HAL Id: tel-01242554
https://tel.archives-ouvertes.fr/tel-01242554
Submitted on 13 Dec 2015HAL is a multi-disciplinary open access
archive for the deposit and dissemination of sci-entific research documents, whether they are pub-lished or not. The documents may come from teaching and research institutions in France or abroad, or from public or private research centers.
L’archive ouverte pluridisciplinaire HAL, est destinée au dépôt et à la diffusion de documents scientifiques de niveau recherche, publiés ou non, émanant des établissements d’enseignement et de recherche français ou étrangers, des laboratoires publics ou privés.
Interplay between autophagy and the primary cilium :
Role in mechanical stress integration
Idil Orhon
To cite this version:
Idil Orhon. Interplay between autophagy and the primary cilium : Role in mechanical stress in-tegration. Cellular Biology. Université Pierre et Marie Curie - Paris VI, 2014. English. �NNT : 2014PA066672�. �tel-01242554�
Université Pierre et Marie Curie (Paris 6) Ecole doctorale Physiologie et Physiopathologie
2014
DOCTORAL THESIS for University Pierre et Marie Curie Paris 6 Public Defense : 11 December 2014 by
Idil ORHON
entitled
INTERPLAY BETWEEN AUTOPHAGY AND THE PRIMARY CILIUM:
ROLE IN MECHANICAL STRESS INTEGRATION
Members of the Jury:
Professor Joëlle SOBCZAK-THEPOT President
Doctor Alexandre BENMERAH Reporter
Professor Fulvio REGGIORI Reporter
Doctor Vincent GALY Examinator
Doctor Sophie PATTINGRE Examinator
Doctor Isabelle BEAU Thesis Director
Doctor Patrice CODOGNO Thesis Director
Institut Necker-Enfants Malades (INEM)
INSERM UMR 1151 -CNRS UMR 8253 Université Paris Descartes-Sorbonne Paris Cité Laboratoire de l’homéostasie cellulaire et signalisation en physiopathologie hépatique et rénale
3
ACKNOWLEDGMENTS
Firstly, I would like to thank the members of the jury, to Alexandre Benmerah and Fulvio Reggiori for accepting to evaluate this manuscript, to Joelle Sobczak for accepting to be the president of the jury and to Sophie Pattingre and Vincent Galy for accepting to be examiner of this work.
Foremost, I would like to express my gratitude to my thesis supervisors Patrice Codogno and Isabelle Beau for giving me the opportunity to work and learn from them. I am sincerely grateful to Patrice Codogno for his expertise, experience and confidence in me throughout these years. Without his extensive knowledge on autophagy but also on “the outside world”, I certainly could not evolve as a researcher during these years. I am deeply thankful for Isabelle Beau and her presence in my PhD journey starting from day one whether from the next door or in distance. She set an inspiring example for a woman in science with her passion of bench work and guidance.
I am greatly thankful to Ana-Maria Cuervo for giving me the opportunity to be part of an inspiring collaboration that changed my perspective on many different levels. Working in her lab was a condensed learning experience with a very dynamic and kind group of people. That leads to a sincere thanks and gratitude to Olatz Pampliega, an amazing collaborator and scientist whom I feel lucky to have worked with, learnt from and hope to work with again. Without them this work could not have happened.
I would also like to acknowledge Gerard Friedlander, for his support and contribution to this work.
A very special thanks goes to the “primary” Nicolas Dupont. During the last year, his presence changed the dynamism of our lab and this project. Without our brainstorms, his motivation and his passion for pubmed search, I wouldn’t see the finish line. I would especially like to thank him for his input and time on this manuscript.
I would like express my thanks to my chocolate loving group of colleagues with whom I feel very lucky to work with daily: Ahmeeeeeeed Hamai who was ready to answer all of my questions at any time with a smile, Joelle Botti and Chantal Bauvy for their kindness and
4
support and Etienne Morel who was a support even before joining the lab, Anna Chiara Nascimbeni and Maryam Mehrpour.
I’d like to thank sincerely to the Chatenay-Malabry team with whom I started this journey: Lina Mouna for her friendship and the Turkish/Syrian coffee time, Francoise Magnin for her daily kindness, Mathieu Bertrand, Marion Lussignol, Audrey Esclatine, Marie-Francoise Bernet-Camard, Anne Greffard and Raymonde Amsellem.
I want to thank everybody from Co-lander team “across the hall”, with a shout-out to Christine and Valérie for their management skills and not leaving us in chaos. I would also like to thank to Marie Courbebaisse and Dominique Prié for their insight on the kidney. I must acknowledge the platforms that helped me: Claudine from Trans-Prot of IPSIT, Nicolas and Raphaelle from IMAGINE.
I must thank Philippe Cardot for his support, Chiara Maiuri, Ezgi Tasdemir and Devrim Gozuacik for their supervision during my ignorant years and for teaching me how to work on the bench, which I love.
I would also like to thank Rosalind, Henrietta, Simone, Charles, Pablo, Samed, Halit Ziya and Yoda for the inspiration.
Last but not least I have to thank my family and friends who make this journey worth sharing. I am grateful for the love and support of my brother, mother and father. Without the patience of my mom and dad, I could not have pursued an education in science so far. I am also thinking of my dear grandmother who passed away at the beginning of this craziness and who I wished could have seen the end of it all. I am very lucky and grateful to have too many friends to name here who shared this experience with me. However, I cannot fail to acknowledge these wonderful people for their love, jokes and support: Seray, Yasemin, Burge, Idil, Irem and especially Isil who supported my daily neurosis towards the end. I love them dearly.
5
TABLE OF CONTENT
Abbreviations ……….7 Table of Illustrations………..9 Summary………...10 Résumé………..11 I. Introduction………..121. Cilium: Structure and Signaling………...13
1.1. Structure ………...13
1.1.1. Motile Cilia and Flagella ………..13
1.1.2. Primary Cilia………..14
1.1.3. Ciliogenesis and Ciliary Subdomains………..15
1.1.4. Intraflagellar Transport Mechanism………....18
1.2. Ciliary Signaling………20
1.2.1. Hedgehog Signaling………...20
1.2.2. PDGF Pathway………..23
1.2.3. Wnt Pathway………..23
1.2.4. Calcium Signaling………..24
1.2.5. Fluid flow and Fluid Shear Stress………..…..25
2. Autophagy……….30
2.1. Role of Autophagy………...….30
2.2. Different Types of Autophagy………..…...30
2.2.1. Macroautophagy………30
2.2.2. Chaperone-mediated Autophagy ………..31
2.2.3. Microautophagy………... ….34
2.3. Autophagosome Machinery and Biogenesis ………34
2.3.1. Initiation……….35
2.3.2. Elongation………..…40
2.3.3. Maturation………..41
2.3.4. The origin of the Autophagosome……….42
2.4. Regulation of Autophagy ……….46
6
2.4.2. Post-translational Modifications of Autophagic Machinery…..50
2.4.3. Transcriptional and Epigenetic Regulations of Autophagy…...53
2.5. Physiological Role of Autophagy………..55
2.5.1. Selective Autophagy………..56
2.5.2. Autophagy during pre- and post-natal state………...58
2.5.3. Autophagy in the liver………...58
2.5.4. Autophagy in the brain………..59
