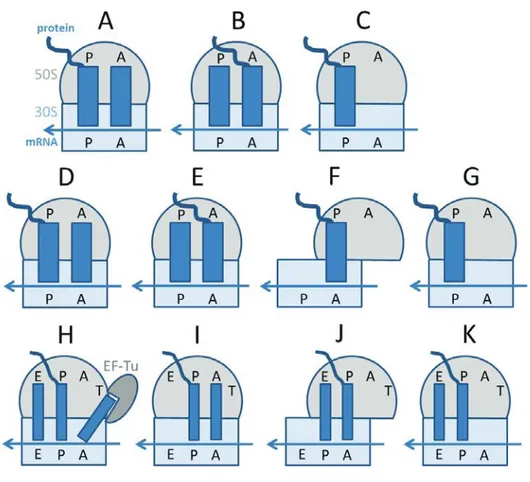
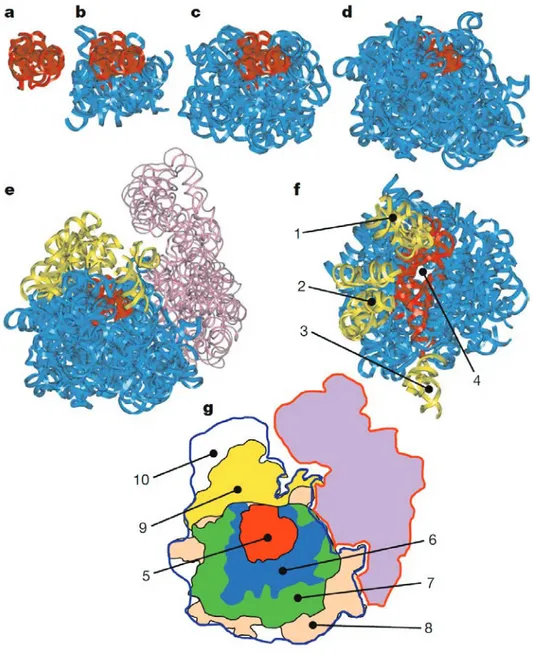
Structural aspects of the ribosome evolution and function
163
0
0
Texte intégral
Figure
Documents relatifs