0
MASTER 1 Biodiversité, Ecologie, Evolution
Université de Lille, Sciences et Technologies (Lille 1)
Année 2017-2018
Rôle des continuités écologiques sur la structuration des
communautés de plantes prairiales
Ophélie Devos
L. Uroy, A. Alignier, A. Ernoult et C. Mony
UMR INRA 0980 BAGAP, Rennes
Equipe paysaclim de l’UMR 6553 ECOBIO, Université de
Rennes 1
2
Table des matières
Introduction ... 1
Matériels et méthodes ... 4
- Zone d’étude ... 4
- Site d’échantillonnage ... 6
- Métriques paysagères ... 7
- Méthode d’échantillonnage de la végétation ... 8
- Indices de diversité ... 9
- Analyses statistiques ... 9
Résultats ... 10
- Caractéristiques des communautés végétales étudiées ... 10
- Résultats des régressions multiples ... 11
- Résultats de l’analyse canonique des correspondances ... 14
Discussion ... 15
- Effet de la proportion d’habitat sur la diversité et la composition spécifique des prairies ... 15
- Effet de connectivité sur la diversité spécifique ... 15
- Effet de la taille de la parcelle sur la diversité spécifique ... 16
Conclusion et perspectives ... 17
Bibliographie... 17
Résumé ... 21
Annexe ... 20
1
Introduction
Depuis la 2ème moitié du 20è siècle, l’intensification des activités agricoles a provoqué une accélération de la fragmentation des habitats au sein des paysages agricoles induisant une perte de biodiversité. En Bretagne, on observe notamment une augmentation de la surface des cultures, un agrandissement des parcelles et une diminution du nombre de haies. Cette fragmentation se caractérise par la réduction de la surface d’habitats disponible et par l’isolement des tâches d’habitat résiduelles (Fahrig 2003 ; Fisher & Lindemeyer, 2007). Elle occasionne un fonctionnement en métapopulation des espèces (Hanski, 1994). Une métapopulation se définit comme un ensemble de populations locales séparées spatialement mais reliées entre elles par les flux d’individus. En réduisant la taille des populations, la fragmentation provoque l’augmentation de leur probabilité d’extinction, de par leur sensibilité importante aux facteurs stochastiques (stochasticité démographique, environnementale, naturelle et génétique) (Fankham et al.,2002). Ainsi, ces populations locales ne se maintiennent que par la dispersion des individus entre les différentes tâches d’habitat (Fahrig & Merriam, 1985 ; Browne & Bowers, 2004).
La dispersion des individus dépend de la connectivité du paysage (Kadoya, 2009), qui se définit comme le degré avec lequel un paysage facilite ou entrave le mouvement des individus ou des propagules (dans le cas des plantes) entre les différentes tâches d’habitat (Taylor et al., 1993). Lorsque le concept de connectivité prend en considération les liens structuraux entre tâches d’habitat, il correspond à la « connectivité structurale » ; quand il inclut l’aspect comportemental des espèces vivant dans ces tâches d’habitat il est nommé « connectivité fonctionnelle ».
Les continuités écologiques sont l’ensemble des connexions entre tâches d’habitat favorable (Beier & Noss, 1998 ; Chetkiewicz et al., 2006). Elles augmentent la connectivité paysagère en permettant la dispersion des individus entre populations. On connait principalement les corridors écologiques qui sont des éléments linéaires reliant deux taches d’habitat, mais les continuités regroupent également les éléments non-linéaires et discontinus (en patch). En permettant aux individus de disperser entre tâches d’habitat, les continuités écologiques luttent ainsi contre les effets néfastes de la fragmentation et contribuent au maintien de la biodiversité (Baudry & Burel, 1999).
Par ailleurs, l’augmentation de la taille des patchs d’habitats favorables au sein de la matrice paysagère, en garantissant le maintien d’un minimum d’habitat favorable pour la persistance des individus (mean patch size requirement, Fahrig 2003 ; Fahrig et al., 2011) ainsi qu’en facilitant les échanges d’individus ou de propagules entre ces tâches d’habitats, favorisera
2 le maintien des populations. De même, une diminution de la distance entre ces tâches (aussi appelé « isolement »), en augmentant la connectivité entre les tâches favorisera la dispersion des individus et donc le maintien des populations (Fahrig 2003).