2.5.5. Autophagy in the muscle………...59
2.5.6. Autophagy in the intestine……….60
2.5.7. Autophagy in the pancreas………60
2.5.8. Autophagy in the lung………61
2.5.9. Autophagy in innate and adaptive immunity……….61
2.5.10. Pathophysiological Role of Autophagy……….63
2.6. The Role of Autophagy in Kidney Physiology and Physiopathology...67
2.6.1. Physiology of the Kidney: An Introduction………...67
2.6.2. Autophagy in the Kidney: Health and Disease………..71
II. AIMS of the study……….77
III. Materials and Methods………..80
IV. Results ………...………...86
1. Objective1:Study the functional interaction between autophagy and primary cilium..87
1.1.Results……….89
1.2. Discussion 1………..90
2. Objective 2: Study the role of autophagy in integration of mechanical stress via primary cilium……….94
2.1. Results………96
2.2. Discussion 2………..106
V. General Discussion and Perspectives………..111
BIBLIOGRAPHY……….…119
7
Abbreviations
3-MA: 3-methyladenine 4EBP-1: 4E binding protein-1
ADPKD: autosomal dominant polycystic kidney disease
aIFT : anterograde IFT AKI: acute kidney injury
AMPK: AMP-activated protein kinase APC: adenomatosis polyposis complex AQP: aquaporin
ATG: autophagy related ATP: adenosine triphosphate BB : Basal body
BBS: Bardet–Biedl syndrome Bcl-2: B cell lymphomas CKD: chronic kidney disease
CMA: chaperone-mediated autophagy
COPD, chronic obstructive pulmonary disease; CQ: Chloroquine
CTS: Ciliary targeting sequence DN: diabetic nephopathy
EndoMT: endothelial-to-mesenchymal transition
ER: endoplasmic reticulum
ERGIC: ER-Golgi intermediate compartment FGF: fibroblast growth factor
FOXO: conserved forkhead box O FSS: Fluid Shear Stress
GFP: green fluorescent protein
Gli: glioma-associated oncogene family GPCRs: G-protein-coupled receptors HDAC6: histone deacetylase-6 Hh: Hedgehog
IFN: interferon
IFT : Intraflagellar transport IKK: IκB kinase
IPTG: Isopropyl β-D-1-thgalactopyranoside JNK1: Jun NH2-terminal kinase 1
KEC: Kidney epithelial cell
MAPK: mitogen-activated protein kinase MC: motile cilium
MDCK: Madin-Darby canine kidney MHC: major histocompatibility complex mTORC1: mammalian target of rapamycin complex 1
NF-κB: nuclear factor-κB NHE1: Na+/H+ exchanger 1
OFD1: oral-facial-digital syndrome type 1 PAS: pre-autophagosomal structure PC: Primary cilia
PC1: polycystin 1 PC2: polycystin 2 PCP: planar cell polarity
8 PDGFRα: platelet-derived growth factor
receptor alpha
PE: phospholipid phosphatidylethanolamine PI3K: class III phosphatidylinositol 3-kinase PI3P: phosphatidylinositol 3-phosphate PKD: polycystic kidney disease Ptc1: patched 1
rIFT : retrograde IFT
ROS: reactive oxygen species S6K: ribosomal protein S6 kinase-1 Shh: sonic hedgehog
Smo: smoothened
Sufu: suppressor of fused protein TFEB: transcription factor EB TGF-β: transforming growth factor-β TGN: trans-Golgi networks
UUO: Unilateral ureteral obstruction Vangl2: Van Gogh-like 2 protein
9
Table of Illustrations
Figure 1: Cilia Structure Figure 2: Classification of cilia Figure 3: Primary Ciliogenesis models Figure 4: Intraflagellar Transport Figure 5: Hedgehog signalingFigure 6: Mechano-sensing by the cell
Figure 7: The different types of autophagy Figure 8: The Autophagic pathway
Figure 9: Autophagosome machinery
Figure 10: The Origin of the Autophagosome
Figure 11: mTOR dependent regulation of Autophagy Figure 12: Physiological Roles of Autophagy
Figure 13: Selective autophagy
Figure 14: Autophagy and Pathophysiology Figure 15: Physiology of the nephron
Figure 16: Polycystic Kidney Disease and Fluid Flow in the Kidney Figure 17: Flow stimulates the autophagic flux in MDCK cells Figure 18: Inhibition of ciliogenesis impairs flow-induced autophagy
Figure 19: Inhibition of autophagy impairs primary cilium cell size regulation
Figure 20: Autophagic machinery (ATG16L1) recruited to the basal body upon fluid flow Figure 21: Signaling regulation of primary cilium-dependent autophagy activation during flow Table 1: Pathways regulated by mechanical stress via Primary Cilia
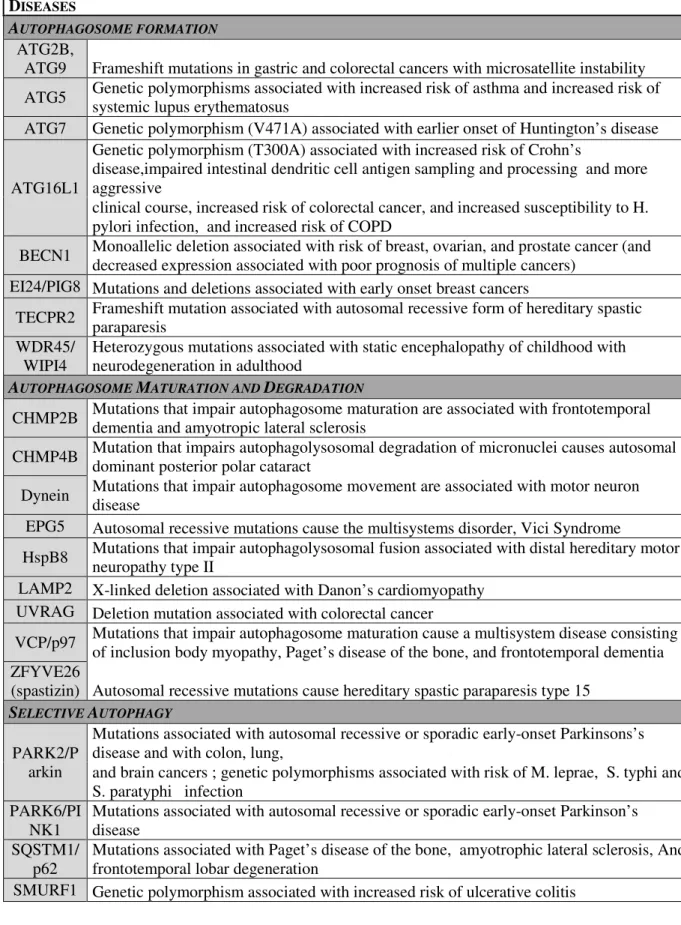
Table 2: Mutations in Autophagy Genes and associated human diseases
Table 3: Structure and signaling from the cilium and relationship with the autophagic pathway Table 4: Cilia-related proteins degraded by autophagy
10
Summary
Motile and primary cilia are microtubule-based structures located at the cell surface of many cell types. Cilia govern cellular functions ranging from motility to integration of mechanical and chemical signaling from the environment. In this work we investigate the potential cross-talk between the primary cilium and macroautophagy. Macroautophagy or self-eating is a lysosomal degradative pathway that allows cells to adapt to various stress situations. The rational for the study was based on the survey of the literature showing that many stress situations that trigger primary cilium signaling also stimulates autophagy (serum starvation, calcium mobilization, cell cycle arrest). In the first part of the study we showed that inhibition of ciliogenesis severely impairs serum-induced autophagy in mouse embryo fibroblasts, kidney epithelial cells and neurons. We also showed that in response to serum deprivation many Autophagy-related proteins (Atg proteins) involved in autophagosome formation are co-localized with cilium subdomains (axoneme and basal body). Notably the protein Atg16L1 is co-transported to the basal body with the ciliary protein IFT20. The localization of Atg16L1 to the basal body as well as serum-induced autophagy were severely impaired by inhibiting the Hedgehog signaling pathway either genetic or pharmacological approaches. We also showed that invalidation of ATG genes induced an increase in primary cilium length in basal condition. Cilia were functional in ATG-deficient cells because of the presence of a ciliary pocket and the activation of the Hedgehog signaling pathway. Finally we identified IFT20 as a substrate for autophagy. Thus autophagy is required to regulate the level of IFT20 and consequently that of the length of the primary cilium. In the second part of the work we investigate the role of the cross-talk between autophagy and the primary cilium in regulating the size of kidney epithelial cells. Previous studies have shown that the primary cilium plays a central role in regulating cell size and cell volume. This regulation is important to keep the physiological functions of tubular renal cells by maintaining the planar polarity in kidney tubule. By applying a liquid flow of 1 dyn/cm2 to MDCK or mouse kidney epithelial cells to mimic physiological conditions, we show that the flow induces autophagy and reduction of the cell volume. In absence of cilium we observed that autophagy is not induced and that the cell size/volume is not responsive to the mechanical stress. Finally we showed that ablation of autophagy led also to an impairment of flow-dependent regulation of cell size/volume in ciliated kidney epithelial cells. In conclusion primary cilium-dependent autophagy plays a major role in controlling the epithelial kidney cell size/volume during mechanical stress induced by fluid flow.
