Etude physiologique du syndrome d’apnées à composante centrale de l’adulte, hors insuffisance cardiaque à fraction d’éjection systolique altérée
130
0
0
Texte intégral
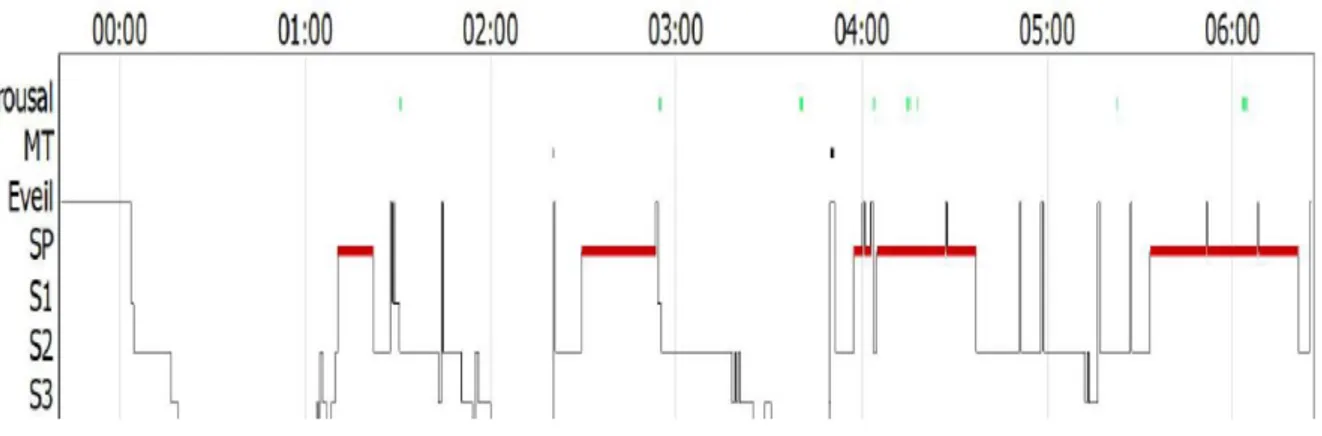
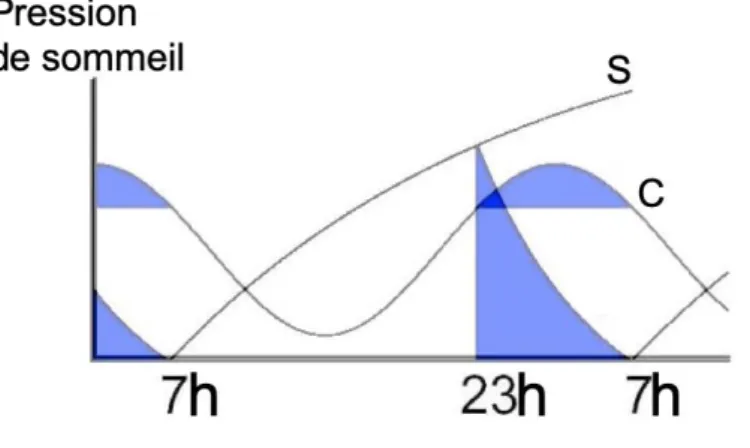
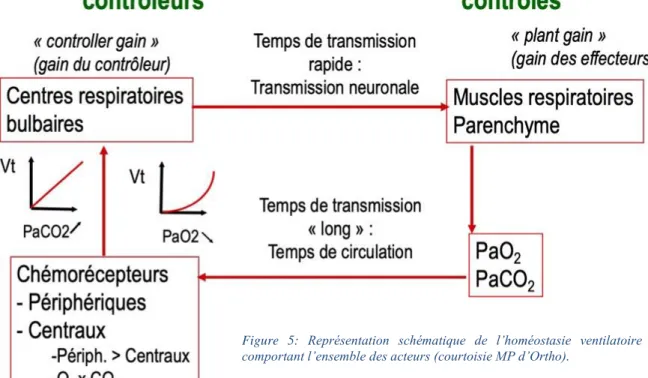