Les prairies, qui représentent 45% de la surface agricole utile (SAU) en France (Huyghe, 2009), montrent un déclin ces dernières années. En Bretagne, on distingue les prairies de courte durée des prairies de longue durée. Les prairies de longue durée sont des prairies n’ayant pas été retournée pendant au moins 5 ans (Peyraud et al., 2012). Elles peuvent avoir été ressemées naturellement ou cultivées. On recense une perte de 30% des surfaces en prairies de longue durée en Europe, dont 33% pour la France, entre 1967 et 2007 (Peyraud et al., 2012).
Pourtant, ces prairies sont d’un grand intérêt : en plus d’héberger une grande biodiversité, elles constituent un agroécosystème multifonctionnel qui apporte à l’homme de nombreux services écosystémiques. Les prairies de longue durée assurent une production fourragère pour l’alimentation du bétail mais elles occupent également un rôle dans la préservation de l’environnement en participant au maintien des cycles bio-géo-chimiques (Huyghe, 2009). Quant aux services écosystémiques rendus à l’homme, les prairies de longue durée contribuent à une eau de meilleure qualité, de par la diminution des traitements phytosanitaires appliqués en comparaison avec ceux appliqués aux cultures. Elles ont également la particularité d’être des puits de carbone, de limiter l’érosion des sols (Chisci & Zanchi, 1981) et de contribuer au maintien la biodiversité des milieux agricoles (Huyghe, 2009 ; Ockinger & Smith, 2007).
D’un point de vue floristique, on retrouve majoritairement les espèces semées dans les prairies jeunes. Après quelques années d’implantation, la colonisation par des adventices de cultures ou « mauvaises herbes » est de plus en plus importante. Le cortège floristique colonisant la prairie dépend ainsi de nombreux facteurs. A l’échelle du paysage, la taille des parcelles, la connectivité entre tâches d’habitats influencent la richesse et la composition des prairies. Tandis qu’à l’échelle locale, c’est-à-dire à l’échelle de la parcelle, les paramètres physico-chimiques (hygrométrie, pH,…), le mode de gestion par l’agriculteur (pâturage ou fauche), la qualité et la quantité d’amendement sont autant de facteurs susceptibles d’influencer la structure et la composition du cortège floristique (Baker & Berendse 1999 ; Olff, Han & Ritchie 1998).
Cette étude s’inscrit dans la thèse « Rôle de la connectivité passée sur la structuration des communautés végétales en paysage agricole : Prise en compte de la complémentarité entre
3 trames » de Léa Uroy dans le cadre du projet BISCO (« BIodiversité et Services écosystémiques associés aux COmplémentarités entre continuités écologiques boisées et cultivées dans les paysages agroforestiers »). Elle contribue également au projet européen Biodiversa). Woodnet (signifiant « corridors boisés » faisant référence aux réseaux de haie). Le projet Woodnet a pour but de fournir des outils novateurs et spatialement explicites pour l'analyse de la connectivité sur une gamme de paysages allant des forêts et des maquis aux paysages agricoles où les éléments de végétation ligneuse jouent un rôle clé pour la conservation de la biodiversité (https://www.biodiversa.org/1026).
Dans cette étude, nous nous intéressons à l’impact du paysage, et notamment au rôle des continuités écologiques, sur la structuration des communautés de plantes prairiales. On émet les hypothèses suivantes :
(1) une plus grande proportion d’habitat prairial dans le paysage augmente la richesse spécifique (nombre d’espèces), l’équitabilité et la diversité de Shannon au sein des prairies ;
(2) une plus grande connectivité entre ces prairies augmente la richesse, l’équitabilité et la diversité de Shannon de ces prairies ;
(3) une plus grande proportion d’habitat prairial dans le paysage et une plus grande connectivité entre pariries influence la composition floristique de ces prairies.
Par ailleurs, nous faisons l’hypothèse que l’effet du paysage peut être différent en fonction de la gestion (pâturage ou fauche) des prairies.