Key words: Macroautophagy, IFT, ATG, primary cilium, fluid flow, cell size/volume, kidney epithelial cells
11
Résumé
Les cils primaires et motiles sont des structures microtubulaires présentent à la surface de nombreux types cellulaires. Les structures ciliées contrôlent de nombreuses fonctions allant de la motilité cellulaire à l’intégration par la cellule de stimuli chimiques et mécaniques. Au cours de cette thèse, nous avons étudié le dialogue entre le cil primaire et l’autophagie, un processus d’autodigestion qui permet à la cellule de s’adapter à des situations de stress. L’hypothèse de ce dialogue reposait sur l’analyse de la littérature montrant que de nombreux médiateurs (calcium, carence en sérum, arrêt du cycle cellulaire) stimulent à la fois l’activité ciliaire et l’autophagie. Dans un premier temps de notre étude nous avons montré que l’inhibition de la ciliogenèse altère l’induction de l’autophagie en réponse à la carence en sérum dans des fibroblastes d’embryon de souris, des cellules épithéliales rénales et des lignées de neurone. Nous avons aussi montré que la carence en sérum induisait une redistribution de nombreuses protéines Atg (Autophagy-related), protéines impliquées dans la biogenèse de l’autophagosome, au niveau du cil primaire (soit au niveau du corps basal soit au niveau de l’axonème). Particulièrement la protéine Atg16L1 est co-transportée vésiculairement au corps basal avec la protéine ciliaire IFT20. L’inhibition génétique ou pharmacologique de la voie de signalisation Hedgehog inhibe à la fois le transport de la protéine Atg16L1 au corps basal et l’induction de l’autophagie en absence de sérum. Nous avons aussi montré que l’invalidation de gènes ATG est associée à une ciliogenèse accrue. Dans ces conditions nous avons conclu sur des bases morphologiques et biochimiques que ces cils primaires sont fonctionnels. La protéine IFT20 s ‘accumule dans les cellules déficientes en autophagie et est dégradée par autophagie dans les cellules sauvage en présence de sérum. Ces résultats montre que l’autophagie basale (autophagie observée en présence de sérum) est un mécanisme qui contribue au contrôle de la croissance du cil primaire. Dans une deuxième partie du travail nous avons étudié l’importance de l’autophagie dans la réponse cellulaire à stress mécanique. Le contrôle de la taille et du volume des cellules épithéliales rénales est un élément important pour maintenir la polarité planaire des cellules tubulaires. Cette propriété est dépendante du cil primaire. Au cours de l’application d’un flux de liquide (1 dyn/cm2) concomitamment à la réduction du volume et de la taille cellulaire nous avons observé une stimulation de l’autophagie. Cette réponse autophagique dépend du cil primaire. L’invalidation de l’autophagie dans des cellules épithéliales ciliées abolit le contrôle du volume et de la taille cellulaire dans les cellules épithéliales rénales. L’ensemble de ces résultats montre le dialogue qui existe en l’autophagie et le cil primaire et l’importance de ce dialogue dans l’intégration par la cellule du stress mécanique.
Mots clés : Macroautophagie, IFT, ATG, cil primaire, stress mécanique, volume et taille cellulaire, cellules épithéliales rénales
12
13
I.
INTRODUCTION
1.
Cilium: Structure and Signaling
1.1. Structure
Cilia are microtubule-based organelles that are present at the surface of many vertebrate cells (Satir and Christensen 2007; Satir, Pedersen et al. 2010). Conventionally, cilia are classified as motile and non-motile according to their microtubule pattern and motility in the organism.
1.1.1. Motile Cilia and Flagella
Motile cilium (MC) is formed by a microtubule based axoneme of 9 outer microtubule doublets and 2 single central microtubules extending from a basal body. Two dynein motors and a radial spoke are attached to the doublets along the axoneme, giving the motility to the organelle and thus the ability to generate fluid and beat-like movement (Figure 1). Motile cilia are found on the surface of the epithelial cells lining the airways and reproductive tracts, as well as on epithelial cells of the ependyma and in the brain and on the embryonic node. Motility of the cilium in these epithelial cells is responsible for mucociliary clearance and ependymal flow. The 9+2 structure of MC is identical to the flagella, which are found in sperm and single-celled eukaryotes (i.e. Paramecium) and that serve traditionally in cell locomotion (Pedersen and Rosenbaum 2008). MC and flagella differ from each other based on their pattern of movement and the number per cell. However, new studies on the structure and function of MC and flagella in different tissues or species show that they can have sensory roles such as MC in the tracheal epithelial cells which have bitter taste receptors or
mammalian sperm which possess pH-dependent Ca2+ channel (Takeda and Narita 2011) (See
14
1.1.2. Primary Cilia
Primary cilium (PC), a solitary non-motile cilium, consists of an axoneme of 9 outer doublet microtubules extending from a basal body (BB) that is derived from the older (mother) centriole of the centrosome, and which lacks the 2 single central microtubules involved in motility (Satir and Christensen 2007) (Satir, Pedersen et al. 2010). PC projects from surface of the plasma membrane of a wide range of polarized cells including stem, epithelial, endothelial, connective-tissue muscle cells and neurons (Figure 1).
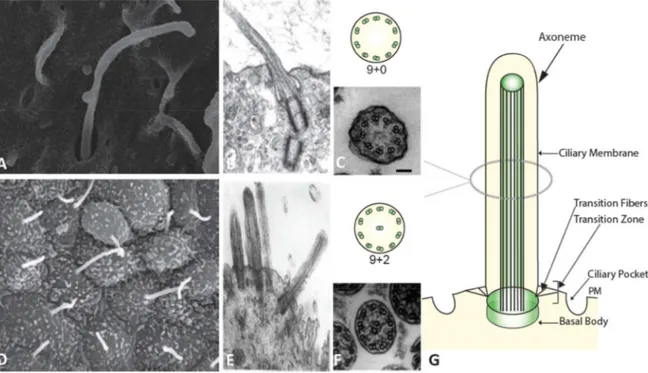
Figure 1: Cilia Structure
(A) Scanning electron microscopy (SEM) of primary cilia from mouse embryonic fibroblasts. (B) Electron microscopy of PC. (C) Electron microscopy (EM) image on the bottom and schematic representation of the 9 microtubule doublets of PC on the top. (D) SEM image of motile cilia from mouse tracheal epithelium. (E) EM image of motile cilia. (F) EM image on the bottom and schematic representation of 9+2 microtubule doublets of the motile cilia on the top. (Images from Pampliega et al. 2013, Satir et al. 2010 and Sorokin 1968) (G) Schematic representation of the cilium (Orhon et al. 2014 review in press)
15 Figure 2: Classification of cilia
Schematic diagram showing the classification of motile “9+2”, motile “9+0”, and non-motile “9+0” cilia. The diagram shows the coordinated synchronous motion of motile “9+2” cilia, the rotary motion of motile “9+0” nodal monocilium, and non-motile “9+0” primary monocilium (modified from (Ferkol and Leigh 2012))
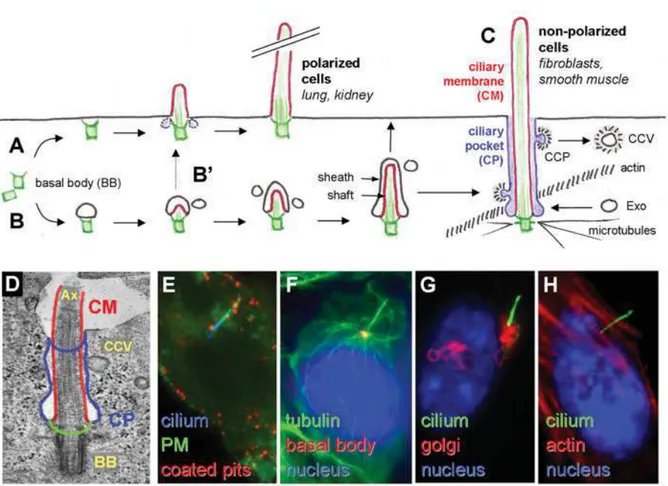
1.1.3. Ciliogenesis and Ciliary Subdomains
Ciliogenesis occurs in quiescent cells when the cells enter the G0 phase. When the cells decide to re-enter in a cell cycle, cilium resorption occurs followed by the S phase. Ciliogenesis and cell cycle is regulated by post-translational modifications (Avidor-Reiss and Gopalakrishnan). Sorokin proposed two models of primary ciliogenesis upon TEM observations from epithelial cells and fibroblasts (Sorokin 1962; Sorokin 1968); an ‘extracellular’ pathway and an ‘intracellular’ pathway. In the polarized cells, the ‘extracellular’ model the centrioles move directly to the plasma membrane and the mother centriole serve as a docking center for the growth of the axoneme thus forming the basal body
16
of the cilium (Benmerah 2012). This model is observed in the ciliated cells of lung or kidney tissues. Alternatively, in non-polarized cells, as in the case for fibroblasts, an intracellular ciliary vesicle grows around the mother centriole together with the axoneme. This vesicle will later fuse with the plasma membrane, which will result in the protrusion of PC to the extracellular space. The BB is formed of a 9 triplet microtubules (Figure 3). The TEM images show that PC can remain invaginated to the plasma membrane which resembles to the flagellar pocket. The ciliary pocket is continuous with the plasma membrane, but functionally different. This small but significant domain of the PC is established after fusion of the plasma membrane with the intracellular ciliary vesicle that grows on top of the BB (Ghossoub, Molla-Herman et al. 2011). The origin and function of the ciliary pocket is not clearly known but it is currently a topic of intense research (Benmerah 2012). The ciliary pocket is characterized by the presence of clathrin coated vesicles resulting from the vesicular trafficking that originates and ends in this region. Acting as a specific endocytic membrane, the ciliary pocket has been described to regulate the transforming growth factor-β (TGF-β) pathway (Molla-Herman, Ghossoub et al. 2010; Clement, Ajbro et al. 2013). The presence or absence of the ciliary pocket depends on the cell type and can be explained by different kinetics during ciliogenesis; for example, ciliary pocket is common in fibroblasts but has rarely been observed in epithelial cells. This rare observation of the pocket in epithelial cells can be explained by a short-lived intracellular pathway of the ciliary pocket docking to the PM (Ghossoub, Molla-Herman et al. 2011). PC is not isolated from the cytoplasm but is highly compartmentalized; it is surrounded by a specialized region of the plasma membrane that holds a different concentration of channels and receptors and the axoneme is separated from the rest of the intracellular compartments by the “transition zone”. The transition zone consists of the “ciliary necklace” and transition fibers and is evolutionarily conserved. It is characterized by the ciliary necklace formed by Y-shaped fibers that connect the microtubule doublets to the ciliary membrane at the base of the cilium distal to the BB. Moreover, this zone is mostly characterized by TEM data (Reiter, Blacque et al. 2012). This ciliary subdomain is highly organized with a molecular structure resembling that of the nuclear pore and serves as a border control where different, precursors and intraflagellar transport (IFT) proteins, along with proteins of different signaling pathways are concentrated and regulated for entry to the axoneme (Kee, Dishinger et al. 2012). The composition of the transition zone and subdomains is not fully known, however several studies suggest that distal appendage proteins play an important role (Avidor-Reiss and Gopalakrishnan 2013). During ciliogenesis, the mother centriole anchors to the associated membrane; the ciliary vesicle membrane or the
17
plasma membrane. The mother centriole can be distinguished from the daughter centriole by the presence of subdistal and distal appendages. Distal appendages are necessart for ciliogenesis. OFD1 (oral-facial-digital syndrome1) protein is associated with the distal ends of centrioler microtubules and is necessary for centriole docking via the distal appendages which forms the transition fibers at the base of the cilium (Reiter, Blacque et al. 2012; Singla, Romaguera-Ros et al. 2010).
