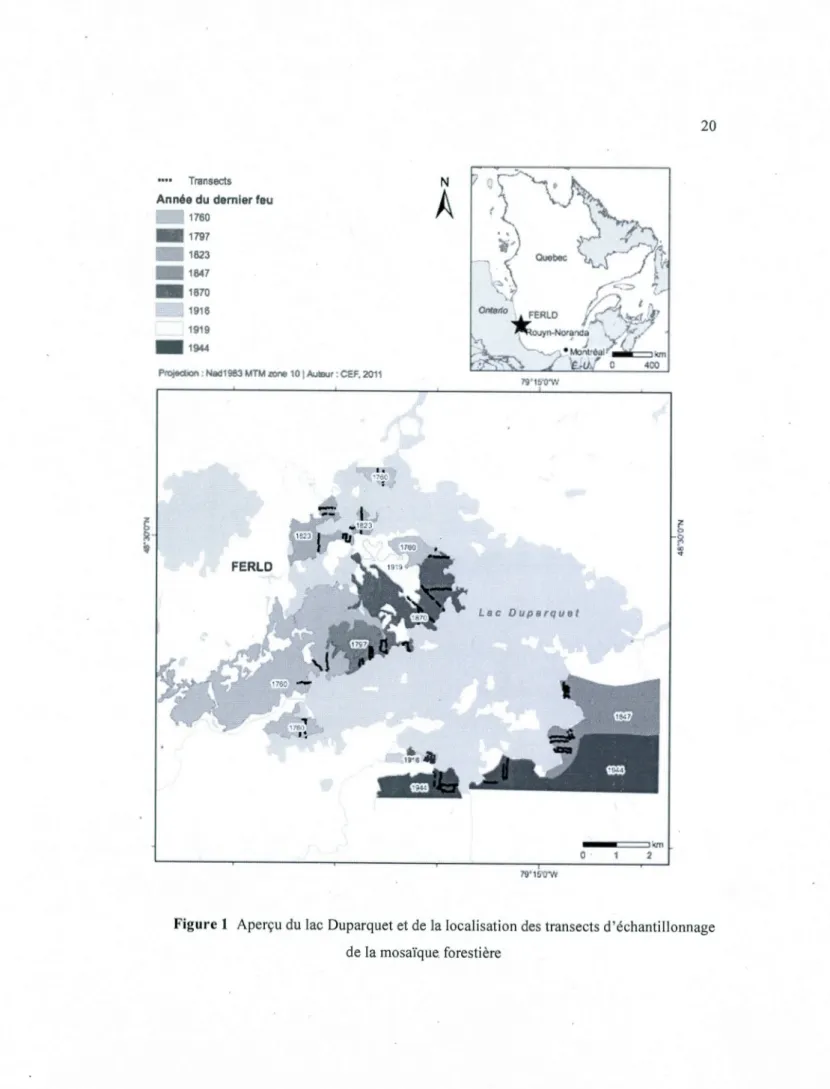
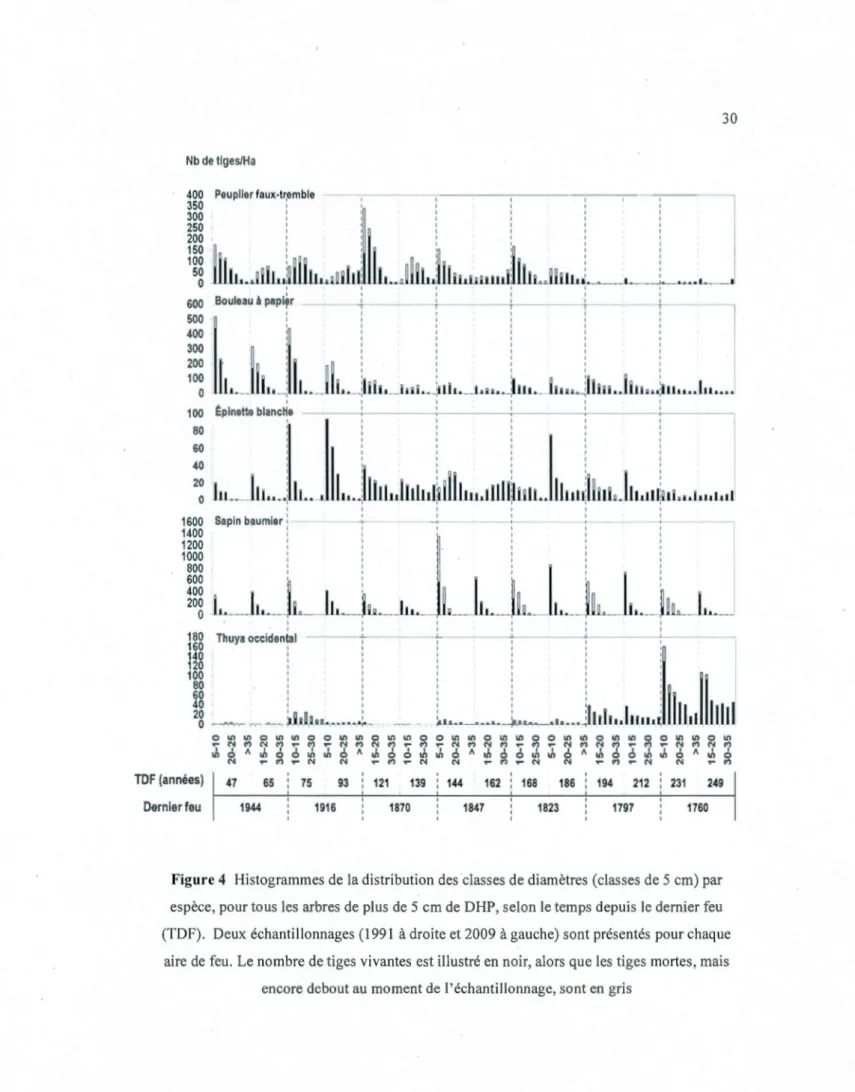
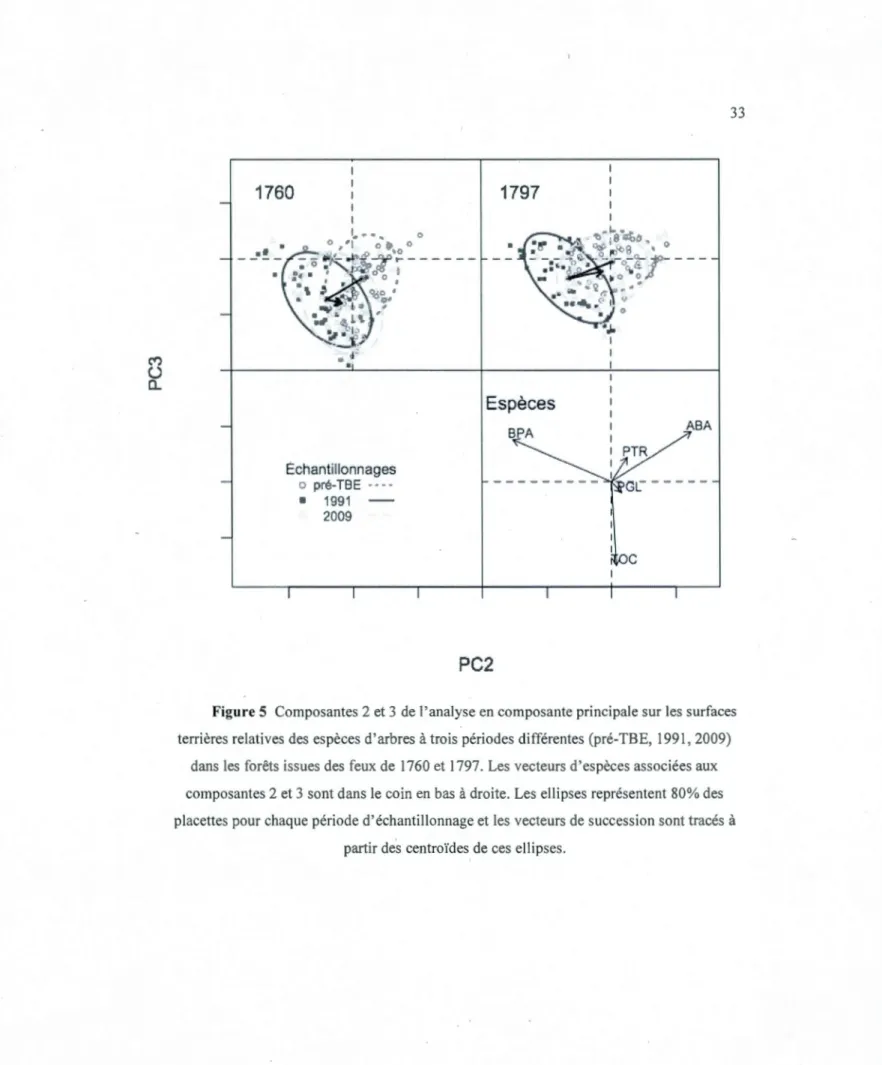