4
Matériels et méthodes
- Zone d’étude
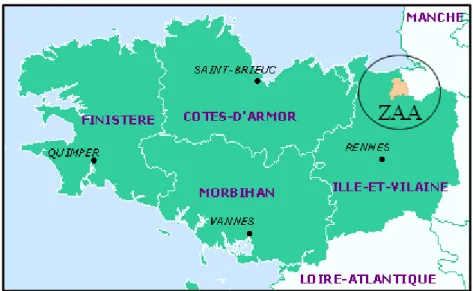
L’étude a été réalisée dans la Zone Atelier Armorique (ZAA), sur le site de Pleine-Fougères. La ZAA se situe au Nord-Est de l’Ille-et-Vilaine et au Sud de la baie du Mont-Saint-Michel (Fig.1). Sur une superficie de 130 km², 69 % des terres sont à vocations agricoles, et sont gérés par des systèmes de polycultures-élevages. Les prairies représentent 34 % de la ZAA. Cette zone d’étude présente un gradient de paysages agricoles caractérisé par une augmentation des surfaces cultivées, une diminution du réseau de haies ainsi qu’une augmentation de la taille des parcelles, du Sud vers le Nord (Fig.2).
Figure 1 : Localisation de la Zone Atelier Armorique, site de Pleine-Fougères sur la carte des départements de la Bretagne.
5 a) Paysage ouvert avec parcelles de grandes tailles et bocage lâche
b) Paysage de bocage intermédiaire
c) Paysage de bocage dense avec des parcelles de petite taille
Figure 2 : Photographies aériennes (Google Earth) illustrant le gradient de paysages agricoles retrouvé sur la Zone Atelier Armorique du Nord (a) vers le Sud (c)
6 - Site d’échantillonnage
37 prairies de longue durée ont été sélectionnées grâce à un Système d’Information Géographique (logiciel ArcGis), à partir de cartographies d’occupation du sol établies depuis des photographies aériennes de l’année 2016. Les prairies ont été sélectionnées avec un espacement minimal de 200 mètres entre elles afin de limiter leur autocorrélation spatiale (Fig. 3).
L’ensemble des prairies échantillonnées ont été standardisées selon de multiples critères (âge, précédent cultural, surface, semis, mode de gestion) afin de réduire l’impact des facteurs locaux. Les prairies sélectionnées étaient âgées de 6 à 9 ans, établies à la suite d’anciennes cultures et de tailles relativement équivalentes (0,94 ± 0,53 ha). Les prairies échantillonnées étaient semées soit (i) en mélange ray-grass (Lolium perenne) et/ou trèfle blanc (Trifolium
repens) ou (ii) en ray-grass italien (Lolium multiflorum) ou (iii) en fétuque (Festuca arundinacea). Le mode de gestion était de type pâturage ou fauche, avec amendement
occasionnel. Les informations sur le semis, l’amendement et le mode de gestion ont été recueillies par prospection auprès des propriétaires et exploitants des parcelles.
7 Figure 3 : Cartographie des 37 prairies étudiées et leur buffer de 200 mètres sur le parcellaire de la
Zone Atelier Armorique, site de Pleine-Fougères.
- Métriques paysagères
La matrice paysagère autour des prairies a été caractérisée à l’aide d’indices paysagers calculés avec le logiciel FRAGSTAT dans des buffers de 200 mètres de rayon autour de chaque prairie (Fig.3). La taille de ce buffer (200m) correspond à l’échelle de réponse des communautés adventices en en milieu agricole (Gaba et al., 2010).
La proportion d’habitat prairial au sein de chaque buffer a été calculée à partir de l’indice PLAND (Percentage of Landscape ; Tab.1). Il est exprimé en pourcentage et varie de 0 à 100%.
8 La connectivité a été caractérisée par l’indice ENN_MN (Euclidean Nearest-Neighbor Distance ; Tab.1), qui mesure la distance moyenne entre tâchesde même type. Ici, nous avons considéré les distances entre tâches de prairies. Plus la distance moyenne entre tâches diminue, plus la connectivité entre tâches de prairies augmente. Ainsi, un indice ENN_MN faible correspond à une forte connectivité.
Tableau 1 : Equations des indices paysagers calculés avec le logiciel FRAGSTAT
- Méthode d’échantillonnage de la végétation
Un échantillonnage exhaustif de la flore a été réalisé entre le 24 avril et le 15 mai 2018. Pour cela, 10 quadrats de 2m x 2m (Annexe 1) ont été positionnés selon un maillage régulier dans chacune des prairies. La flore a été déterminée in situ et par prélèvement pour les espèces les plus difficiles à identifier. Pour chaque espèce rencontrée, un pourcentage de recouvrement a été attribué. Afin d’éviter un effet de lisière, aucun quadrat n’est placé à moins de 5 mètres du bord de la parcelle (Fig.4).