Figure 3: Primary Ciliogenesis models
(A)The “extracellular pathway” which takes place primarily in polarized cells where the mother centriole of the centrosome (green) directly docks to the plasma membrane forming the basal body (BB) and the primary cilium (PC, membrane in red) grows directly in the extracellular milieu. (B) The “intracellular pathway” which takes place primarily in fibroblasts during which a primary ciliary vesicle interacts with the distal appendages of the mother centriole, the axoneme grows then within this vesicle, which elongates thanks to the fusion with incoming secondary vesicles then fusing with the plasma membrane and forming the ciliary pocket (CP).(CP, blue). (B’) Ciliogenesis model proposed for polarized cells with a faster fusion event with the plasma membrane and/or complete extrusion of the cilium. (C) CP acts as a specific endocytic membrane domain (CCP, CCV clathrin-coated pits and vesicles respectively, CM is ciliary membrane). (D) PC in RPE1, human retinal pigment epithelial cells. (E-H) IF images of RPE1 PC. (Ghossoub, Molla-Herman et al. 2011)
18
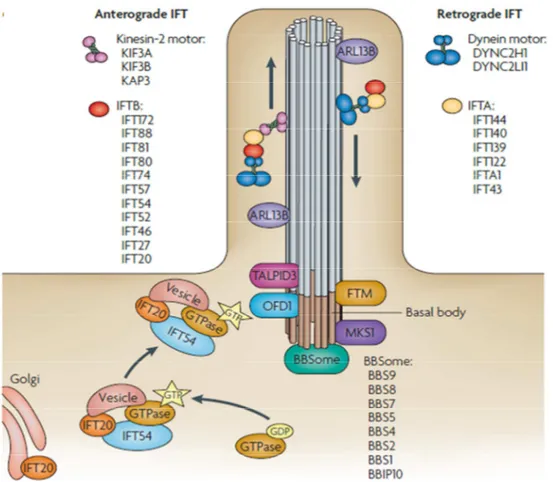
1.1.4. Intraflagellar Transport Mechanism
A bi-directional movement in the Chlamydomonas flagella was fist observed by Rosenbaum lab and identified as the intraflagellar transport (IFT) (Kozminski, Johnson et al. 1993). IFT complexes are conserved in ciliated organisms and have different functions. The IFT mechanism plays an essential role in the assembly and maintenance of the cilia or flagella and during the transport and trafficking of ciliary components such as cargo along the axoneme (Rosenbaum and Witman 2002; Sung and Leroux 2013). The bi-directional trafficking along the polarized microtubules of the axoneme is maintained through the action of two motor protein families: kinesins and dyneins (Figure 4) (Pedersen and Rosenbaum 2008). Anterograde IFT (aIFT) trafficking with base-to-tip cargo direction, depends on the heterotrimeric kinesin-2 motor, also known as KIF3 motor complex. This complex consists of two kinesin-2 family proteins KIF3A, KIF3B and KIF-associated protein 3 (KAP3). Retrograde IFT (rIFT) with tip-to-base cargo direction depends on the dynein motor that consists of cytoplasmic dynein 2 heavy chain 1 (DYNC2H1) and cytoplasmic dynein 2 light intermediate chain 1 (DYNC2LI1)) (Pedersen, Veland et al. 2008). The kinesin and dynein motors are conserved in all ciliated organisms with the exception of Plasmodium falciparum (Sung and Leroux 2013). Two large complexes composed of more than 20 proteins called IFT A and IFT B are essential for the building and trafficking along the cilium, the first for retrograde and the latter for anterograde trafficking. Mutations in the two IFT subunits have different consequences; IFT B protein defects lead to absent or shortened cilia whereas defects in IFT A proteins lead to a bulged cilia (Goetz and Anderson 2010). The docking and assembly mechanism of IFT particles are not well characterized. Several IFT proteins are identified over the years in several organisms and a number of these proteins possess specialized roles in ciliogenesis, antero- or retrograde trafficking and transport of ciliary components to the cilia or flagella. Some subunits of IFT complexes, such as IFT20 and DYF-11/MIP-T3, which are part of the IFT B complex are found in other cellular compartments from where they facilitate transport of vesicles with specific cargo to the BB and axoneme (Pedersen, Veland et al. 2008). Continuous shuttling of IFT20 protein between the Golgi and the PC is required for ciliogenesis and cilia dependent signalizations (Follit, Tuft et al. 2006; Keady, Le et al. 2011). OFD1 protein plays an important role in association with IFT assembly and is required for the recruitment of IFT88 protein (Singla, Romaguera-Ros et al. 2010). Maintenance of the cilia also depends on the BBsome, a complex formed by proteins encoded by genes mutated in Bardet–Biedl syndrome (BBS). Ciliary targeting sequence
19
(CTS) is recognized by the BBsome and serves target the proteins with CTS to the cilia in a BBSome dependent manner. Several proteins involved in the vesicular trafficking such as the small GTPase Rab8 or Rab11 are evolutionarily conserved in the vesicular trafficking and also in cilia assembly via IFT and BBsome (Sung and Leroux 2013). The mechanisms of the transport and trafficking of proteins involved in various cilia-dependent signaling pathways are not known in details (Goetz and Anderson 2010). However various proteins are observed to interact with IFT. For example the Chlamydomonas orthologue of the calcium channel PKD2 is IFT-dependent (Huang, Diener et al. 2007).
Figure 4: Intraflagellar Transport
Cargo is transported from the base to the tip of the cilium along the microtubule axoneme by the kinesin-2 motor (which consists of KIF3A, KIF3B and KAP3) together with the intraflagellar transport A (IFT A) complexe regulating the anterograde IFT. The retrograde IFT of the cargo, from the tip to the base of the cilium, is mediated by the dynein motor (which consists of cytoplasmic dynein 2 heavy chain 1 (DYNC2H1) and cytoplasmic dynein 2 light intermediate chain 1 (DYNC2LI1)) and by IFT B complexe. Certain basal body proteins also influence ciliary trafficking. Among these are components of the BBSome. (Goetz and Anderson 2010)
20
1.2. Ciliary Signaling
The major function of PC is sensing the extracellular medium which is mediated by different signaling pathway receptors, ion channels, transporter proteins and their downstream molecules located to the axoneme or to the basal body. PC responds to different sensory modalities such as mechanical stimulus (bending of the cilia following a shear stress) or chemosensation (specific ligand, growth factor, hormone or morphogen recognition). A variety of signaling pathways are coordinated through this organelle during development, tissue homeostasis, cell migration, cellular differentiation, cell cycle and apoptosis. Examples of such pathways are Hedgehog (Hh) pathway, PDGF pathway; Wnt pathway and Ca2+ signaling cascade (Eggenschwiler and Anderson 2007; Satir, Pedersen et al. 2010).