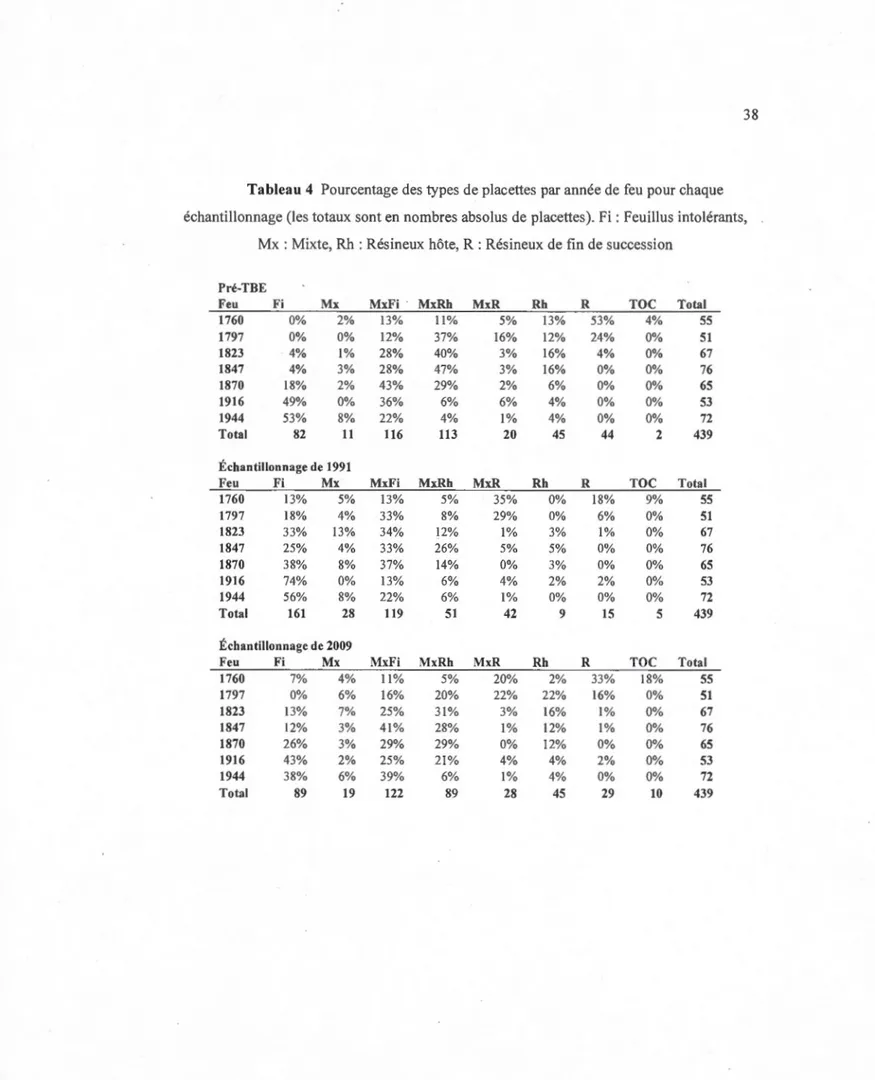
Dix-huit ans de changements in situ du couvert forestier dans une chronoséquence de 249 ans de la forêt boréale mixte en Abitibi
70
0
0
Texte intégral
Figure



+7



Documents relatifs