Figure 4 : Schéma illustratif de la méthode d’échantillonnage. Les 10 quadrats de 2mx2m sont situés à plus de 5 mètres du bord de la parcelle, répartis en un maillage régulier.
𝐸𝑁𝑁_𝑀𝑁 =∑ ℎ𝑖𝑗
𝑛 𝑖=0
𝑁
Hij = distance (m) du patch ij au patch de même type le plus proche, basé sur la distance d’un bord à l’autre, calculé de centre à centre des cellules
N = Nombre de paires de patch
Pi = proportion du paysage occupé par l’habitat de type (classe) i Aij = surface (m²) du patch ij
A = surface totale du paysage (m²)
𝑃𝐿𝐴𝑁𝐷 = 𝑃𝑖 = ∑ 𝑎𝑖𝑗
𝑛 𝑗=1
9 - Indices de diversité
Pour caractériser les communautés végétales, nous avons calculé trois indices de diversité : (i) la richesse S correspondant au nombre total d’espèces (ii) l’indice de diversité spécifique de Shannon-Wiener H’ (Shannon & Weaver, 1949), caractérisant l’hétérogénéité des communautés. Cet indice varie entre 0 et log S. Une valeur proche de 0 de cet indice signifie que la communauté est dominée par une seule espèce. (iii) l’indice d’équitabilité de Pielou J’ (Pielou, 1966), qui varie de 0 (dominance d’une espèce) à 1 (équi-répartition des espèces).
Ces indices de diversité ont été calculés de deux façons différentes :
- chaque indice a été calculé pour chaque quadrat. La moyenne sur l’ensemble des quadrats a ensuite été réalisée par prairie. On parlera de l’échelle « alpha ».
- chaque indice a été calculé directement à l’échelle de la parcelle. La richesse correspond au nombre total d’espèces trouvé au sein de la prairie. On parlera de l’échelle « gamma ».
Enfin, chaque plante a été catégorisée selon son caractère « spécialiste » ou « généraliste » au sein de l’habitat prairial d’après une liste pré-établie par un panel de 5 experts en écologie. Les mêmes indices de diversité (Ss, H’s et J’s) ont été calculés uniquement pour la communauté des espèces spécialistes des prairies.
- Analyses statistiques
Pour tester l’influence de la connectivité et de la proportion d’habitat disponible sur les communautés prairiales, des régressions multiples ont été réalisées entre chaque indice de diversité (S, H’,J’ et Ss H’s, J’s) et les indices paysagers PLAND et ENN_MN (Tab.1)
A ces régressions, a été ajoutée la covariable « taille de la parcelle » pour intégrer au modèle son effet potentiel sur la diversité spécifique. Les mêmes analyses ont ensuite été réalisées sur deux sous-groupes de prairies : les prairies pâturées (28 parcelles) et les prairies de fauche (9 parcelles), afin de prendre en compte un effet potentiel du mode de gestion.
Enfin, une analyse canonique des correspondances a été réalisée afin de tester l’influence des variables environnementales (ENN_MN, PLAND, taille de la parcelle et mode de gestion) sur la composition floristique des prairies.
10
Résultats
- Caractéristiques des communautés végétales étudiées
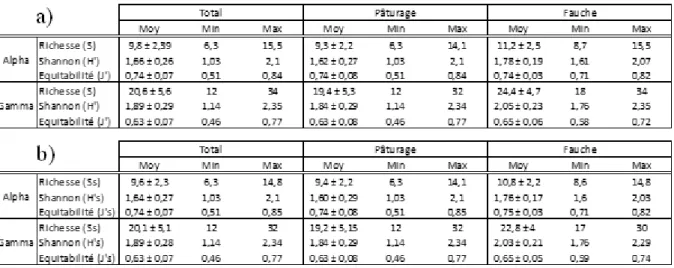
En comparant les caractéristiques des communautés de la matrice totale et de la matrice spécialiste (Tab.2), nous remarquons qu’il existe très peu de différence. Cela indique que les prairies échantillonnées étaient représentées majoritairement par des espèces spécialistes.
Nous observons une richesse spécifique et une diversité plus importante en prairie de fauche plutôt qu’en prairie pâturée.
Tableau 2 : Distribution des indices de diversité (S, H’, J’) à l’échelle alpha et gamma, sur la totalité des données, sur le sous-ensemble « Pâturage » pour les prairies pâturées et le sous-ensemble « Fauche » pour les prairies fauchées de la matrice totale d’espèces (a) et de la matrice d’espèces « spécialistes » (b).