1.2.1. Hedgehog Signaling
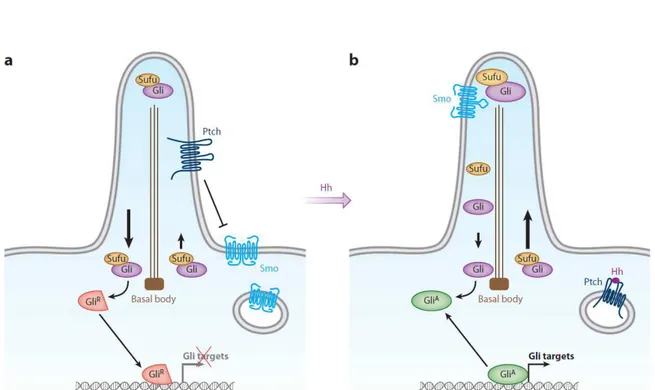
Hedgehog (Hh) signaling is one of the pathways that orchestrate development in animals, along with other signaling pathways such as Wnt, Notch, TGF- β and fibroblast growth factor (FGF). The name of the pathway originates from the short and “spiked” phenotype of the cuticle of the Hh mutant Drosophila larvae. Hh pathway is researched in detail in Drosophila and the mammalian pathway remains to be fully discovered in adult cells (Varjosalo and Taipale 2008). The Hh pathway is involved in regulation of cell proliferation, cell fate determination, in the epithelial-to-mesenchymal transition and in maintenance of homeostasis in adult cells (Simpson, Kerr et al. 2009; Briscoe and Therond 2013). The cellular trafficking of these pathways includes coordinated uptake, secretion and localizations of necessary molecules throughout the cell. A cell’s fate is determined according to the concentration gradient of the Hh proteins, which are diffusible lipid-modified morphogens. Primary cilium is a key coordinator of the mammalian Hh signaling pathway as its major signaling components are localized to the cilium, and IFT proteins are essential for trafficking of Hh molecules (Goetz and Anderson 2010). A recent study showed that in the ciliated olfactory sensory neurons of Drosophila, the Hh pathway is dependent on the presence of PC (Kuzhandaivel, Schultz, et al. 2014). This observation suggests that there may be two pathways for Hh mediation within the same organism; a PC-dependent and PC-independent pathway in non-ciliated cells.
21
There are three secreted Hh ligand member family proteins: Sonic hedgehog (Shh), Indian hedgehog, and desert hedgehog (Varjosalo and Taipale 2008; Ingham, Nakano et al. 2011). Shh is the most widespread ligand, whose auto-catalytically processed lipidated form is secreted into the extracellular matrix. It is expressed in the midline tissue and in the zone of polarizing activity of the limb bud during vertebrate embryogenesis. During organogenesis it is expressed during the development of most epithelial tissues (Varjosalo and Taipale 2008). Hh signaling depends on the balance between activator and repressor forms of the glioma-associated oncogene family (Gli) of zinc-finger transcription factors. The basic signaling pathway involves a 12-transmembrane domain receptor protein Patched 1 (Ptc1 or Ptch1), the 7-transmembrane domain protein Smoothened (Smo), the suppressor of fused protein (Sufu), and the transcription factors Gli members (Figure 5). Hh signaling is suppressed in the absence of Hh ligand through the physiological inhibitory effect of Ptc1 on Smo. When the pathway is activated by Hh ligand binding to Ptc1, Smo is released from Ptc1 and re-localizes to the PC axoneme where it inhibits Sufu activity. Sufu controls the processing and translocation of Gli family transcription factors from the cilium to the nucleus (Goetz and Anderson 2010).
The Gli protein family includes Gli1, Gli2, and Gli3. They are highly related but their biochemical properties differ from one another along with their functions in vivo. This family of zinc-finger transcription factors balances between activator and repressor forms. Activated Gli forms directly induce transcription of Hh target genes including genes encoding Ptc1 and Gli family members. The activity of Gli1 depends on Gli2, and Gli1 and Gli2 act primarily as transcriptional activators. Gli3 exists in two-forms: a non-processed, full-length form that can function as transcriptional activator and a truncated amino-terminal fragment that acts as transcriptional repressor (Robbins, Fei et al. 2012; Aberger and Ruiz 2014). Also, it has been recently described that the activity of Kif7 at the cilium creates a specialized compartment at the top of the cilia where the activity of the Gli proteins is regulated (He, Subramanian et al. 2014). The bifunctional Gli proteins have roles in embryogenesis and adult homeostasis as their target genes regulate cell proliferation (cyclin and Myc genes), cell death (Bcl-2), differentiation (genes encoding Forkhead family transcription factors), and stem cell renewal (Bmi-1). Target genes regulated by the Hh pathway depend on tissue and cell types (Jiang and Hui 2008; Briscoe and Therond 2013). Hh is a key morphogen for body patterning during development of various organs such as pancreas, kidney, lung, foregut, nervous system and limb. Target genes regulated by the Hh pathway may vary among tissues and cell types.
22
Misregulation of such diversified signaling pathway leads to various unfavorable developmental and pathological conditions including birth defects and tumorigenesis (Keady, Le et al. 2011).
Figure 5: Hedgehog signaling
(A) In the absence of ligand, Ptc1 (Ptch) localizes to the PC and prevents the entry of Smo into the axoneme. Low levels of Sufu-Gli protein complexes are localized to the cilium which promotes the repressor forms of Gli (GliR), inhibiting the transcription of target genes. (B) After activation of the pathway, Smo moves to the ciliary membrane promoting Sufu-Gli complexes translocation, dissociation and Gli (Gli2) accumulation at tip of the cilium. Activated Gli proteins (GliA) are transported out of the cilium by the dynein motor and intraflagellar transport (IFT) particles for the transcriptional activation of the target genes. (Hui and Angers 2011)
23
1.2.2. PDGF Pathway
Platelet-derived growth factor receptor alpha (PDGFRα), a widely expressed protein, localizes specifically to the PC in quiescent fibroblasts and induces activation of Akt, Mek1/2, Erk1/2, and Rsk pathways at the axoneme and in the basal body of the cilium (Satir and Christensen 2007). The PDGF pathway also induces cilia-dependent activation of MAPK and regulates proliferation, cell survival, and migration during embryogenesis and tissue homeostasis. Primary cilium-dependent PDGFRα activation regulates the ubiquitous plasma membrane Na+/H+ exchanger NHE1, which controls proliferation, cell survival, and migration (Schneider, Stock et al. 2009; Clement, Mally et al. 2013). Defects in this pathway can lead to different human pathologies including gastrointestinal stromal tumors, lung tumors, and ovarian carcinoma (Schneider, Clement et al. 2005).
1.2.3. Wnt Pathway
Canonical and non-canonical Wnt pathways are crucial during development (Kestler and Kuhl 2008; Angers and Moon 2009). The canonical Wnt pathway is regulated by β-catenin and controls proliferation, apoptosis, and cell fate determination via activation of target genes. Activation of Wnt target genes depends on ligand binding to the Frizzled receptor at the plasma membrane and subsequent β-catenin translocation into the nucleus. The stability of β-catenin depends on its degradation by the adenomatosis polyposis complex complex (APC) at the basal body. Recent studies have established the association of Wnt pathways with ciliary components such as kinesin motors. Transport of APC to its various cellular locations via kinesins plays a vital role in microtubule stabilization, activation of protein kinases, and cell polarization. These findings shed light on the contribution of the interaction of Wnt pathway with the PC in the regulation of cytoskeletal architecture (May-Simera and Kelley 2012).
The non-canonical Wnt pathway is controlled by a membrane protein called Van Gogh-like 2 (Vangl2) and regulates cytoskeletal changes, cell adhesion, migration, and polarity (May-Simera and Kelley 2012). Vangl2 functions in asymmetric positioning of motile cilia and in centrosome positioning, which is important for cytoskeletal rearrangement. Mutations of different ciliary proteins, such as Kif3a, IFT88, OFD1 (the protein mutated in oral-facial-digital syndrome type 1) result in dysregulation of planar cell polarity (PCP) and
24
are associated with hyper- or de-activation of the Wnt pathway depending of the tissue, timing, and localization of these proteins (Corbit, Shyer et al. 2008). PCP is critical for the development of various organs. Apart from the apical/ basal polarity that defines cellular orientation, epithelial cells can also be polarized within the plane of the tissue, orthogonal to the apical/basal axis which characterizes PCP (Carroll and Das 2011). Vertebrate tissues such as postnatal kidney tubule epithelia show PCP components such as oriented cell division which is characterized by the cell division parallel to the axis of kidney tubule elongation and in the plane of the epithelium. (Fedeles and Gallagher 2012).Components of the PCP pathway, such as Vangl2 and inversin (also called nephrocystin 2), localize at the PC, specifically at the basal body and the inversin compartment which is localized at the proximal segment of the cilium beyond the transition zone, during development (Shiba, Yamaoka et al. 2009). Inversin seems to serve as a molecular switch between canonical and non-canonical Wnt pathways by regulating Dishevelled protein degradation at the base of the cilium by the APC (May-Simera and Kelley 2012).