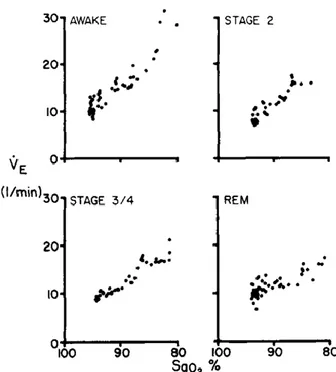
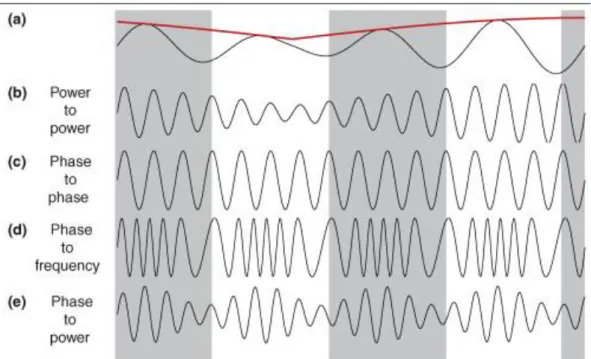
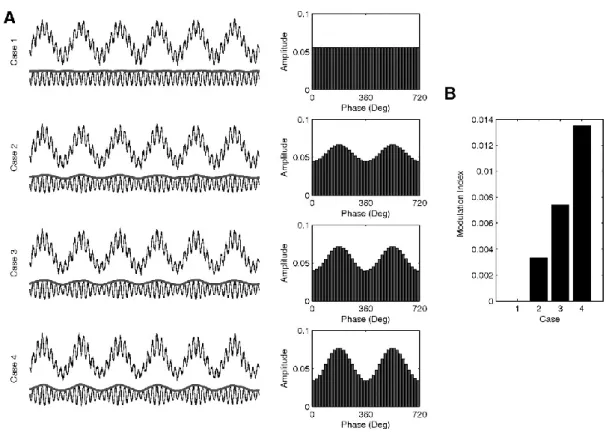
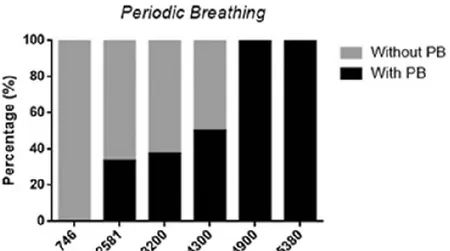
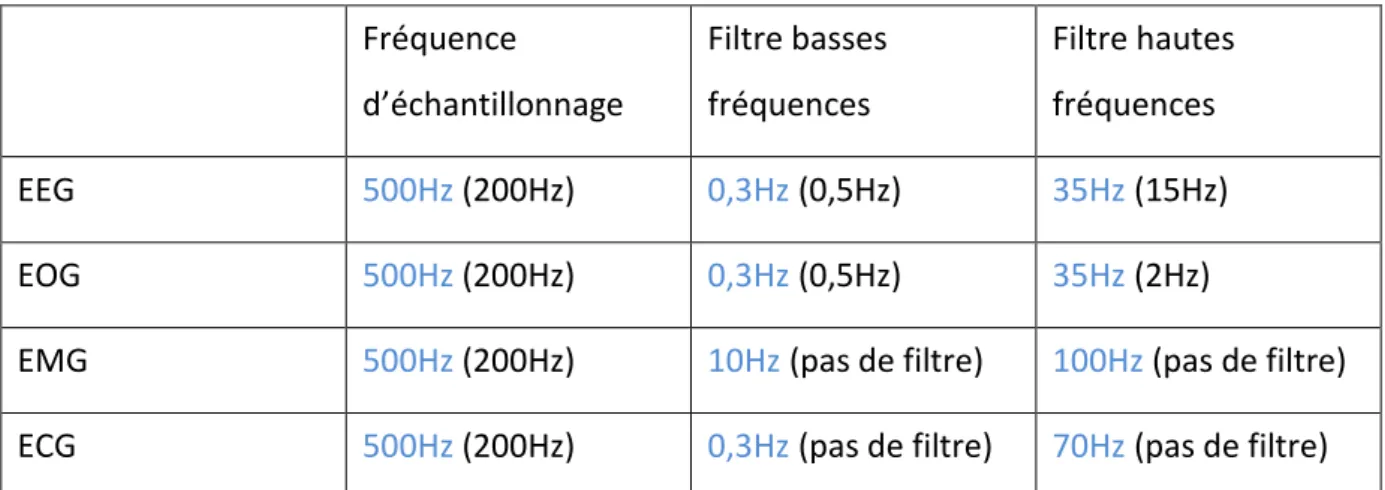
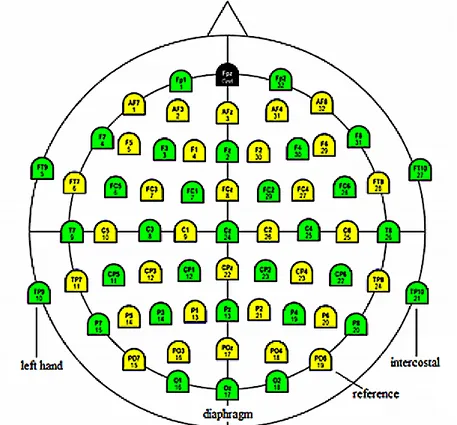
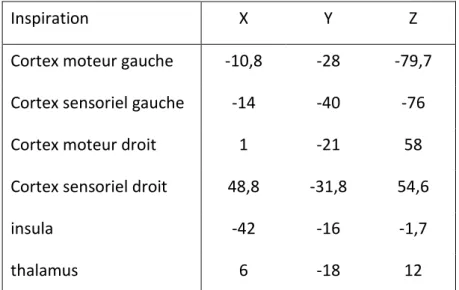
Figure
+7
Documents relatifs