11 - Résultats des régressions multiples
Tableau 3 : Résultats du modèle de régression linéaire pour la richesse, la diversité de Shannon et l’équitabilité, alpha et gamma, en fonction de la proportion d’habitats prairiaux (PLAND) et la connectivité entre les prairies (ENN_MN) dans un buffer de 200 mètres autour des prairies échantillonnées sur la matrice totale. P= Prairies pâturées ; F=Prairies de Fauche ; ns : non significatif ; • : < 0.1 ; * : < 0.05 ; ** : < 0.01
p-value R² estimate p-value estimate p-value estimate p-value
Total • 0,18 ns ns -1,37 • P • 0,28 -0,059 • -0,056 * ns F ns Total ns P ns F ns Total ns P ns F • 0,69 ns ns ns Total ns P ns F ns Total • 0,2 ns ns -0,19 * P ns F ns Total ns P ns F * 0,81 ns -0,003 ** -0,067 • Alpha Gamma Shannon (H') Equitabilité (J') Equitabilité (J') Richesse (S) ENN_MN PLAND Taille Richesse (S) Shannon (H') Modèle global
12 Tableau 4 : Résultats du modèle de régression linéaire pour la richesse, la diversité de Shannon et l’ équitabilité, alpha et gamma, en fonction de la connectivité entre les prairies (ENN_MN) et la proportion d’habitats prairiaux (PLAND), dans un buffer de 200 mètres autour des prairies échantillonnées, sur la matrice « spécialistes ». P= Prairies pâturées ; F=Prairies de Fauche ; ns : non significatif ; • : < 0.1 ; * : < 0.05 ; ** : < 0.01
La proportion d’habitat n’a pas d’effet significatif sur la diversité prairiale au seuil α = 5%. En prenant un seuil α = 10% pour le modèle global et l’effet des variables testées, on peut tout de même observer une tendance. La proportion d’habitat tend à influencer négativement la richesse alpha S et Ss (Tab.3 et 4 ; p-value < 0,1) des prairies pâturées mais avec un pourcentage faible de variance expliquée par le modèle (r²=0,28).
La connectivité présente un effet significatif sur l’équitabilité (r²=80%, p-value < 0,01 ) J’ et J’s (Tab.3 et 4), à l’échelle gamma des prairies de fauche. Quand la distante entre prairie augmente (augmentation de la valeur de l’indice ENN_MN), l’équitabilité diminue. L’augmentation de la connectivité favorise ainsi une forte équitablité des communautés végétales des prairies de fauche à l’échelle de la parcelle.
Si nous tenons compte des résultats des modèles globaux et des effets des variables testées obtenus au seuil α=10%, une augmentation de l’indice ENN_MN tend à diminuer la
p-value R² estimate p-value estimate p-value estimate p-value
Total • 0,19 ns -0,032 • -1,28 • P * 0,29 -0,061 • -0,056 * ns F ns Total ns P ns F ns Total ns P ns F ns Total ns P ns F ns Total • 0,21 ns ns -0,19 • P ns F ns Total ns P ns F * 0,8 ns -0,003 ** -0,067 •
Modèle global PLAND ENN_MN
Gamma Richesse (Ss) Shannon (H's) Equitabilité (J's) Taille Alpha Richesse (Ss) Shannon (H's) Equitabilité (J's)
13 richesse alpha (r² = 29% , p-value < 0,05) S et Ss (Tab.3 et 4) des prairies pâturées, de même que la richesse alpha (r²=19% , p-value <0,1) Ss (Tab.4) de l’ensemble des prairies. Ainsi, la richesse des communautés végétales augmente quand la connectivité entre prairies augmente.
La taille de la parcelle n’a pas d’effet significatif sur la diversité des commuanutés prairiales au seuil α= 5%. En tenant compte des résultats au seuil α = 10%, la taille de la parcelle tend à influencer négativement : (i) la richesse alpha ( r²=18%, p-value <0,1) S et Ss (Tab.3 et 4) de l’ensemble des prairies (ii) l’indice de Shannon à l’échelle gamma H’ (Tab.3 ; p-value < 0,1 ) et H’s (Tab.4 ; p-value <0,05) avec un modèle expliquant peu la variance observée (r²=20%) (iii) l’équitabilité J’ et J’s (Tab.3 et 4 ) des prairies de fauche (p-value < 0,1) avec un modèle global significatif (p-value < 0.05) et un bon pourcentage de variance expliquée (r²=80%).