1.2.4. Calcium Signaling
The Ca2+-specific cation channel PC2, which forms an ion channel complex with the G-protein coupled receptor PC1 localized in the axoneme, regulates calcium signaling (Patel and Honore 2010; Patel 2014). PC1 and PC2 are necessary for the calcium response and nitric-oxide release in endothelial cells in response to blood flow (Nauli, Kawanabe et al. 2008). Extracellular flow-induced calcium influx leads to Ca2+ release from the endoplasmic reticulum via ryanodine and inositol triphosphate receptors, resulting in AMP release (Winyard and Jenkins 2011). Kidney epithelial cells respond to urinary flow in the lumen of renal tubules by bending of the PC. PC acts as a mechanosensory antenna in the tubule lining epithelial cells upon fluid shear stress generated by the fluid flow. This stimuli increases the membrane permeability to Ca2+. It has also been shown that different renal cells have additional cilia-independent mechanosensitive Ca2+ responses (Liu, Xu et al. 2003; Rodat-Despoix, Hao et al. 2013). The flow dependent calcium signaling is addressed further in the manuscript. Recently, Delling et al. elegantly showed that the PC is a specialized organelle with respect to intracellular ions by using a ratiometric Ca2+ sensor for individual cilia and patch-clamp methods (Delling, DeCaen et al. 2013). The study demonstrated that ciliary calcium channels, specifically the heteromeric PKD1L1-PKD2L1 channel, maintain a high Ca2+ concentration in the cilium with respect to cytoplasm in resting state, thus regulating a steady diffusion into the cell. The rapid calcium dilution to cytoplasm does not activate a Ca2+
25
release but positively regulates ciliary Hh trafficking via IFT25, which contains a unique Ca2+ binding site (Delling, DeCaen et al. 2013).
1.2.5. Fluid flow and Fluid Shear Stress
The mechanosensory role of PC is evolutionarily important for various organs. PC can sense different fluid movements such as urine in the kidney tubules, blood in the vascular system, bile in the hepatic biliary system, lacunocanalicular fluid in the bone and cartilage, digestive fluid in the pancreatic duct, nodal flow in the Hensin’s node, and cerebral spinal movement in the nervous system. The transduction of mechanical stimuli by the PC depends on different receptors and channels such as calcium channels polycystin 1 (PC1) and polycystin 2 (PC2) on the axoneme depending on the tissue and thus can induce various intracellular signalizations explained above. See Table 1 for an overview of cilia-dependent mechanical stress in different tissues. There are other mechanosensory cellular compartments in different tissues including microvilli, actin filaments in the kidney, glycocalyx, a membrane-bound filament structure found or caveolae, an invagination of the plasma membrane rich in lipids, receptors and channels including K+ channels, Cl- channels, and Ca2+ channels on the surface of endothelial cells. Tyrosine kinase receptor (TKR), G-protein-coupled receptors (GPCRs) and G-protein itself have been shown to be activated by fluid shear stress. Adhesion proteins (PECAM-1, and VE-cadherin) at sites of cell–cell and cell– matrix attachment are subjected to tension under shear stress. Cytoskeleton is responsible for cell shape regulation and can respond to mechanical forces that deform cells (Ando and Yamamoto 2013) (Figure 6).
26
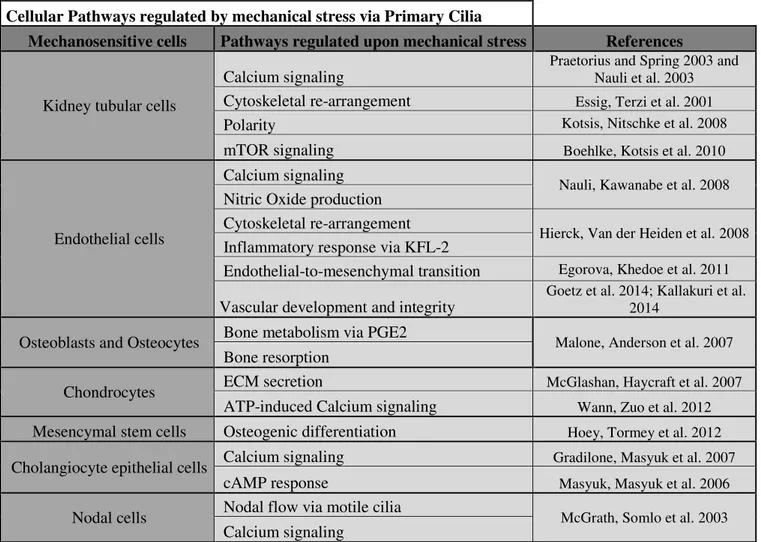
Cellular Pathways regulated by mechanical stress via Primary Cilia
Mechanosensitive cells Pathways regulated upon mechanical stress References
Kidney tubular cells
Calcium signaling
Praetorius and Spring 2003 and Nauli et al. 2003
Cytoskeletal re-arrangement Essig, Terzi et al. 2001
Polarity Kotsis, Nitschke et al. 2008
mTOR signaling Boehlke, Kotsis et al. 2010
Endothelial cells
Calcium signaling
Nauli, Kawanabe et al. 2008
Nitric Oxide production Cytoskeletal re-arrangement
Hierck, Van der Heiden et al. 2008
Inflammatory response via KFL-2
Endothelial-to-mesenchymal transition Egorova, Khedoe et al. 2011
Vascular development and integrity
Goetz et al. 2014; Kallakuri et al. 2014
Osteoblasts and Osteocytes Bone metabolism via PGE2 Malone, Anderson et al. 2007
Bone resorption
Chondrocytes ECM secretion McGlashan, Haycraft et al. 2007
ATP-induced Calcium signaling Wann, Zuo et al. 2012
Mesencymal stem cells Osteogenic differentiation Hoey, Tormey et al. 2012
Cholangiocyte epithelial cells Calcium signaling Gradilone, Masyuk et al. 2007
cAMP response Masyuk, Masyuk et al. 2006
Nodal cells Nodal flow via motile cilia McGrath, Somlo et al. 2003
Calcium signaling
27 Figure 6 : Mechano-sensing by the cell
Candidates for shear stress sensors on the plasma membranes are presented. Various types of ion channels including K+ channels, Cl- channels, and Ca2+ channels have been shown to be shear stress responsive.TKR, GPCRs, and G-protein itself have been shown to be activated by shear stress. Caveola are membrane microdomains containing a variety of receptors, ion channels, and signalling molecules are involved in shear-stress sensing. Adhesion proteins (PECAM-1, and VE-cadherin) at sites of cell–cell and cell–matrix attachment are subjected to tension under shear stress are such adhesion proteins. Cytoskeleton is responsible for cell shape regulation and can respond to mechanical forces that deform cells. Glycocalyx unfolds under flow conditions, and this conformational change affects the local concentrations and transport of ions, amino acids, and growth factors, or triggers signal transduction. Primary cilia is known to detect flow. Well known flow-responsive calcium channels on the cilium are PC1 and PC2. Shear stress induces an influx of extracellular Ca2+ via PC2. (Ando and Yamamoto 2013)
Endothelial cells cover the inner surface of blood vessels. They are under fluid shear stress (FSS) which is the drag force induced by the viscous blood acting on the vessel wall in parallel to the blood flow (Hierck, Van der Heiden et al. 2008). Endothelial cells are also subjected to cyclic stretch forces perpendicular to the shear forces caused by hydrostatic and blood pressures (Resnick, Yahav et al. 2003). In the vascular system fluid shear stress can vary from 10-40 dyn/cm2 in large arteries such as the aorta with a laminar pulsatile flow and to 1-6 dyn/cm2 on the walls of the veins. FSS is responsible for the homeostasis of the circulatory system and various phenotypes including angiogenesis, blood coagulation or vascular remodeling (Ando and Yamamoto 2013). The impairment of flow sensing can lead to various pathologies including hypertension and atherosclerosis (Davies, Mundel et al. 1995). The ability of endothelial cells to sense the mechanical stimuli depend on various proteins such as ion channels, G-protein receptors, adhesion molecules expressed on the plasma membrane or on supporting structures such as cytoskeleton and lipid bilayer
28
membrane or on microdomains such as caveolae and primary cilium (Davies, Mundel et al. 1995). The presence of PC in endothelial cells depends on the blood flow pattern and exposed shear stress levels in the circulatory system. For example, most ciliated endothelial cells are present in inner curvature of aorta or various arteries where the flow can be disturbed and observed to be oscillatory whereas the endothelial cells on the areas exposed to high shear stress such as on the outer curvature of the aorta are deciliated (Egorova, Van der Heiden et al. (2012). PC is not the sole mechano-sensor in endothelial cells however bending of the cilium is vital for the influx of extracellular calcium and cytoskeletal re-arrangement by appearance of stress actin fibers with re-orientation to the shear axis and also for the regulation of zinc-finger transcription factor Kruppel-like factor-2 (KLF-2) (Hierck, Van der Heiden et al. 2008). The cytosolic Ca2+ increase is followed by nitric oxide production which is important for vascular contractility. Endothelial cells mutated for PKD1 or Tg737 (IFT88) failed to induce this response (Nauli, Kawanabe et al. 2008). The regulation of KLF-2 expression by PC is important in atherosclerosis as during low oscillatory, “athero-prone”, flow in ciliated areas of the circulatory system, KLF-2 expression is suppressed and an inflammatory response is induced which may result in plaque formation (Hierck, Van der Heiden et al. 2008). In areas exposed to high shear stress, KLF-2 expression is induced and cells are observed to lose their cilia which can also correspond to endocardial cushions in the embryo which undergo TGF-β mediated endothelial-to-mesenchymal transition (EndoMT) (Egorova, Khedoe et al. 2011). Two recent studies showed another role of PC in flow sensing during vascular development and integrity in zebrafish. These studies showed that PC was involved during early embryonic stages, during which a low blood flow is established. PC is observed to mediate vascular morphogenesis via PC2-mediated calcium signaling upon flow induced bending (Goetz, Steed et al. 2014). It is also shown that the presence of PC is essential for vascular integrity via the Hh pathway since the cilia impairment increased the risk of intracranial hemorrhage (Kallakuri, Yu et al. 2014).
