml / min / 100 g. Oxygen consumption and CO2 production are
expressed as ml / min / 100 g. In hypoxia, HVR is expressed as % change from normoxia. The average minute-by-minute respiratory parameters are used to illustrate temporally the changes in respiratory rate, tidal volume and minute ventilation for hypoxia. The results under hypoxia conditions are then separated in the initial phase and in the stable state. For the initial phase, the recording of the second minute value is used as the peak value of the respiratory parameters. The steady state is calculated by averaging the last five minutes of recording for each period of hypoxia.
Apneas are analyzed when the breathing pattern is stable and regular. Two types of apneas were assessed, spontaneous apnea are defined as respiratory pauses equivalent to two regular breathing cycles. Post-sigh apnea preceded by a sigh, defined as a breath with an amplitude of at least twice the resting tidal volume. Under normoxia, apneas are analyzed during 10 minutes of selected regular breathing. In hypoxia, apneas are analyzed during the steady state, ie during the last 10 minutes of recording of each hypoxia. The total number of apnea is calculated by summing the number of spontaneous and post-sighs apneas. Apnea duration was reported in seconds. Total apnea time (reported as seconds/10 minutes) was calculated as the total combined time spent in both spontaneous and post-sigh apneas.
F.2.3 Circulating progesterone and allopregnanolone levels
Twelve 12 day old rats were kept under room air for approximately 20 to 30 minutes after hypoxic exposure. Then they were deeply anesthetized (ketamine/xylasine, 100/15 µg/100 g) and blood samples were collected by cardiac puncture. The serum was separated and stored at -80°C. We used enzyme immunoassay kits to detect serum levels of progesterone (582601, Cayman Chemical, Michigan USA; detection limit 10 pg/ml; 50%
sensitivity at 70 pg/ml) and allopregnanolone (KO44-H1, Arbor Assays, Michigan USA; detection limit 0.24 ng/ml; sensitivity 0.13 ng/ml).
F.2.4 Statistical analysis
In the 1st study, for each age, we used a two-way ANOVA to detect significant
differences within groups (control, caffeine) and treatments (saline, progesterone). If a difference was detected by ANOVA, we used a Fisher’s LSD post-hoc test to determine where the difference occurred: within groups, within treatments, or between groups and treatments. For hypoxic ventilatory responses within each group or treatment, we used a two-way repeated-measures ANOVA with time in hypoxia as the repeated variable and with groups (control, caffeine) or treatments (saline, progesterone) as the independent variable, followed by a post-hoc analysis to determine significant effects at each time point. No comparisons between ages were performed, as the aim was to determine specific effects of progesterone at each age. Similar analyses were used for bicuculline, allopregnanolone, and progesterone plus finasteride experiments.
For the 2nd Study, we used a two-way ANOVA to first detect significant differences within groups (control, caffeine) and treatments (ex: saline vs progesterone). Then we used a Fisher’s LSD post-hoc test to determine where the difference occurred: within groups, within treatments, or between groups and treatments. For hypoxic ventilatory responses within each groups or treatments, we used two-way repeated-measures ANOVA with time in hypoxia as the repeated variable and with groups (control, caffeine) or treatments (saline, progesterone) as the independent variables, followed by a post-hoc analysis to determine significant effects at each time point.
All values are mean ± sem, and p<0.05 was considered statistically significant. All figures were made using Prism software (version 6.0, Graphpad Software, La Jolla, CA, USA).
CHAPTER I
Inhibitory respiratory responses to progesterone and
allopregnanolone in newborn rats chronically treated
with caffeine.
NagaPraveena Uppari, Vincent Joseph, Aida Bairam
Unité de recherche en périnatologie, Centre Hospitalier Universitaire de Québec, Hôpital Saint-François d’Assise, Département de Pédiatrie, Université Laval, Québec, Canada. Running title: Caffeine, progesterone, allopregnanolone, and GABA in newborn rats.
Key Points Summary
- In premature newborn recurrent apnea are systematically treated with caffeine to
prevent long-term neurocognitive disorders, but a substantial percentage of apnea persists particularly in neonates born before 28 weeks of gestation
- Progesterone has been proposed as a respiratory stimulant potentially suitable for the
treatment of newborn apnea persistent to caffeine. Accordingly we asked whether acute progesterone administration reduces apnea frequency in newborn rats treated with caffeine.
- Surprisingly our results show that in newborn rats treated with caffeine, administration
of progesterone inhibits breathing and increases apnea frequency.
- Additional experiments showed an enhanced GABAergic inhibitory drive on breathing
after caffeine treatment, and that progesterone is converted to allopregnanolone (an allosteric modulator of GABAA receptors) to inhibit breathing.