Les principaux constats sont les suivants :
(1) La richesse à l’échelle du quadrat (alpha) et l’équitabilité à l’échelle de la parcelle (gamma) semblent être les meilleurs indicateurs pour montrer l’effet du paysage sur les communautés de plantes prairiales ;
(2) La communauté d’espèces spécialistes de l’habitat prairial sont plus sensibles à la connectivité sur l’ensemble des parcelles que la communauté totale ;
(3) Le mode de gestion (pâturage vs fauche) influence la structuration des communautés de plantes prairiales ;
14 - Résultats de l’analyse canonique des correspondances
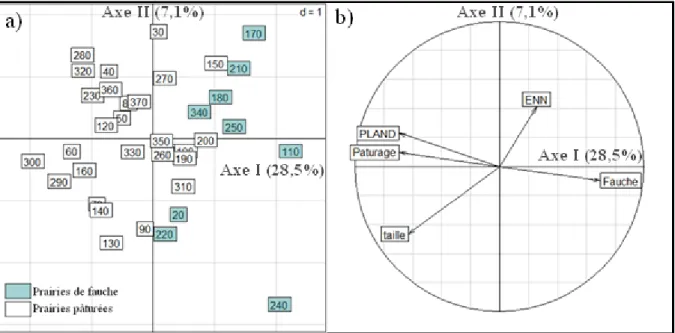
Figure 5 : Représentation graphique des quadrats (a) et des variables environnementales (b) sur l’axe I et sur l’axe II, de l’analyse canonique des correspondances. Les quadrats sont numérotés de 0 à 370, avec 10 quadrats par prairie.
L’analyse canonique des correspondances (ACC) explique significativement 45,8% de la variance observée (Test de Monte-Carlo, p-value = 0,01) dont 28,5% est expliqué par l’axe I. L’axe I est caractérisé par le mode de gestion et oppose les prairies de fauche (à droite) et aux prairies pâturées (à gauche ; Fig.5).
Le mode de gestion montre un effet important sur la répartition des espèces car les variables « Pâturage » et « Fauche » sont proches du cercle de corrélation (Fig.5b).
La variable PLAND se trouve également proche du cercle de corrélation (Fig.5b) et semble influencer plus spécifiquement la répartition des espèces de prairies pâturées.
Enfin, l’ACC semble confirmer l’effet de la superficie des prairies observé dans les régressions multiples (Tab.3 et 4).
15
Discussion
- Effet de la proportion d’habitat sur la diversité et la composition
spécifique des prairies
Contrairement à notre première hypothèse, une plus grande proportion de prairies dans le paysage n’augmente pas la richesse, la diversité et l’équitabilité des communautés végétales au sein des prairies.
Nous avons démontré qu’une forte proportion de prairies autour de la prairie étudiée tend à diminuer la richesse alpha et à influencer la composition spécifique des prairies pâturées. Elle ne présente pas d’effet sur leur diversité et leur équitabilité. Tandis que les prairies de fauche ne présentent aucune tendance.
Ce résultat contredit les résultats d’études précédentes qui ont montré que la richesse spécifique des tâches d’habitat d’une matrice environnementale augmentait avec la proportion d’habitat à l’échelle du paysage local, pour des sites de même taille.
Notre résultat peut s’expliquer par une forte pression de la composante "gestion". Une forte intensité de pâturage a perturbé les communautés végétales à travers un effet plus fort des facteurs locaux qui ont éclipsé ceux du paysage, et en influençant l’établissement des plantes (Matthews et al., 2009).
On peut supposer que le nombre de prairies de fauche était insuffisant et que la variabilité propre aux facteurs locaux de chaque prairie était trop importante pour montrer un effet de la proportion d’habitat.
- Effet de connectivité sur la diversité spécifique
Notre deuxième hypothèse est partiellement validée. A l’échelle gamma, c’est-à-dire à l’échelle de la parcelle, la connectivité paysagère augmente l’équitabilité des prairies de fauche, mais n’influence pas l’équitabilité des prairies pâturées. A l’échelle locale, la connectivité augmente la richesse des prairies pâturées mais n’influence pas la richesse des prairies de fauche. Elle tend à augmenter la richesse de la communauté de spécialistes sur l’ensemble des
16 prairies. La composition floristique n’est pas dépendante de la connectivité, de même que la diversité.