Lacuno-canalicular fluid in the bone and cartilage creates a dynamic flow that is sensed by the primary cilium of osteocytes in the bone matrix and osteoblasts. The nature of the mechanical stress in the bone, as well as in the vascular system, is still under investigation (Fritton and Weinbaum 2009). PC is required for osteogenesis and bone resorption in response to mechanical stress (Weinbaum, Duan et al. 2011). Osteoblasts up-regulate osteopontin expression, a bone matrix protein along with cytokine prostaglandin E2 (PGE2)
29
resorption regulation of osteocytes in the bone is a calcium independent homeostatic response regulated by the PC (Malone, Anderson et al. 2007). As for the cartilage cells, it was previously shown a role of PC in the chondrocyte differentiation and hypertrophy (McGlashan, Haycraft et al. 2007). A mechano-sensory role of chondrocytes is also in question during the mechanical physiological joint loading. Compressive loading in chondrocytes regulates extracellular matrix secretion and the ATP induced Ca2+ signaling via purine receptors in a cilia dependent manner (Wann, Zuo et al. 2012). Mechanical loading induced by fluid shear stress stimulates mesenchymal stem cell differentiation into bone forming osteoblasts in a PC dependent manner via osteogenic gene expression regulation (Hoey, Tormey et al. 2012).
The role of PC in the bile duct is important as the liver cysts observed in ADPKD patients originate from the epithelial cells lining the bile duct, cholangiocytes. It is reported that luminal bile flow induces Ca2+ signaling response and cAMP decrease in ciliary dependent manner via PC1 and PC2 in vitro (Masyuk, Masyuk et al. 2006). The osmotic stress caused by luminal flow regulates absorption of solutes and water secretion by cholangiocytes via TRPV4 channel on the PC leading to the secretion of bicarbonate thus ductal bile formation (Gradilone, Masyuk et al. 2007).
It is known that primary cilia, known as “nodal” cilia is vital for left-right body axis determination during embryonic development. McGrath et al. showed that ciliary PC2 is responsible for the mechansosensation of the nodal flow created by the motile cilia across the node and creating an intracellular Ca2+ signaling (McGrath, Somlo et al. 2003).
The role of primary cilia and its mechanosensory role in the kidney will be detailed later in the manuscript (see 2.6.1.Physiology of the kidney: an introduction).
In the case of motile ciliated tissues, including ependymal cilia, it is shown that the organelle can have a role in flow direction rather than flow sensing. It is reported that motile cilia of the ependymal cells are important for directional regulation of cerebrospinal fluid flow and neuronal migration (Sawamoto, Wichterle et al. 2006).
30
2.
Autophagy
2.1. Role of Autophagy
The word “autophagy” is derived from the Greek words “auto” and “phagy” meaning “self-eating”. The term “autophagy” is first introduced in 1963 by the biochemist Christian de Duve who received a Nobel Prize in Physiology or Medicine in 1974 for his discovery and work on lysosome (Klionsky 2007). Autophagy is an evolutionarily conserved mechanism for eukaryotes and refers to catabolic processes and turnover of cellular contents such as intracellular misfolded or long-lived proteins, damaged organelles and invading microorganisms. It is characterized by the formation of double-membrane vesicles, “autophagosomes”, which sequester cytoplasmic material. However, autophagy is more than just a degradative process, it is a recycling system that responds to various intra- and extra-cellular stresses by generating metabolites as sources of energy or building blocks for synthesis of new macro-molecules such as amino acids, free fatty acids and sugars (Yang and Klionsky 2010; Boya, Reggiori et al. 2013). The leading role of autophagy is maintaining the energy homeostasis. Stimuli that activate autophagy include for example, nutrient deprivation and other nutritional challenges as well as growth factor withdrawal, oxidative stress, protein accumulation, infection. Autophagy plays an essential role in multitude of physiological processes ranging from adaptation to starvation, cell differentiation and development, as well as tumor suppression, innate and adaptive immunity, lifespan extension, and cell death. Autophagy mechanism is deregulated in various human pathologies, for example cancer, neurodegeneration, obesity, type-2 diabetes, heart and liver diseases, infectious and immune diseases and even physiologic stages such as aging (Rubinsztein, Codogno et al. 2012; Choi, Ryter et al. 2013).
31
2.2. Different Types of Autophagy
There are three major types of autophagy described in mammals: macroautophagy, chaperone-mediated autophagy (CMA) microautophagy (Figure 7) (Boya, Reggiori et al. 2013).
2.2.1. Macroautophagy
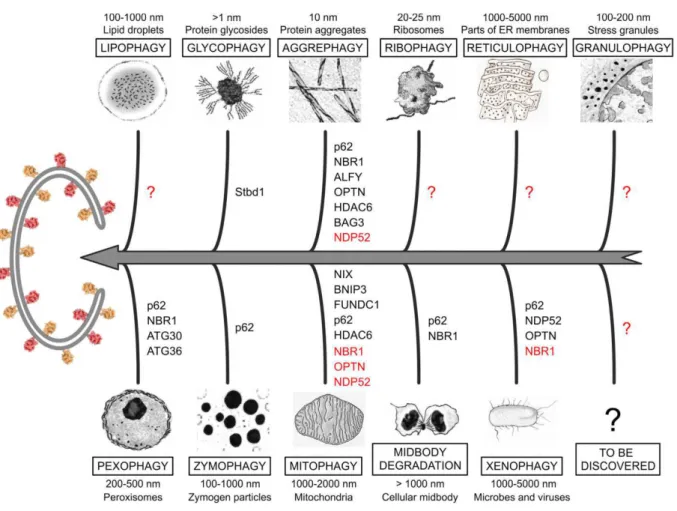
Macroautophagy, usually referred to as autophagy, is the process of bulk degradation of cytoplasmic material. It is characterized by de novo formation of the isolation membrane, called phagophore, which sequesters cytoplasmic content or organelles and sealing of the double membrane vesicle “autophagosome” that will later fuse with lysosomes. This process is regulated by a group of proteins described as autophagy-related (Atg) proteins that are conserved in eukaryotic cells (Rubinsztein, Shpilka et al. 2012). The discovery of Atg genes in yeast was revolutionary for the field and lead to our understanding of the molecular mechanism of autophagy (Tsukada and Ohsumi 1993; Thumm, Egner et al. 1994; Harding, Morano et al. 1995). The formation of autophagosomes as well as the membrane sources of this double-membrane structures are still not 100% characterized. Autophagy is a bulk process, but, in many cases, it can be highly selective of cellular structures, including peroxisomes (pexophagy), mitochondria (mitophagy), endoplasmic reticulum (reticulophagy), ribosomes (ribophagy), midbody, micro-nuclei and part of the nuclei (nucleophagy), lipid
droplets (lipophagy), aggregate-prone proteins (aggrephagy), secretory granules
(zymophagy), damaged lysosomes (lysophagy), and microorganisms (xenophagy) (Rogov, Dotsch et al. 2014).The molecular aspect of macroautophagy will be focused further in detail in the manuscript.