1.1 Abstract
Caffeine is the main treatment for apnea in preterm neonates, but its interactions with other respiratory stimulants like progesterone are unknown. We tested the hypothesis that the addition of progesterone to caffeine treatments further stimulates ventilation. Newborn rats were treated with water (control) or caffeine (15 mg/kg) by daily gavage between the postnatal (P) days 3-12. At P4 and P12, we measured apnea frequency, ventilatory responses, and metabolic parameters under both normoxia and hypoxia (12% O2, 20 min) following an
acute administration of either saline or progesterone (4 mg/kg; i.p.). Progesterone injection increased the serum levels of both progesterone and its neuroactive metabolite allopregnanolone. Progesterone had no effect on ventilation in control rats under normoxia. Progesterone depressed ventilation in P12 caffeine-treated rats under normoxia and hypoxia and increased apnea frequency in both P4 and P12 rats. Because allopregnanolone is an allosteric modulator of GABAA receptors and caffeine may enhance GABAergic inhibition
in newborns, we studied the effects of the GABAA receptor antagonist bicuculline at 0, 1, 2,
and 3 mg/kg doses and allopregnanolone (10 mg/kg dose) in P12 rats. In caffeine-treated rats, bicuculline enhanced ventilation, while allopregnanolone decreased ventilation and increased total apnea time. Progesterone had no effect on ventilation and apnea frequency in caffeine-treated rats injected with finasteride, which blocks the conversion of progesterone to allopregnanolone. We conclude that combining progesterone and chronic caffeine therapy is not an option for the treatment of persistent apnea in preterm neonates, unless the effects of allopregnanolone could be counteracted.
Keywords: Apnea, Premature Infant, Central Nervous System Stimulants, Central Nervous System Depressants.
1.2 Résumé
La caféine est le principal traitement de l'apnée chez les nouveau-nés prématurés, mais ses interactions avec d'autres stimulants respiratoires comme la progestérone sont inconnues. Nous avons testé l'hypothèse que l'ajout de progestérone aux traitements à la caféine stimule davantage la ventilation. Les rats nouveau-nés ont été traités avec de l'eau (controle) ou de la caféine (15 mg / kg) par gavage quotidien entre les jours 3 à 12 postnatals (P). À P4 et P12, nous avons mesuré la fréquence de l'apnée, les réponses ventilatoires et les paramètres métaboliques sous normoxie et hypoxie (12% O2, 20 min) après une administration aiguë de solution saline ou de progestérone (4 mg / kg; i.p.). L'injection de progestérone a augmenté les taux sériques de progestérone et de son métabolite neuroactif allopregnanolone. La progestérone n'a eu aucun effet sur la ventilation chez les rats controle sous normoxie. La progestérone a diminué la ventilation chez les rats traités à la caféine P12 sous normoxie et hypoxie et a augmenté la fréquence d'apnée chez les rats P4 et P12. Parce que l'allopregnanolone est un modulateur allostérique des récepteurs GABAA et que la
caféine peut augmenter l'inhibition GABAergique chez les nouveau-nés, nous avons étudié les effets de l'antagoniste des récepteurs GABAA bicuculline à des doses de 0, 1, 2 et 3 mg /
kg et allopregnanolone (dose de 10 mg / kg) chez les rats P12. Chez les rats traités à la caféine, la bicuculline a amélioré la ventilation, tandis que l'allopregnanolone a diminué la ventilation et augmenté le temps d'apnée total. La progestérone n'a eu aucun effet sur la ventilation et la fréquence des apnées chez les rats traités à la caféine auxquels on a injecté du finastéride, ce qui bloque la conversion de la progestérone en allopregnanolone. Nous concluons que l'association de la progestérone et de la caféine chronique n'est pas une option pour le traitement de l'apnée persistante chez les nouveau-nés prématurés, à moins que les effets de l'allopregnanolone ne puissent être neutralisés.
1.3 Introduction
In premature newborns, recurrent apneas are systematically treated to prevent long-term neurocognitive disorders (Martin & Wilson, 2012). Methylxanthines such as aminophylline, theophylline and caffeine stimulate breathing by antagonizing adenosine A1
and A2A receptors. Caffeine is currently the mainstay pharmacological treatment for apnea
(Zhao et al., 2011; Spitzer, 2012). However, a substantial percentage of apnea persists despite adequate treatment with caffeine, particularly in neonates born before 28 weeks of gestation (Zhao et al., 2011; Martin & Wilson, 2012; Spitzer, 2012). This suggests that other mechanisms may also work to inhibit breathing.
Progesterone is a well-known respiratory stimulant (Behan & Wenninger, 2008) that reduces apnea frequency in adult women (Shahar et al., 2003) and has been listed among the drugs that should be used in studies of apnea of prematurity (Finer et al., 2006). However, the impact of progesterone on neonatal respiratory physiology is poorly understood, and this lack of knowledge prevents the potential for clinical use. In newborn rats, acute progesterone administration enhanced the respiratory response to hypoxia in a dose- and age-dependent manner (Hichri et al., 2012; Bairam et al., 2013a), particularly in rats younger than 7 days old, but progesterone reduced apnea frequency only in 1-day-old rats. However, chronic progesterone administration through maternal milk reduced apnea frequency and enhanced the respiratory response to hypoxia in 10-day-old rats (Lefter et al., 2007). Conversely, acute administration of caffeine enhanced basal respiration and decreased apnea frequency to a greater extent in 12-day-old compared with in 4-days-old rats (Bairam et al., 2013b). Therefore, these two drugs appear to have different effective time windows of ventilatory regulation during postnatal development.
We asked whether acute progesterone administration might help to reduce apnea frequency and enhance the hypoxic ventilatory response in rats chronically treated with caffeine. To this end, we used a model of chronic caffeine administration in newborn rats (15 mg/kg/day, once/day by oral gavage between postnatal days 3 to 12) to mimic the routine daily usage of caffeine in the clinic (administered orally for several days or weeks). To investigate age-dependent effects, we studied rats at 4 and 12 postnatal (P) days. Surprisingly, our results showed that progesterone increased apnea frequency in P4 and P12