Ainsi, les prairies répondent différemment à la connectivité en fonction de leur mode de gestion (Brudvig, 2016 ; Matthews et al., 2009).
Les prairies pâturées sont des milieux hétérogènes. En effet, le piétinement et l’herbivorie retrouvé dans ce genre de prairies va créer des micro-niches qui vont favoriser la pousse de propagules arrivées par dispersion, grâce à la connectivité paysagère entre les prairies alentours. Nous retrouverons alors des zones riches en espèces et d’autres zones plus pauvres. C’est pourquoi nous observons l’effet de la connectivité à l’échelle locale et non pas à l’échelle gamma.
Les prairies de fauche sont plus homogènes que les prairies pâturées. Ainsi la connectivité influence ce milieu à une échelle supérieure, celui de la parcelle. La diminution de la distance entre les tâches d’habitat favorable, en augmentant leur connectivité, favorisera la dispersion des propagules.
Enfin, la différence d’effet de la connectivité sur la richesse alpha de l’ensemble des prairies entre la communauté totale et la communauté de spécialistes est peut être due à une sensibilité plus importante des espèces spécialistes à la connectivité. L’ensemble des parcelles échantillonnées comportant une majorité de prairies pâturées (28 contre 9 de fauches), leur réponse à la connectivité, comme expliqué ci-dessus, doit dominer.
- Effet de la taille de la parcelle sur la diversité spécifique
L’effet de la superficie a été agrémenté au modèle afin d’y ajouter son effet éventuel sur la diversité et la richesse de plantes prairiales.
Nous avons démontré que la taille de la parcelle tend à diminuer la richesse alpha et la diversité gamma de l’ensemble des prairies. De même, elle diminue l’équitabilité des prairies de fauches.
Ce résultat contredit l’attendu classique, qui annonce qu’une grande taille de l’habitat local maintient une plus grande richesse spécifique
17
Conclusion et perspectives
Dans cette étude, nous avons cherché à montrer l’influence des continuités écologiques sur la structuration des communautés de plantes prairiales en analysant l’effet de la connectivité et de la proportion d’habitat sur la richesse, la diversité, l’équitabilité et la composition spécifique de 37 prairies de longue durée. Nos résultats montrent ainsi l’effet positif de la connectivité. Cependant, nous avons pu observer l’importance des facteurs locaux dans l’expression des facteurs paysagers et notamment celui de la gestion. Il serait donc intéressant de déterminer la part de chaque facteur pour répondre au mieux à la problématique.
Les continuités écologiques apparaissant aujourd’hui comme des solutions aux pertes massives de biodiversité dues à la fragmentation, cette étude peut s’intégrer dans le projet de mise en place de la Trame Verte et Bleue, lancé par les lois Grenelle. Il a pour pour but de favoriser la protection des réservoirs de Biodiversité et les échanges entre ces réservoirs en identifiant, protégeant, et restaurant les continuités écologiques.
Bibliographie
Bakker, J. P., & Berendse, F. (1999). Constraints in the restoration of ecological diversity in grassland and heathland communities. Trends in ecology & evolution, 14(2), 63-68.
Beier, Paul, et Reed F. Noss. « Do Habitat Corridors Provide Connectivity? » Conservation
Biology 12, no 6 (s. d.): 1241-52.
Bowne, David R., et Michael A. Bowers. « Interpatch Movements in Spatially Structured Populations: A Literature Review ». Landscape Ecology 19, no 1 (1 janvier 2004): 1-20.
Bravard, Jean-Paul. « Burel F., Baudry j., Écologie du paysage. Concepts, méthodes et applications ». Annales de géographie 110, no 618 (2001): 201-201.
Chetkiewicz, Cheryl-Lesley, Colleen St. Clair, et Mark Boyce. « Corridors for Conservation: Integrating Pattern and Process ». Annual Review of Ecology Evolution and Systematics 37 (1 décembre 2006): 317-42.
18 Chisci, G., and Zanchi, C. (1981). The influence of different tillage systems and different crops on
soil losses on hilly silty-clayey soil. In 'Soil Conservation'. (Ed. R. P. C. Morgan.) (Wiley: New York.)
Fahrig, Lenore. « Effects of Habitat Fragmentation on Biodiversity ». Annual Review of Ecology,
Evolution, and Systematics 34, no 1 (1 novembre 2003): 487-515.