2.2.2. Chaperone-mediated Autophagy
Chaperone-mediated autophagy (CMA) is a selective form of autophagy where the substrates bind to a cytosolic chaperone, a heat shock-cognate protein of 70 kDa (Hsc70), and then to a lysosomal protein, the lysosomal associated membrane protein type 2A (LAMP‐2A). The recognition of the CMA substrates by Hsc70 requires a motif related to the pentapeptide KFERQ (Dice 1990) after which they are directly targeted to lysosomal surface and interact with the cytoplasmic tail of LAMP-2A. This is followed by the unfolding of the substrate
32
protein in order to translocate across the lysosomal membrane and to be degraded (Cuervo and Wong 2014). The substrates of CMA are soluble proteins but not organelles, because they are not engulfed by the lysosome but rather transported across the lysosomal membrane. This translocation of substrates requires a multi-protein complex formed by the association of LAMP-2A monomers at the lysosomal membrane. The stability of the LAMP-2A is maintained via Hsp90 at the luminal membrane. Following substrate translocation, LAMP-2A complex disassembles for others to bind. This process can be regulated in a GTP-dependent manner through GTP-bound EIF1α- GFAP (Glial fibrillary acidic protein) interaction during the disassembly of the multimeric complex (Bandyopadhyay, Sridhar et al. 2010).
CMA contributes to quality control of proteins and exists at basal levels however it is maximally induced by stress conditions similar to macroautophagy, such as starvation, oxidative stress or exposure to various toxic compounds (Massey, Zhang et al. 2006; Cuervo and Wong 2014). However in contrast to rapid activation of macroautophagy upon starvation, CMA is gradually activated upon 10 hours of starvation and persists up to 3 days which contributes to the selective degradation of non-essential proteins to generate building blocks of essential proteins’ synthesis (Massey, Kaushik et al. 2006). Both the perturbation and increase of CMA activity can have roles in various pathologies. The reduction of CMA contributes primarily to neurodegenerative diseases such as Parkinson’s and Alzheimer’s diseases; whereas in certain cancer cases increased CMA is observed, contributing to rapid cell growth by sustaining enhanced glycolysis. Another important role of CMA is defined during aging as its activity is significantly decreased over time along with total protein degradation rate reduction in all organisms (Cuervo and Wong 2014). Recent data showed a previously unknown role for CMA via a liver specific conditional LAMP-2A knock-out mouse model. CMA impaired mice showed altered hepatic carbohydrate and lipid metabolism in aging mice suggesting that CMA loss has a negative role on the energetic balance in aging organisms (Schneider, Suh et al. 2014).
33 Figure 7 : The different types of autophagy
(A)Macroautophagy is characterized by the sequestration of cytosolic material in double-membrane vesicles called autophagosomes. (B) During chaperone-mediated autophagy, proteins with KFERQ motifs are recognized by the Hsc70 chaperone. Later they associates with the integral lysosome membrane protein LAMP-2A. (C) Microautophagy entails the recruitment of targeted components in proximity with the lysosomal membrane, which subsequently invaginates and pinches off. (Boya, Reggiori et al. 2013).
34
2.2.3. Microautophagy
In the case of microautophagy, the cytoplasmic content is directly engulfed by the lysosome by the invagination of the lysosomal membrane itself. This invagination of lysosomes regulate the lysosomal size by consumption of the membrane formed during macroautophagy (Li, Li et al. 2012). The accompanying membrane structures can function in closure of the cargo in an Atg protein dependent manner. The vesicles containing the autophagic substrates pinch off into the lysosomal lumen to be degraded by lysosomal hydrolases (Cuervo and Wong 2014). Both macro- and micro-autophagy can mediate the degradation of various organelles including peroxisomes, mitochondrion, lipid droplets and mucleus in yeast, however the elimination of various organelles by microautophagy in mammals remain to be investigated (Okamoto 2014). Microautophagy can engulf multivesicular bodies, therefore, it can participate to renewal of cell membrane proteins in endosomes. It has also been shown to participate in glycogen delivery into the lysosome serving as a delivery route (Li, Li et al. 2012).
2.3. Autophagosome Machinery and Biogenesis
Macroautophagy, referred as autophagy from now on, depends on a tight and complex molecular regulation that involves more than 30 Atg proteins that were firstly identified in yeast. Autophagy related proteins involved in the formation and maturation of autophagosomes are almost entirely conserved in all eukaryotes (Mizushima, Yoshimori et al. 2011). Autophagy is characterized by a dynamic membrane re-organization which starts with the formation of a cup-shaped double-membraned sac called the isolation membrane (also referred to as phagophore) in the cytoplasm which expands in size and later seals as the double-membraned autophagosome. Autophagosome size can differ from 0.3-0.9 µm in yeast (Baba, Osumi et al. 1997) and 0.5-1.5 µm in mammals (Mizushima, Ohsumi et al. 2002). Autophagosomes later fuse with lysosomes (with vacuoles in yeast), forming the autolysosomes in which the intracellular content and the inner membrane of the autophagosome are degraded by lysosomal hydrolases. Formation of autophagosome and autolysosomes are highly dynamic. The two processes together can take 7-9 minutes in yeast (Geng, Baba et al. 2008) and autophagosome formation can take 5-10 minutes in mammals (Fujita, Hayashi-Nishino et al. 2008).
35
There are three important stages in autophagic machinery: 1. Initiation and nucleation (formation of the phagophore); 2.Vesicle elongation (growth and closure of autophagosome); 3.Maturation into autolysosomes. These stages depend on different core Atg protein units: Atg1/ULK complex, the class III phosphatidylinositol 3-kinase (PI3K) (Beclin-1-PtdIns3KC3-Atg14L) complex, the Atg2-Atg18/ WIPI complex, the Atg12-Atg5-Atg16L1 conjugation system, the Atg8/LC3 conjugation system, and Atg9 vesicles. For the relevance of the manuscript, the main focus will be on mammalian autophagic machinery (Mizushima, Yoshimori et al. 2011) (Figure 8).
2.3.1. Initiation
ULK complex
ULK1, mammalian orthologue of yeast Atg1, is considered to have the major role in autophagy (Chan, Kir et al. 2007). ULK complex is the most upstream module of autophagic machinery. The multimeric complex includes unc-51-like kinase 1 (ULK1), focal adhesion kinase family interacting protein of 200kD (FIP200), Atg13 and Atg101 (Itakura and Mizushima 2010). The ULK1-mAtg13-FIP200-Atg101 complex is constitutively formed in the cytosol independent of nutrient conditions. Under nutrient rich conditions ULK1 and Atg13 is phosphorylated and the complex is inactivated by mammalian target of rapamycin complex 1 (mTORC1). Upon autophagy induction, this stable complex translocate to the autophagosome assembly site at the ER membrane, where ULK1 is no longer phosphorylated by mTORC1 (Itakura and Mizushima 2010) (Figure 9). ULK1, a Ser/Thr kinase is known to be required for autophagy induction and recruitment of the ULK complex on the phagophore membrane in mammals (Hara, Takamura et al. 2008). ULK1 is known to phosphorylate Atg13 and FIP200 as well as other proteins such as AMBRA1 or a non autophagic zipper-interacting protein kinase ZIPK protein however the relevance of the substrates of this kinase remains to be further elucidated (Mizushima, Yoshimori et al. 2011). Atg101 interacts with and stabilizes Atg13. ULK1 and ULK2 are mammalian orthologues closely related to yeast Atg1 protein and are shown to be functionally redundant in vivo and to form complexes with Atg13 and FIP200 in a compensatory manner in response to nutrient deprivation (McAlpine, Williamson et al. 2013).
36 Figure 8: The Autophagic pathway
There are two important stages in autophagy pathway: (i) autophagosome formation, which includes the initiation/nucleation (formation of the phagophore) and elongation/closure, and (ii) autophagosome fusion with the lysosome. Autophagy-related (ATG) proteins involved in the formation of autophagosome and a non-exhaustive list of the protein families involved in the maturation/fusion step are shown in italics.
The class III PI3K complex
There are at least three class III phosphatidylinositol kinase (PI3K) complexes in mammalian cells which have different functions in the cell. The major class III PI3K complex that is required for the autophagosome formation include a PI3 kinase the vacuolar protein sorting 34 (Vps34), Vps15, Beclin1, Atg14 (L) (also called Barkor) and activating molecule in Beclin1-regulated autophagy (AMBRA1) (Itakura and Mizushima 2010).