Fahrig, Lenore, et al. « Functional landscape heterogeneity and animal biodiversity in agricultural landscapes ». Ecology letters 14.2 (2011): 101-112.
Fahrig, Lenore, et Gray Merriam. « Habitat Patch Connectivity and Population Survival ». Ecology 66, no 6 (s. d.): 1762-68.
Fischer, Joern, et David B. Lindenmayer. « Fischer J, Lindenmayer DB Landscape modification and habitat fragmentation: a synthesis. Glob Ecol Biogeogr ». Global Ecology and
Biogeography 16 (1 mai 2007): 265-80.
Frankham, Richard, Jonathan D Ballou, et David A Briscoe. « Introduction to Conservation Genetics ». Cambridge University Press, s. d., 19.
Hanski, Ilkka. « A Practical Model of Metapopulation Dynamics ». Journal of Animal Ecology 63, no 1 (1994): 151-62.
Huyghe, Christian. « La multifonctionnalité des prairies en France II. Conciliation des fonctions de production et de préservation de l’environnement ». Cahiers Agricultures 18, no 1 (1 janvier
2009): 7-16 (1).
Kadoya, Taku. « Assessing Functional Connectivity Using Empirical Data ». Population Ecology 51, no 1 (1 janvier 2009): 5-15.
Matthews, Jeffrey W., Ariane L. Peralta, Diana N. Flanagan, Patrick M. Baldwin, Arun Soni, Angela D. Kent, et Anton G. Endress. « Relative Influence of Landscape vs. Local Factors on Plant Community Assembly in Restored Wetlands ». Ecological Applications 19, no 8 (s. d.): 2108-23.
19 Öckinger, E., & Smith, H. G. (2007). Semi‐natural grasslands as population sources for pollinating
insects in agricultural landscapes. Journal of applied ecology, 44(1), 50-59.
Olff, H., & Ritchie, M. E. (1998). Effects of herbivores on grassland plant diversity. Trends in
ecology & evolution, 13(7), 261-265.
Peyraud, Jean-Louis, Alain Peeters, et Alex De Vliegher. « Place et atouts des prairies permanentes en France et en Europe ». Fourrages 211 , 195-204. (2012), 2012.
Taylor, Philip D., Lenore Fahrig, Kringen Henein, et Gray Merriam. « Connectivity Is a Vital Element of Landscape Structure ». Oikos 68, no 3 (1993): 571-73.
Sites internet :
20
Annexe
Annexe 1 : Photographie d’un quadrat de 4 m² dans une des prairies échantillonnées
Annexe 2 : Tableau de la distribution des variables PLAND, ENN_MN et taille de la parcelle des prairies de la matrice totale.
Annexe 3 : Tableau de l’inertie obtenue par chacun des axes de l’analyse canonique des correspondances.
Glossaire
Zone Atelier : dispositif interdisciplinaire de recherche à long terme sur l’environnement et
les interactions hommes-milieux
Moy Min Max
PLAND (%) 40,2 ± 16 9,1 73,4
ENN_MN (m) 33,4± 23,5 0 95
Taille (ha) 0,94 ± 0,53 0,19 2,33
Inertie Inertie cumulée Inertie cumulée relative (%)
Ax1 0,285 0,285 62,2
Ax2 0,071 0,356 77,8
Ax3 0,061 0,417 91,1
21
Résumé
Cette étude est basée sur l’analyse du rôle de continuités écologiques sur la structuration des communautés de plantes prairiales, en réponse à la fragmentation toujours grandissante des paysages et notamment des paysages agricoles. Elle cherche ainsi à montrer l’influence de la proportion d’habitat favorable et de la connectivité entre ces tâches d’habitat sur la structuration des communautés de plantes prairiales et ceux à plusieurs échelles. Elle montre ainsi le rôle de la continuité sur la dispersion et l’établissement des plantes de prairies, mais également l’importance de facteurs locaux dans ce genre d’étude.
Abstract
This study is based on the analysis of the role of ecological continuities on the structuring of grassland plant communities, in response to the ever-increasing fragmentation of landscapes and particularly agricultural landscapes. It seeks to show the influence of the proportion of favorable habitat and connectivity between these habitat tasks on the structuring of grassland and multi-scale plant communities. It shows the role of continuity in the dispersal and establishment of grassland plants, but also the importance of local factors in this type of study.